Стабильность и устойчивость экосистем — Промышленная экология
Важнейшими показателями динамики экосистем являются устойчивость и стабильность. Определение устойчивости экосистем уже давалось — это способность экосистемы возвращаться в исходное состояние после снятия внешнего воздействия, выведшего ее из равновесия. Под стабильностью экосистемы понимают ее способность сохранять свою структуру и функциональные свойства при воздействии на нее внешних факторов. Иногда понятия устойчивость и стабильность рассматриваются как синонимы, но тогда следует различать два вида устойчивости: резидентная устойчивость (стабильность) — способность оставаться в устойчивом (равновесном) состоянии под нагрузкой, и упругая устойчивость (собственно устойчивость) — способность быстро восстанавливаться при снятии нагрузки. В разных терминах имеются различные смысловые оттенки, которые нужно учитывать.
Системы с высокой резидентной устойчивостью способны воспринимать значительные воздействия, не изменяя существенно своей структуры, то есть практически не выходя за пределы равновесного состояния. Поэтому понятие упругой устойчивости для них не определено (если система не выходила за пределы равновесия, то как можно говорить о возвращении в равновесное состояние после снятия возмущения). Если внешнее воздействие превышает определенные критические значения, то такая система обычно разрушается. В технике подобное качество называется жесткостью. Предельные значения внешних воздействий, которые система способна выдержать без разрушения соответствуют запасу жесткости. Когда говорят о высокой резидентной устойчивости, то имеется в виду именно высокий запас жесткости данной системы. Это несколько отличается от понятия высокой стабильности, так как здесь в первую очередь внимание обращается на неизменность структуры. Тундра, например, обладает высокой стабильностью, но она очень ранима, у нее малый запас жесткости, то есть малая резидентная устойчивость. Экосистему тундры очень легко разрушить. Достаточно проехать вездеходу. Колеи, которые он оставляют за собой, сохраняются десятилетиями. Такие экосистемы по аналогии с техникой можно назвать хрупкими.
Системы с малой резидентной устойчивостью для нормального существования должны обладать высокой упругой устойчивостью. Они более чувствительны к внешним возмущениям, под действием которых они как бы «прогибаются», частично деформируя свою структуру, но после снятия или ослабления внешних воздействий быстро возвращаются в исходное равновесное состояние. При превышении пороговых воздействий такая система теряет устойчивость, то есть все дальше удаляется от состояния равновесия. Диапазон воздействий, которые может выдержать система без разрушения, в технике соответствует запасу упругости. Таким образом, степень упругой устойчивости можно оценить как упругостью, определяющей степень сопротивления внешнему воздействию и скорость возврата в исходное состояние после снятия воздействия, так и запасом упругости. В отличие от упругих систем, пластичные системы после снятия внешнего воздействия не возвращаются в исходное состояние, а приходят к какому-то другому равновесному состоянию. Так согласно точке зрения оппонентов теории моноклимакса, для экосистем характерно не одно, а несколько состояний равновесия (климакса). Таким образом, для пластичных экосистем характерна малая упругая и малая резидентная устойчивость.
Похоже на то, что резидентная и упругая устойчивости взаимоисключают друг друга, точнее, экосистеме трудно развивать оба вида устойчивости. Например, одни леса состоят из деревьев с толстой корой, обладающих повышенной резидентной устойчивостью к пожарам. Но если такой лес все-таки сгорит, то его восстановление, как правило, крайне проблематично. Напротив, многие леса очень часто горят (низкая резидентная устойчивость), но быстро восстанавливаются (высокая упругая устойчивость). Ориентация экосистем на один из видов устойчивости определяется, как правило, изменчивостью среды: при стабильных условиях экосистемы склонны к более высокой резидентной устойчивости, при изменчивых условиях предпочтение отдается упругой устойчивости.
Рис.12.3
Природа обоих видов устойчивости состоит в наличии в экосистеме разного рода обратных связей. В основном это отрицательные обратные связи, которые направлены на стабилизацию параметров экосистемы, возвращая их значения к какой-то изначально заданной величине. Однако немаловажную роль играют и положительные обратные связи, усиливающие благоприятные для системы изменения, например, в плане роста и выживаемости организмов. Однако деятельность положительных обратных связей обязательно должна быть ограничена соответствующими отрицательными обратными связями, иначе ничем не контролируемая экспансия жизни может привести экосистему к гибели. В обоих случаях схема управления выглядит одинаково (рис.25).
Механизмы реализации обратных связей называются механизмами гомеостаза (от греческих слов homoios — подобный, одинаковый и ststis — состояние). Обычно они достаточно сложны, так как их компоненты связаны в единые информационные сети факторами среды, а также различными физическими и химическими агентами – «посредниками», подобно тому, как нервная или гормональная системы связывают в одно целое части организма. Примером может служить сильная реакция организмов в экосистеме на слабые концентрации некоторых веществ. С энергетической точки зрения можно выделить одну закономерность, о которой уже говорилось ранее: обладатели высококачественной энергии, оставаясь в меньшинстве, управляют большими потоками энергии более низкого качества. Например, хищники управляют численностью травоядных животных. В то же время активность хищников и их численность управляется численностью их жертв по цепи обратной связи. По этой цепи на более высококачественный энергетический уровень (к хищникам) подается небольшая часть низкокачественной энергии — хищники уничтожают не всех жертв, а лишь небольшой их процент. Причем влияние этой части энергии на управление всей системой существенно усиливается деятельностью хищников.
Помимо отрицательной обратной связи устойчивость экосистемы может быть обеспечена избыточностью компонентов. Например, если в системе имеется несколько автотрофов, каждый из которых характеризуется своим температурным диапазоном функционирования, то скорость фотосинтеза сообщества в целом может оставаться неизменной, несмотря на колебания температуры. То же самое можно сказать и про гетеротрофов. Поэтому видовое разнообразие способствует повышению устойчивости экосистем (закон Эшби). При этом как в случае резидентной, так и в случае упругой устойчивости расширяется диапазон пороговых воздействий, которые способны выдержать экосистема без разрушения (запас жесткости и запас упругости).
Так, например, для тундры с ее бедным разнообразием характерны такие проявления низкой упругой устойчивости, как периодические резкие увеличения численности мелких грызунов — леммингов. О малой резидентной устойчивости тундры мы уже говорили. Экосистема тундры очень ранима и очень долго восстанавливается. В то же время тропические леса с их богатым разнообразием способны достаточно быстро залечивать свои раны. Легенды повествуют о целых городах, поглощенных джунглями, население которых не могло справиться с их натиском и вынуждено было уходить в другие районы.
Если разные стадии сукцессии экосистемы считать квазистатическими состояниями, то здесь можно отметить ряд закономерностей. На первых стадиях сукцессии экосистемы способны очень быстро возобновляться. Например, мы можем распахать по весне все поле, но уже к осени оно будет укрыто травами. Это говорит о достаточно высокой упругой устойчивости на начальных стадиях сукцессии, которая по мере движения к климаксу постоянно снижается. То есть на каждой следующей стадии для восстановления исходного состояния требуется гораздо больше времени. Климаксный лес характеризуется очень малой упругой устойчивостью.
Для резидентной устойчивости тенденция обратная. На первых стадиях сукцессии резидентная устойчивость остается достаточно невысокой — системы очень чувствительны к внешним воздействиям, откликаясь на них повышенным динамизмом. Например, заросли кустарника или молодой березняк может быть легко поврежден вездеходом (малая резидентная устойчивость), однако вторичная сукцессия достаточно быстро «затянет» повреждение (высокая упругая устойчивость). По мере роста видового разнообразия резидентная устойчивость непрерывно растет. При этом растет одновременно и жесткость системы (стабильность) и запас жесткости (взрослый лес вездеход так просто не одолеет). Однако по мере приближения к климаксу, как уже было показано, экосистема становится более чувствительна к действию некоторых катастрофических факторов среды (бури, пожары и т.п.). То есть на фоне продолжающегося роста стабильности (жесткости), запас жесткости экосистемы при движении ее к климаксу, по-видимому, начинает уменьшаться, что можно объяснить преобладанием в ней старых деревьев. Это говорит о том, что она становится все более хрупкой.
Как уже было сказано, для реальных лесов характерен, как правило, мозаичный климакс. По-видимому, именно такой лес имеет наилучший комплекс показателей устойчивости и стабильности. Периодические сильные воздействия на него в целом лес выдерживает достаточно жестко, жертвуя лишь наиболее старыми и хрупкими участками, которые, даже если они превращаются после пожара в абсолютно пустые пространства, в условиях непосредственной близости неповрежденного леса восстанавливаются сравнительно быстро.
Примером абсолютно нежизнеспособных экосистем являются агроценозы, создаваемы человеком из однолетних и двулетних культурных растений, то есть продукты сельскохозяйственной деятельности человека. Многие экологи даже не считают их экосистемами, хотя есть все основания считать их экосистемами с искусственно поддерживаемыми начальными стадиями сукцессии. Заброшенные поля сразу же втягиваются в естественный ход сукцессии. Что касается возделываемых полей, то они существуют только за счет человека. Свидетельством их нежизнеспособности является крайне малая устойчивость как упругая, так и резидентная, что является следствием очень скудного видового разнообразия как флоры (все сорняки подавляются гербицидами), так и фауны («вредных» насекомых мы травим пестицидами). Малая упругая устойчивость проявляется, например, в резких вспышках численности конкретных видов вредителей. Подобные вспышки в естественной природе наблюдаются очень редко. Малая резидентная устойчивость проявляется, например, в повышенной чувствительности посевов к природным условиям, таким как град, засуха, повышенное количество осадков и т.д.
Осваивая все большие территории планеты, человек продолжает стратегию обеднения видового разнообразия. Этим он все более подрывает способность природы сопротивляться внешним воздействиям и возвращаться в исходное состояние. Как знать, может быть, роковая черта, за которой начинаются необратимые изменения, уже пройдена. Это значит, что биосфера никогда больше не придет к исходному состоянию. Это значит, что мы вступили на путь глобальных перемен, которые постепенно изменят жизнь на Земле самым коренным образом. Будет ли в этом мире место человеку?
ekologyprom.ru
Высокая резидентная устойчивость — МегаЛекции
Содержание
Введение……………………………………………….………….………….3
1. Высокая резидентная устойчивость …………………….………………4
2. Высокая упругая устойчивость …… ……………….……………………4
3. Обратные связи ………………………………………………….……….5
4. Нежизнеспособные экосистемы ……………………………….…………7
5. Биоразнообразие и устойчивость в экосистемах …………….………….7
6. Сложность и устойчивость экосистем………………………….………..8
6.1. Параметры сложности и их влияние на устойчивость.……………….8
Заключение …………………………….……………..…………………….10
Список литературы…….……………………………………..…………….11
Введение
Важнейшими показателями динамики экосистем являются устойчивость и стабильность. Устойчивость экосистем — это способность экосистемы возвращаться в исходное состояние после снятия внешнего воздействия, выведшего ее из равновесия.
Под стабильностью экосистемы понимают ее способность сохранять свою структуру и функциональные свойства при воздействии на нее внешних факторов.
Устойчивость и сбалансированность процессов, протекающих в экосистемах, позволяет констатировать, что им в целом свойственно состояние гомеостаза, подобно входящим в их состав популяциям и каждому живому организму. Нестабильность среды обитания в экосистемах компенсируется биоценотическими адаптивными механизмами.
В естественной экосистеме постоянно поддерживается равновесие, исключающее необратимое уничтожение тех или иных звеньев трофической сети. Это является следствием длительного эволюционного процесса, названного Ч. Дарвиным естественным отбором. Любая экосистема всегда сбалансирована и устойчива (гомеостатична), причем системы тем стабильнее во времени и пространстве, чем они сложнее.
Гомеостаз — саморегуляция, способность открытой системы сохранять постоянство своего внутреннего состояния посредством скоординированных реакций, направленных на поддержание динамического равновесия. Стремление системы воспроизводить себя, восстанавливать утраченное равновесие, преодолевать сопротивление внешней среды.
Обычно выделяют два типа гомеостаза: резистентный — способность экосистем сохранять структуру и функции при негативном внешнем воздействии и упругий — способность экосистемы восстанавливать структуру и функции при утрате части компонентов экосистемы.
Иногда выделяют третий аспект устойчивости — устойчивость экосистемы по отношению к изменениям характеристик среды и изменению своих внутренних характеристик. В случае, если экосистема устойчиво функционирует в широком диапазоне параметров окружающей среды и/или в экосистеме присутствует большое число взаимозаменяемых видов (то есть, когда различные виды, сходные по экологическим функциям в экосистеме, могут замещать друг друга), такое сообщество называют динамически прочным (устойчивым). В обратном случае, когда экосистема может существовать в весьма ограниченном наборе параметров окружающей среды, и/или большинство видов незаменимы в своих функциях, такое сообщество называется динамически хрупким (неустойчивым). Необходимо отметить, что данная характеристика в общем случае не зависит от числа видов и сложности сообществ. Классическим примером может служить Большой Барьерный риф у берегов Австралии (северо-восточное побережье), являющийся одной из «горячих точек» биоразнообразия в мире — симбиотические водоросли кораллов, динофлагелляты, весьма чувствительны к температуре. Отклонение от оптимума буквально на пару градусов ведёт к гибели водорослей, а до 50-60 % (по некоторым источникам до 90 %) питательных веществ полипы получают от фотосинтеза своих мутуалистов.
Высокая резидентная устойчивость
Системы с высокой резидентной устойчивостью способны воспринимать значительные воздействия, не изменяя существенно своей структуры, то есть практически не выходя за пределы равновесного состояния. Поэтому понятие упругой устойчивости для них не определено (если система не выходила за пределы равновесия, то как можно говорить о возвращении в равновесное состояние после снятия возмущения). Если внешнее воздействие превышает определенные критические значения, то такая система обычно разрушается. В технике подобное качество называется жесткостью. Предельные значения внешних воздействий, которые система способна выдержать без разрушения соответствуют запасу жесткости. Когда говорят о высокой резидентной устойчивости, то имеется в виду именно высокий запас жесткости данной системы. Это несколько отличается от понятия высокой стабильности, так как здесь в первую очередь внимание обращается на неизменность структуры. Тундра, например, обладает высокой стабильностью, но она очень ранима, у нее малый запас жесткости, то есть малая резидентная устойчивость. Экосистему тундры очень легко разрушить. Достаточно проехать вездеходу. Колеи, которые он оставляют за собой, сохраняются десятилетиями. Такие экосистемы по аналогии с техникой можно назвать хрупкими.
2. Высокая упругая устойчивость.
Системы с малой резидентной устойчивостью для нормального существования должны обладать высокой упругой устойчивостью. Они более чувствительны к внешним возмущениям, под действием которых они как бы “прогибаются”, частично деформируя свою структуру, но после снятия или ослабления внешних воздействий быстро возвращаются в исходное равновесное состояние. При превышении пороговых воздействий такая система теряет устойчивость, то есть все дальше удаляется от состояния равновесия. Диапазон воздействий, которые может выдержать система без разрушения, в технике соответствует запасу упругости. Таким образом, степень упругой устойчивости можно оценить как упругостью, определяющей степень сопротивления внешнему воздействию и скорость возврата в исходное состояние после снятия воздействия, так и запасом упругости. В отличие от упругих систем, пластичные системы после снятия внешнего воздействия не возвращаются в исходное состояние, а приходят к какому-то другому равновесному состоянию. Так согласно точке зрения оппонентов теории моноклимакса, для экосистем характерно не одно, а несколько состояний равновесия (климакса). Таким образом, для пластичных экосистем характерна малая упругая и малая резидентная устойчивость.
Оба типа стабильности исключают друг друга, или, другими словами, системе трудно одновременно развить оба типа устойчивости. Так, калифорнийский лес из секвойи довольно устойчив к пожарам (для этих деревьев характерна толстая кора и другие адаптации), но если он все же сгорит, то восстанавливается очень медленно или вовсе не восстанавливается. Напротив, калифорнийские заросли чапараля очень легко выгорают (низкая резистентная устойчивость), но быстро восстанавливаются, буквально за несколько лет (отличная упругая устойчивость). Как правило, при благоприятных физических условиях среды экосистемы в большей степени проявляют резистентную устойчивость, а не упругую, но в изменчивых физических условиях наблюдается прямо противоположное.
Обратные связи.
Природа обоих видов устойчивости состоит в наличии в экосистеме разного рода обратных связей. В основном это отрицательные обратные связи, которые направлены на стабилизацию параметров экосистемы, возвращая их значения к какой-то изначально заданной величине. Однако немаловажную роль играют и положительные обратные связи, усиливающие благоприятные для системы изменения, например, в плане роста и выживаемости организмов. Однако деятельность положительных обратных связей обязательно должна быть ограничена соответствующими отрицательными обратными связями, иначе ничем не контролируемая экспансия жизни может привести экосистему к гибели.
Механизмы реализации обратных связей называются механизмами гомеостаза (от греческих слов homoios — подобный, одинаковый и ststis — состояние). Обычно они достаточно сложны, так как их компоненты связаны в единые информационные сети факторами среды, а также различными физическими и химическими агентами — “посредниками”, подобно тому, как нервная или гормональная системы связывают в одно целое части организма. Примером может служить сильная реакция организмов в экосистеме на слабые концентрации некоторых веществ. С энергетической точки зрения можно выделить одну закономерность: обладатели высококачественной энергии, оставаясь в меньшинстве, управляют большими потоками энергии более низкого качества. Например, хищники управляют численностью травоядных животных. В то же время активность хищников и их численность управляется численностью их жертв по цепи обратной связи. По этой цепи на более высококачественный энергетический уровень (к хищникам) подается небольшая часть низкокачественной энергии — хищники уничтожают не всех жертв, а лишь небольшой их процент. Причем влияние этой части энергии на управление всей системой существенно усиливается деятельностью хищников.
Помимо отрицательной обратной связи устойчивость экосистемы может быть обеспечена избыточностью компонентов. Например, если в системе имеется несколько автотрофов, каждый из которых характеризуется своим температурным диапазоном функционирования, то скорость фотосинтеза сообщества в целом может оставаться неизменной, несмотря на колебания температуры. То же самое можно сказать и про гетеротрофов. Поэтому видовое разнообразие способствует повышению устойчивости экосистем (закон Эшби). При этом как в случае резидентной, так и в случае упругой устойчивости расширяется диапазон пороговых воздействий, которые способны выдержать экосистема без разрушения (запас жесткости и запас упругости).
Так, например, для тундры с ее бедным разнообразием характерны такие проявления низкой упругой устойчивости, как периодические резкие увеличения численности мелких грызунов — леммингов. О малой резидентной устойчивости тундры мы уже говорили. Экосистема тундры очень ранима и очень долго восстанавливается. В то же время тропические леса с их богатым разнообразием способны достаточно быстро залечивать свои раны. Легенды повествуют о целых городах, поглощенных джунглями, население которых не могло справиться с их натиском и вынуждено было уходить в другие районы.
Если разные стадии сукцессии экосистемы считать квазистатическими состояниями, то здесь можно отметить ряд закономерностей. На первых стадиях сукцессии экосистемы способны очень быстро возобновляться. Например, мы можем распахать по весне все поле , но уже к осени оно будет укрыто травами. Это говорит о достаточно высокой упругой устойчивости на начальных стадиях сукцессии, которая по мере движения к климаксу постоянно снижается. То есть на каждой следующей стадии для восстановления исходного состояния требуется гораздо больше времени. Климаксный лес характеризуется очень малой упругой устойчивостью.
Для резидентной устойчивости тенденция обратная. На первых стадиях сукцессии резидентная устойчивость остается достаточно невысокой — системы очень чувствительны к внешним воздействиям, откликаясь на них повышенным динамизмом. Например, заросли кустарника или молодой березняк может быть легко поврежден вездеходом (малая резидентная устойчивость), однако вторичная сукцессия достаточно быстро “затянет” повреждение (высокая упругая устойчивость). По мере роста видового разнообразия резидентная устойчивость непрерывно растет. При этом растет одновременно и жесткость системы (стабильность) и запас жесткости (взрослый лес вездеход так просто не одолеет). Однако по мере приближения к климаксу, как уже было показано, экосистема становится более чувствительна к действию некоторых катастрофических факторов среды (бури, пожары и т.п.). То есть на фоне продолжающегося роста стабильности (жесткости), запас жесткости экосистемы при движении ее к климаксу, по-видимому, начинает уменьшаться, что можно объяснить преобладанием в ней старых деревьев. Это говорит о том, что она становится все более хрупкой.
Как уже было сказано, для реальных лесов характерен, как правило, мозаичный климакс. По-видимому, именно такой лес имеет наилучший комплекс показателей устойчивости и стабильности. Периодические сильные воздействия на него в целом лес выдерживает достаточно жестко, жертвуя лишь наиболее старыми и хрупкими участками, которые, даже если они превращаются после пожара в абсолютно пустые пространства, в условиях непосредственной близости неповрежденного леса восстанавливаются сравнительно быстро.
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
12.3. Стабильность и устойчивость экосистем
Важнейшими показателями динамики экосистем являются устойчивостьистабильность. Определение устойчивости экосистем уже давалось — это способность экосистемы возвращаться в исходное состояние после снятия внешнего воздействия, выведшего ее из равновесия. Под стабильностью экосистемы понимают ее способность сохранять свою структуру и функциональные свойства при воздействии на нее внешних факторов. Иногда понятия устойчивость и стабильность рассматриваются как синонимы, но тогда следует различать два вида устойчивости:резидентная устойчивость(стабильность) — способность оставаться в устойчивом (равновесном) состоянии под нагрузкой, иупругая устойчивость(собственно устойчивость) — способность быстро восстанавливаться при снятии нагрузки. В разных терминах имеются различные смысловые оттенки, которые нужно учитывать.
Системы с высокой резидентной устойчивостью способны воспринимать значительные воздействия, не изменяя существенно своей структуры, то есть практически не выходя за пределы равновесного состояния. Поэтому понятие упругой устойчивости для них не определено (если система не выходила за пределы равновесия, то как можно говорить о возвращении в равновесное состояние после снятия возмущения). Если внешнее воздействие превышает определенные критические значения, то такая система обычно разрушается. В технике подобное качество называется жесткостью. Предельные значения внешних воздействий, которые система способна выдержать без разрушения соответствуют запасу жесткости. Когда говорят о высокой резидентной устойчивости, то имеется в виду именно высокий запас жесткости данной системы. Это несколько отличается от понятия высокой стабильности, так как здесь в первую очередь внимание обращается на неизменность структуры. Тундра, например, обладает высокой стабильностью, но она очень ранима, у нее малый запас жесткости, то есть малая резидентная устойчивость. Экосистему тундры очень легко разрушить. Достаточно проехать вездеходу. Колеи, которые он оставляют за собой, сохраняются десятилетиями. Такие экосистемы по аналогии с техникой можно назвать хрупкими.
Системы с малой резидентной устойчивостью для нормального существования должны обладать высокой упругой устойчивостью. Они более чувствительны к внешним возмущениям, под действием которых они как бы «прогибаются», частично деформируя свою структуру, но после снятия или ослабления внешних воздействий быстро возвращаются в исходное равновесное состояние. При превышении пороговых воздействий такая система теряет устойчивость, то есть все дальше удаляется от состояния равновесия. Диапазон воздействий, которые может выдержать система без разрушения, в технике соответствует запасу упругости. Таким образом, степень упругой устойчивости можно оценить как упругостью, определяющей степень сопротивления внешнему воздействию и скорость возврата в исходное состояние после снятия воздействия, так и запасом упругости. В отличие от упругих систем, пластичные системы после снятия внешнего воздействия не возвращаются в исходное состояние, а приходят к какому-то другому равновесному состоянию. Так согласно точке зрения оппонентов теории моноклимакса, для экосистем характерно не одно, а несколько состояний равновесия (климакса). Таким образом, для пластичных экосистем характерна малая упругая и малая резидентная устойчивость.
Похоже на то, что резидентная и упругая устойчивости взаимоисключают друг друга, точнее, экосистеме трудно развивать оба вида устойчивости. Например, одни леса состоят из деревьев с толстой корой, обладающих повышенной резидентной устойчивостью к пожарам. Но если такой лес все-таки сгорит, то его восстановление, как правило, крайне проблематично. Напротив, многие леса очень часто горят (низкая резидентная устойчивость), но быстро восстанавливаются (высокая упругая устойчивость). Ориентация экосистем на один из видов устойчивости определяется, как правило, изменчивостью среды: при стабильных условиях экосистемы склонны к более высокой резидентной устойчивости, при изменчивых условиях предпочтение отдается упругой устойчивости.
Рис. 12.3
Природа обоих видов устойчивости состоит в наличии в экосистеме разного рода обратных связей. В основном это отрицательные обратные связи, которые направлены на стабилизацию параметров экосистемы, возвращая их значения к какой-то изначально заданной величине. Однако немаловажную роль играют и положительные обратные связи, усиливающие благоприятные для системы изменения, например, в плане роста и выживаемости организмов. Однако деятельность положительных обратных связей обязательно должна быть ограничена соответствующими отрицательными обратными связями, иначе ничем не контролируемая экспансия жизни может привести экосистему к гибели. В обоих случаях схема управления выглядит одинаково (рис.25).
Механизмы реализации обратных связей называются механизмами гомеостаза(от греческих слов homoios — подобный, одинаковый и ststis — состояние). Обычно они достаточно сложны, так как их компоненты связаны в единые информационные сети факторами среды, а также различными физическими и химическими агентами – «посредниками», подобно тому, как нервная или гормональная системы связывают в одно целое части организма. Примером может служить сильная реакция организмов в экосистеме на слабые концентрации некоторых веществ. С энергетической точки зрения можно выделить одну закономерность, о которой уже говорилось ранее: обладатели высококачественной энергии, оставаясь в меньшинстве, управляют большими потоками энергии более низкого качества. Например, хищники управляют численностью травоядных животных. В то же время активность хищников и их численность управляется численностью их жертв по цепи обратной связи. По этой цепи на более высококачественный энергетический уровень (к хищникам) подается небольшая часть низкокачественной энергии — хищники уничтожают не всех жертв, а лишь небольшой их процент. Причем влияние этой части энергии на управление всей системой существенно усиливается деятельностью хищников.
Помимо отрицательной обратной связи устойчивость экосистемы может быть обеспечена избыточностью компонентов. Например, если в системе имеется несколько автотрофов, каждый из которых характеризуется своим температурным диапазоном функционирования, то скорость фотосинтеза сообщества в целом может оставаться неизменной, несмотря на колебания температуры. То же самое можно сказать и про гетеротрофов. Поэтому видовое разнообразие способствует повышению устойчивости экосистем (закон Эшби). При этом как в случае резидентной, так и в случае упругой устойчивости расширяется диапазон пороговых воздействий, которые способны выдержать экосистема без разрушения (запас жесткости и запас упругости).
Так, например, для тундры с ее бедным разнообразием характерны такие проявления низкой упругой устойчивости, как периодические резкие увеличения численности мелких грызунов — леммингов. О малой резидентной устойчивости тундры мы уже говорили. Экосистема тундры очень ранима и очень долго восстанавливается. В то же время тропические леса с их богатым разнообразием способны достаточно быстро залечивать свои раны. Легенды повествуют о целых городах, поглощенных джунглями, население которых не могло справиться с их натиском и вынуждено было уходить в другие районы.
Если разные стадии сукцессии экосистемы считать квазистатическими состояниями, то здесь можно отметить ряд закономерностей. На первых стадиях сукцессии экосистемы способны очень быстро возобновляться. Например, мы можем распахать по весне все поле, но уже к осени оно будет укрыто травами. Это говорит о достаточно высокой упругой устойчивости на начальных стадиях сукцессии, которая по мере движения к климаксу постоянно снижается. То есть на каждой следующей стадии для восстановления исходного состояния требуется гораздо больше времени. Климаксный лес характеризуется очень малой упругой устойчивостью.
Для резидентной устойчивости тенденция обратная. На первых стадиях сукцессии резидентная устойчивость остается достаточно невысокой — системы очень чувствительны к внешним воздействиям, откликаясь на них повышенным динамизмом. Например, заросли кустарника или молодой березняк может быть легко поврежден вездеходом (малая резидентная устойчивость), однако вторичная сукцессия достаточно быстро «затянет» повреждение (высокая упругая устойчивость). По мере роста видового разнообразия резидентная устойчивость непрерывно растет. При этом растет одновременно и жесткость системы (стабильность) и запас жесткости (взрослый лес вездеход так просто не одолеет). Однако по мере приближения к климаксу, как уже было показано, экосистема становится более чувствительна к действию некоторых катастрофических факторов среды (бури, пожары и т.п.). То есть на фоне продолжающегося роста стабильности (жесткости), запас жесткости экосистемы при движении ее к климаксу, по-видимому, начинает уменьшаться, что можно объяснить преобладанием в ней старых деревьев. Это говорит о том, что она становится все более хрупкой.
Как уже было сказано, для реальных лесов характерен, как правило, мозаичный климакс. По-видимому, именно такой лес имеет наилучший комплекс показателей устойчивости и стабильности. Периодические сильные воздействия на него в целом лес выдерживает достаточно жестко, жертвуя лишь наиболее старыми и хрупкими участками, которые, даже если они превращаются после пожара в абсолютно пустые пространства, в условиях непосредственной близости неповрежденного леса восстанавливаются сравнительно быстро.
Примером абсолютно нежизнеспособных экосистем являются агроценозы, создаваемы человеком из однолетних и двулетних культурных растений, то есть продукты сельскохозяйственной деятельности человека. Многие экологи даже не считают их экосистемами, хотя есть все основания считать их экосистемами с искусственно поддерживаемыми начальными стадиями сукцессии. Заброшенные поля сразу же втягиваются в естественный ход сукцессии. Что касается возделываемых полей, то они существуют только за счет человека. Свидетельством их нежизнеспособности является крайне малая устойчивость как упругая, так и резидентная, что является следствием очень скудного видового разнообразия как флоры (все сорняки подавляются гербицидами), так и фауны («вредных» насекомых мы травим пестицидами). Малая упругая устойчивость проявляется, например, в резких вспышках численности конкретных видов вредителей. Подобные вспышки в естественной природе наблюдаются очень редко. Малая резидентная устойчивость проявляется, например, в повышенной чувствительности посевов к природным условиям, таким как град, засуха, повышенное количество осадков и т.д.
Осваивая все большие территории планеты, человек продолжает стратегию обеднения видового разнообразия. Этим он все более подрывает способность природы сопротивляться внешним воздействиям и возвращаться в исходное состояние. Как знать, может быть, роковая черта, за которой начинаются необратимые изменения, уже пройдена. Это значит, что биосфера никогда больше не придет к исходному состоянию. Это значит, что мы вступили на путь глобальных перемен, которые постепенно изменят жизнь на Земле самым коренным образом. Будет ли в этом мире место человеку?
ПОВТОРИМ
Охарактеризовать циклическую и поступательную динамику экосистем.
Экзодинамические и эндодинамические сукцессии.
Климаксные и моноклимаксные экосистемы.
Первичная и вторичная, автотрофные и гетеротрофные сукцессии.
Закономерности сукцессионного процесса.
Влияние внешних, в том числе и антропогенных воздействий на динамику климаксных систем.
Охарактеризовать виды устойчивости экосистем.
studfiles.net
Устойчивость экосистемы — материалы для подготовки к ЕГЭ по Биологии
Автор статьи — Л.В. Окольнова.
Устойчивость экосистемы не постоянна. Со временем все меняется — там, где были реки и леса, появляются пустыни.
Однако, в течение определенного и достаточно длительного времени экосистема устойчива и у этой устойчивости есть определенные критерии.
Критерий №1
Поток энергии и круговорот веществ.
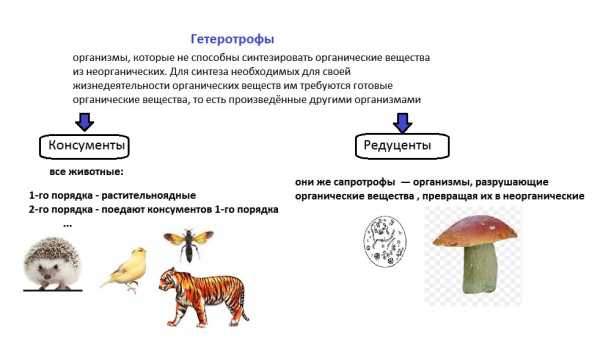
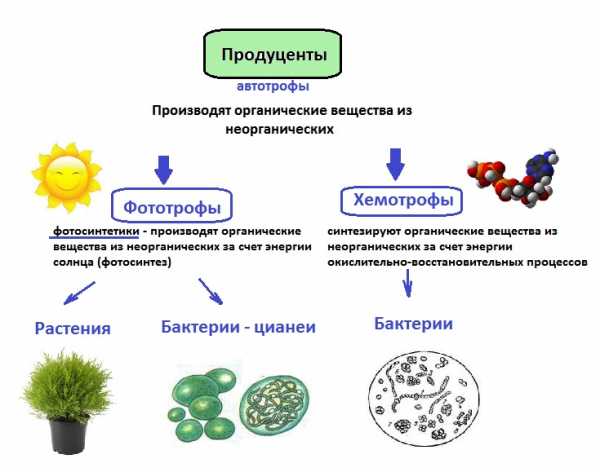
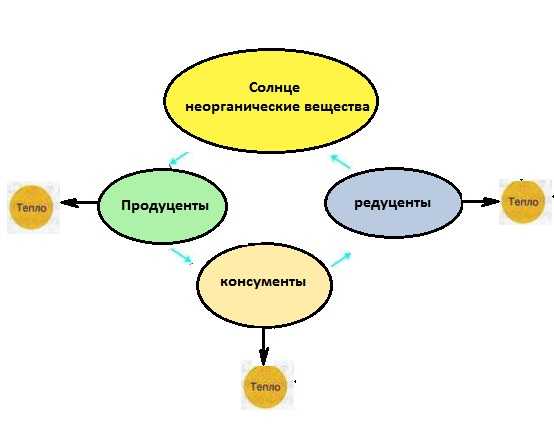
Откуда берется энергия? Растения (продуценты) получают солнечную энергию и неорганические вещества, превращают это в энергию соединений — органических веществ.
Дальше по известной схеме трофических сетей органические вещества переходят с одного уровня на другой и поступают к редуцентам.
Задача редуцентов — вернуть элементы соединений в окружающую среду — органические вещества перерабатываются в неорганические.
Критерий №2
Видовое разнообразие.
Чем больше видов на каждом трофическом уровне, тем выше устойчивость. Логика здесь очень простая — если исчезнет какой-то один вид, то от этого исчезновения не должен зависеть трофический уровень — кто-то встанет на его место.

Критерий №3
Саморегуляция.
Самый загадочный критерий. Как организм залечивает раны, так и экосистема способная восстанавливаться. Природные катаклизмы, естественные процессы и человек могут серьезно повлиять на устойчивость — животные могут мигрировать, погибать, пищевые связи “трещат” по швам”, но проходит время и система самовосстанавливается.

И все же экосистемы вечными не бывают.

Давайте рассмотрим примеры смен экосистем (кстати, в ЕГЭ эти вопросы встречаются в части С).
Каменистая экосистема -> лесной массив
Абиотические разрушающие факторы: свет, выветривание, влажность;
Биотические разрушающие факторы: бактерии, лишайники, водоросли, грибы;
Создаются органические вещества, которые способствуют образованию почвы.
Первыми ее заселят неприхотливые организмы — лишайники ( это вообще “пионеры” экосистем) и мхи.
Затем появятся травянистые формы, потом кустарники и затем уже деревья.

Сукцессия водоема
Водоем зарастает зелеными водорослями. Образуется “ковер”, другим растениям и животным не хватает кислорода и постепенно водоем заболачивается. Но, появляются организмы. приспособленные к такой среде. Появляется все больше травы, потом появляются кустарники — появляется мокрый луг, где свободно могут прорастать ягоды, грибы, обитать птицы. Еще несколько лет, и на месте водоема появляется лес.


Звоните нам: 8 (800) 775-06-82 (бесплатный звонок по России) +7 (495) 984-09-27 (бесплатный звонок по Москве)
Или нажмите на кнопку «Узнать больше», чтобы заполнить контактную форму. Мы обязательно Вам перезвоним.
ege-study.ru
Глава 6. Стабильность и устойчивость водных экосистем
Проблема стабильности и устойчивости экосистем и входящих в их состав сообществ организмов — одна из важнейших в современной экологии. Она активно обсуждается, и к настоящему времени в литературе накопилось достаточно большое количество различных, часто противоречивых представлений о стабильности и устойчивости экосистем. Например, считается, что существовать в течение длительного времени могут лишь устойчивые экосистемы и пределы их устойчивости определяются максимальными нагрузками, которые они могут выдержать, не нарушаясь. Одни авторы стабильность и устойчивость рассматривают как синонимы (Одум, 1986, Недорезов, Сидько, 1995), другие с их помощью описывают разные состояния экосистем. Рассмотрим некоторые наиболее часто встречающиеся представления об этих важнейших свойствах биологических систем и экосистем.
Стабильность экосистем возникает в результате внутренних взаимодействий (Margalef,1968),она требует внутривидовых взаимоотношений, в то время как межвидовые взаимодействия приводят к дестабилизационным влияниям (Yodzis, 1981) и являются мерой изменчивости системы (Риклефс, 1979). Стабильность часто связывают с сложностью систем — более сложно организованные системы — более стабильны (MacArthur,1955; Voris et al.,1980). Сложность структуры сообществ организмов, оцениваемую их разнообразием, многие исследователи связвают с их стабильностью: чем разнообразнее система, тем она стабильнее. Такие системы контролируются биотическими факторами. Под влиянием антропогенных факторов и эвтофировании водоемов разнообразие и стабильность систем уменьшаются. Однако стабильность экосистем нельзя связывать только с их разнообразием. По мнению Мэя (May,1972,1987) увеличение числа элементов в системе ведет к ее нестабильности, т.е. к несоответствию реакции системы на незначительную смену условий.
Стабильность экосистем может быть кратковременной (сукцессионной) и долговременной (эволюционной). При этом в соответствии с гипотезой эволюционного времени следует ожидать прямое соответствие между разнообразием экосистем и их возрастом (Thiery,1982).Так, например, на ранних стадиях развития озер их видовое разнообразие низкое, а затем возрастает (Goulden, 1973). Согласно другой точке зрения, олиготрофные озера более организованы по сравнению с эвтрофными, которые более продвинуты и отличаются меньшим разнообразием (Margalef, 1964; Reed,1978).
Другое понимание стабильности — способность их сохраняться неизменными, несмотря на воздействие нарушающих факторов. Стабильность рассматривается как способность системы сохранять относительно неизменное состояние под влиянием некатострофических явлений (Заугольнова, 1985) и противостоять изменениям (абиотических и биотических воздействий окружающей среды), сохраняя динамическое равновесие (гомеостаз) (Дедю,1989). Приведенные точки зрения на стабильность экосистем по сути дела не принимают во внимание связи между их структурными и функциональными характеристиками.
Структура систем, не подверженных антропогенным воздействиям, меняется во времени в зависимости от изменений внешних и внутренних факторов. Это эволюционные процессы. Так в процессе эволюции озер происходят постепенные изменения структуры и функционирования их экосистем. Аналогичным образом структурные изменения и связанные с ними функциональные происходят при антропогенных воздействиях, например, эвтрофировании, загрязнении, ацидофикации. В этих случаях процессы структурных и функциональных изменений протекают с большими скоростями. Связь структурных, функциональных характеристик и временного фактора учитывается при понимании стабильности, как обобщенного свойства живой системы, которое характеризует нормальное ее развитие по генетической программе в конкретных экологических условиях и она обусловлена общей структурно-функциональной организацией системы на протяжении всего времени ее существования (Голубець, Царик, 1992).
Устойчивость чаще всего связывают со способностью систем выдерживать изменения, вызванные влиянием извне и возвращаться в исходное состояние после воздействия на них ( Margalef, 1968; Риклефс, 1979; Одум,1986; Голубець, Царик, 1992; Заугольнова и др.,1992;Краснощеков, Розенберг, 1992; Галковская,1995; Михайлов, Эдельштейн, 1996), сохраняя свою структуру и функциональные особенности (Дедю, 1989), сохраняя «лицо», зависящее от предыстории системы (Федоров, 1977). Некоторые авторы рассматривают устойчивость как способность сообществ к саморегуляции, основанную на возможности адаптироваться к изменениям внешних условий (Шарашова,1989) и как меру их чувствительности к нарушениям, сохраняя свои особенности во времени (Бигон и др. 1989). При этом различают упругую устойчивость — меру скорости возврата системы в исходное состояние и устойчивость сопротивления — способность системы избегать изменений. В. Г. Драбкова (1994) рассматривает устойчивость к конкретным воздействиям, а не устойчивость вообще и в зависимости от характера воздействия система может быть устойчива, менее устойчива и неустойчива.
Каждая экосистема и входящие в нее сообщества организмов адаптированы к сезонным, годовым изменениям факторов внешней среды. Это выражается в флюктуациях значений их структурных и функциональных характеристик таких, например, как видовой состав, разнообразие, биомасса, численность, продукция, траты на обмен относительно некоторых средних значений за эти периоды времени. Поэтому обычно сообщества бентосных или планктонных организмов в разных по типу и географическому положению водоемах или водотоках характеризуются средними за год или вегетационный сезон величинами биомасс или численности. Сохранение такого среднего уровня, видового состава и разнообразия отражает стабильность экосистемы во времени.
Многие исследователи, как уже говорилось ранее, рассматривают устойчивость экосистем и сообществ организмов как их способность возвращаться в исходное состояние после воздействий на них. При этом часто проводится аналогия с устойчивостью шарика, который возвращается в исходную точку. Такой механистический подход к определению устойчивости экосистем и сообществ организмов не правомочен. Эти системы, изменившие свои структуру под влиянием факторов среды, в том числе и антропогенных, после прекращения их воздействия не могут вернуться к первоначальной структуре уже потому, что часть видов организмов в результате таких воздействий может быть просто уничтожена. В отличие от физических тел, не меняющих свою структуру при выведении их из состояния равновесия и другого воздействия ( если не произошло их полного разрушения), экосистемы и сообщества организмов чаще всего изменяют свою структуру, а значит и функционирование под влиянием меняющихся факторов среды, когда сила их превышает адаптационные возможности, входящих в их состав видов. Таким образом, нельзя рассматривать устойчивость экосистем с позиций оценки устойчивости механических систем. Для популяций и, как можно предположить, для экосистем, характерно не одно, а несколько состояний равновесия и после стрессовых воздействий они часто возвращаются не в то состояние равновесия, из которого были выведены, а в другое (Holling,1973).
Флуктуации структурных и функциональных характеристик в течение года или вегетационного сезона относительно их средних за эти периоды значений отражают устойчивость системы. Таким образом, устойчивость системы может быть охарактеризована размахом колебаний в течение года или вегетационного сезона ее качественных и количественных показателей. Чем больше размах этих колебаний, тем менее устойчива система и, наоборот, с уменьшением размаха возрастает устойчивость системы.
Под устойчивостью сообщества или экосистемы предлагается понимать отклонение их характеристик от некого среднего уровня, свойственного конкретной системе как исторически сложившейся при определенных условиях. В качестве меры устойчивости системы можно рассматривать вариабельность динамики биомассы (ВДБ), которая была подробно рассмотрена в главе 2. Этот показатель значим уже потому, что с биомассой связаны все функциональные и некоторые структурные характеристики экосистем и сообществ организмов.
По мере упрощения структуры сообществ и экосистем возрастает амплитуда сезонных колебаний их функциональных характеристик, в том числе ВДБ. Наиболее устойчивы сообщества и экосистемы с более высоким разнообразием, в которых доминируют стенобионтные виды (Алимов, 1989):
St = 0.045*e 0.51H , R2 = 0.98, (54)
St = 0.04*e 3.641(с/э) , R2 = 0.94, (55)
Устойчивость сообществ и экосистем находится в прямой зависимости от прозрачности воды в водоемах (Алимов, 1989). Это имеет принципиальное значение, так как первичная продукция планктона находится в обратной зависимости от прозрачности воды. Таким образом, при увеличении продуктивности или степени эвтрофирования водоемов или водотоков уменьшается устойчивость экосистем и их составляющих.
Устойчивость сообществ гидробионтов и экосистем меняется так же при изменении степени эксплуатации их. Так увеличение пресса рыб в озерах — питомниках приводило к снижению устойчивости планктонных и бентосных сообществ (табл.19).
Таблица 19
Устойчивость (Bmin/Bmax) разных сообществ в озерах — рыбопитомниках Б,Окуненок и Щучье при разной посадке рыб(х103экз./га)
Год | посадка рыб | фитопланк. | Коловратки | планкт. ракообр. | Макробент. | Мейобентос | вся экосист. |
Б.Окунен | |||||||
1986 | 31.5 | 0.01 | 0.21 | 0.052 | 0.005 | 0.11 | 0.26 |
1987 | 14.5 | 0.022 | 0.13 | 0.30 | 0.055 | 0.09 | 0.20 |
1988 | 10.0 | 0.071 | 0.04 | 0.35 | 0.27 | 0.38 | 0.35 |
Щучье | |||||||
1981 | 20 | 0.14 | 0.051 | 0.062 | 0.34 | — | — |
1982 | 62 | 0.06 | 0.008 | 0.044 | 0.62 | — | — |
В оз. Б. Окуненок подращивались как карпы, так и пелядь, но главным образом карпы, оз. Щучье использовалось как маточный водоем для подращивания пеляди. Такие различия в использовании озер, как видно из приведенной таблицы, отразились на устойчивости экосистем озер и особенно сообществ гидробионтов. Манипуляции с целыми озерными экосистемами показали, что изменения структуры сообществ рыб приводят к таким же изменениям первичной продукции которые вызываются физико-химическими факторами (Carpenter, 1989). Исследование детерминистских моделей озерных экосистем показало, что вариабельность численности и биомассы хищных рыб, как верхнего звена трофической цепи, каскадируемая на нижние уровни, может вызывать вариабельность первичной продукции, которую невозможно объяснить только воздействием физических и химических факторов, а зависит в большой степени от конфигурации и динамики пищевых цепей (Carpenter, Kitchel,1987). При этом важно, что внутригодовые вариации первичной продукции возрастали при увеличении вариабельности плотности населения рыб.
Возрастание биогенной нагрузки на водоем приводит к его эвтрофированию, с которым связано увеличение общей концентрации органического вещества, ВОВ, РОВ и возрастание размаха колебаний соотношения между растворенной и взвешенной формой органического фосфора, усиление роли бактерий в деструкции органического вещества, снижение удельной эффективности биотического круговорота органических веществ а так же увеличение размаха колебаний концентраций этих форм органического вещества (Умнов, 1997), Последнее говорит о том, что эвтрофирование приводит к снижению устойчивости экосистемы водоема. При этом в ряду озер с повышающейся трофностью устойчивость снижается. Например, в системе озер Нарочь — Мястро — Баторино годовые величины St для фитопланктона в среднем за период с 1968 по 1980 г.г. были 0.25, 0.1, 0.08, соответственно. При этом амплитуда их межгодовых колебаний возрастала в том же направлении.
Потоки вещества не отделимы от потоков энергии и информации в экосистемах. Понятно, что реальные экосистемы не могут иметь безотходные, предельно замкнутые циклы — часть биогенов постоянно теряется и выходит из круговорота. В то же время некоторые организмы способны пополнять запасы минеральных веществ путем разложения различных веществ, в том числе и материнских пород. Это особенно относится к микроорганизмам. Эти качества делают экосистемы удивительно стабильными во времени. Способность экосистемы сохранять свои характерные свойства обусловлена присутствием в экосистеме биотической составляющей, вступающей в сложные связи с абиотической частью системы, трансформируя ее, и оказывая на нее влияние таким образом, чтобы компенсировать возможные возмущения (Куприянова, 1983). Эта стабильность проявляется в первую очередь в постоянстве видового состава и структуры сообществ и экосисистемы. Так, например, в наименее продуктивном в системе нарочанских озер, оз. Нарочь индекс Шеннона (рассчитанный по данным: «Многолетние показатели …»,1973) в период с 1955 по 1970 год менялся незакономерно от 2.81 до 3.81 бит/экз, составляя в среднем 3.44 ( коэффициент вариации — 13.3). В наиболее продуктивном — оз. Баторино этот индекс в течение того же периода времени менялся от 2.55 до 3.45., в среднем — 3.08 бит/экз. (коэффициент вариации — 48.98). И. Н. Андронникова (1980, 1996), анализируя результаты многолетних исследований сообществ зоопланктона в различных озерах, подчеркнула, что при естественном развитии озер авторы отмечают поразительную стабильность видового состава сообществ, который может сохраняться в водоемах десятилетиями и даже столетиями. Биоразнообразие стабилизирует процессы в сообществе и экосистеме, но не популяционные процессы (Tilman,1996). Существенно, что так же мало меняется и устойчивость в сообществах гидробионтов в малопродуктивных озерах, по сравнению с более эвтрофными. В сообществах фитопланктона оз. Нарочь значения St в 1968-1970. так же как и 1976-1980 г.г. составляли 0.25, зоопланктона в период 1955 — 1958 — 0.18, 1959 — 1971 — 0.22, 1976 -1979 г.г. — 0.2. В оз. Широ (Хакасия), мало подвергавшееся антропогенным воздействиям, St в сообществах зоопланктона ( рассчитано по данным Дегерменджи с соавторами, 1996) в 1950 г. составляло 0.3, в 1975 г. — 0.25, что с учетом возможных ошибок практически одинаково. Чем выше видовое богатство, тем ниже коэффициент вариации биомассы сообщества (Tilman, 1996).
По мере эвтрофирования озер происходят постепенно структурные изменения, которые выражаются не столько в смене видового состава, сколько в замене доминирующих видов и роста их численности. Так, в процессе эволюции озер происходят постепенные изменения структуры и функционирования их экосистем. Аналогичным образом структурные изменения и связанные с ними функциональные имеют место при антропогенном эвтрофировании или загрязнении. В этих случаях процессы структурных и функциональных перестроек протекают с большими скоростями.
Предлагается следующая трактовка стабильности экосистем и сообществ организмов. В конкретных условиях среды со свойственной им сезонной изменчивостью, определяющейся особенностями географического положения, складываются определенные по структуре экосистемы. Средние за год или вегетационный сезон значения структурных и функциональных характеристик этих систем остаются неизменными в течение достаточно продолжительного времени. Это свидетельствует о стабильности экосистемы в конкретных условиях. Именно так следует рассматривать и оценивать стабильность экосистем и сообществ организмов, в них входящих. При действии определенных факторов среды конкретная экосистема может оставаться стабильной до тех пор, пока воздействия отдельных факторов находятся в пределах, к которым она исторически адаптирована. При изменении качества внешних факторов или силы их воздействия, превышающей адаптационные возможности организмов, например, при поступлении загрязнения или других антропогенных нагрузках, к которым данная система не адаптирована, ее структура и функционирование изменятся и система перейдет в другое состояние с новыми значениями структурных и функциональных характеристик. Она будет оставаться стабильной до тех пор, пока будут действовать данные факторы.
Для оценки возможных изменений в сообществах гидробионтов и экосистем под влиянием антропогенных факторов введем понятие выносливости. Под выносливостью будем понимать их способность противостоять изменениям внешних условий. В качестве меры выносливости в первом приближении можно использовать первую производную функции, описывающую связь между степенью сложности системы, оцениваемую индексом разнообразия, и конкретным фактором среды. В качестве примера рассмотрим влияние такого важнейшего и интегрированного фактора как биологическое потребление кислорода (например, БПК5). Как было показано выше, зависимость индекса разнообразия для сообществ планктона и бентоса описывается уравнением (1):
Н = 3.741*БПК5 — 0.269
Первая производная этого уравнения:
dH/dБПК5 = — 1.006*БПК5 — 1.269 .
Поскольку производная возрастает по мере снижения БПК5 , следует ожидать более медленных изменений структуры, а значит и функционирования системы в более продуктивных или более загрязненных водах. В малопродуктивных чистых или слабо загрязненных водах небольшие изменения БПК5 приводят к существенным изменениям структурных показателей экосистем, и, наоборот, в более загрязненных и более продуктивных водах значительные изменения БПК5 приводят к меньшим изменениям структуры (табл. 20).
Таблица 20
Выносливость экосистем разной продуктивности
Трофность водоемов | БПК5 | dН/dБПК5 , бит/ккал |
ультраолиготрофные | 2.11 — 6.13 | (-0.39) — (-0.101) |
Олиготрофные | 6.13 — 12.71 | (-0.101) — (-0.04) |
Мезотрофные | 12.71 — 28.27 | (-0.04) — (-0.014) |
Эвтрофные | 28.27 — 58.64 | (- 0.014) — (-0.006) |
Гиперэвтрофные | 58.64 — 92.82 | (-0.006) — (- 0.003) |
Таким образом, более продуктивные системы оказываются менее устойчивыми и более выносливыми. При этом наиболее выносливы системы с малой устойчивостью, поскольку они исторически адаптированы к значительным сезонным и межгодовым колебаниям факторов. В таких системах преобладают виды с широкими экологическими спектрами (эврибионтные). Наименее выносливы системы, адаптированные к меньшим колебаниям внешних условий, в них преобладают узкоспециализированные виды. В этом отношении представляет интерес исследование динамики биоценозов с применением методов математического моделирования и методов статистической механики (Алексеев и др., 1978). Оно показало, что максимальное видовое разнообразие может быть получено, когда консументы трофически специализированы. При эврифагии косументов резко возрастает размер областей, в которых существуют автоколебательные и триггерные режимы. С увеличением видового разнообразия амплитуда колебаний биомасс как хищников так и жертв убывает. Эти заключения существенно усиливают высказанные мною положения, которые, как было показано, имели и экспериментальное подтверждение.
С этих позиций можно объяснить, почему тропические, арктические, антарктические сообщества и экосистемы, а так же экосистемы подземных вод, горячих источников, эфемерных водоемов и т.п. очень уязвимы и легко разрушаются. Они обладают меньшей выносливостью по сравнению, например, с экосистемами умеренных широт, которые более выносливы и способны выдерживать значительные антропогенные нагрузки.
Приняв во внимание все сказанное, нет основания говорить о стабильных и нестабильных экосистемах. Экосистема находится в стабильном состоянии, пока на нее с постоянной силой воздействуют конкретные факторы среды. Она характеризуется конкретными структурными и функциональными характеристиками, устойчивостью.
В сообществах гидробионтов в течение года или вегетационного сезона из-за сезонных циклов развития организмов может происходить смена видов доминантов, что наиболее заметно в сообществах, организованных видами с короткими циклами развития, например в сообществах планктона. Поэтому определять стабильность системы более надежно по средним за сезон или год значениям структурных и функциональных характеристик.
Как уже говорилось, при изменении силы или качества воздействия на систему она переходит в новое состояние, характеризующееся новыми структурными и функциональными показателями. Переход системы из одного стабильного состояния в другое стабильное состояние обусловлен изменениями условий среды, в том числе и поступления энергии, и происходит не в один момент, а в течение некоторого промежутка времени.
Сказанное достаточно хорошо может быть представлено в виде «фазовых портретов», отражающих динамику функции при изменении аргумента. Такой метод анализа сообществ организмов дает возможность выявить стационарные состояния системы и характер её динамики при отклонении от них. При этом под стационарным состоянием понимается состояние с нулевой скоростью изменения параметра.
Такой подход к анализу сложной и динамичной системы, какой и является сообщество организмов или экосистема, наиболее активно используется при исследованиях динамики планктонных и ихтиоценозов (Терещенко, Надиров, 1996; Терещенко, Вербицкий, 1997).В исследованиях этих авторов использовались фазовые портреты dH/dt = f(H), где Н — индекс Шеннона, t — время.
Последовательное применение и анализ фазовых портретов сообществ зоопланктона и рыб при воздействии различных факторов среды (вселение рыб, влияние хищников, ацидофикации, солей тяжелых металлов, пестицидов и т.д.) позволило В. Т. Терещенко и В. Б. Вербицкому выявить пять типов структурных фазовых портретов.
Такой подход к пониманию стабильности, устойчивости, выносливости экосистем и сообществ гидробионтов создает возможность разработки способов расчета ожидаемых усилий при желании или необходимости вывести систему из одного стабильного состояния, перевода и поддержания ее в другом стабильном состоянии. Например, при желании получить определенный экономический эффект от удобрения водоемов, удаления из них сорных рыб и заселения их ценными видами рыб, при прудовом рыбоводстве и т.п. Однако для этого необходимо проведение ряда мероприятий, направленных на поддержание системы в новом состоянии, резко отличающемся от исторически сложившегося. Для поддержания такого состояния следует непрерывно прилагать вполне определенные усилия, внешние по отношению к экосистеме, которые в наиболее общей форме могут быть оценены в виде затрат энергии. Аналогичным образом следует оценивать усилия при других воздействиях на экосистемы при их хозяйственном использовании или при деэвтрофикации или деацидофикации водоемов. В первом приближении можно ожидать, что для перевода экосистемы водоема из эвтрофного состояния в мезотрофное на единицу информации должно быть затрачено большее количество энергии, чем из мезотрофного в олиготрофное.
Антропогенные воздействия на экосистемы могут приводить либо к их упрощению, что наблюдается, например, при стрессовых на них воздействиях со всеми вытекающими последствиями, либо к усложнению. В последнем случае их сложность и разнообразие возрастают. Это может иметь место при восстановлении нарушенных ранее систем или, например, при разумной акклиматизации организмов. В случаях антропогенного увеличения сложности экосистем такие воздействия внешние по отношению к экосистеме, выступают как фактор, влияющий на ход сукцессионного процесса или запускающий его. Так усиление пресса рыб как результат увеличения числа их видов и количества в озерах-питомниках изменяет структуру пищевых цепей и продуктивность сообществ донных животных, блокирует основные направления сукцессионного процесса и возвращает сообщество на ранние стадии сукцессии (Голубков, 1997).
Если стрессовые воздействия на экосистемы привлекают по понятным причинам наибольшее внимание, то количественных примеров антропогенного усложнения экосистем водоемов в настоящее время очень мало. Однако высказанные мною положения и сделанные расчеты позволяют надеяться на успех и в этом направлении. Надо сказать, что возможность интродукции в сообщество дополнительного вида будет всегда существовать, если с одним или несколькими существующими видами он образует связь. При этом следует отметить, что система может быть стабильной, если, по мнению Мэя, произведение связности на число видов будет ниже 4 (Маргалеф, 1992).
studfiles.net
УСТОЙЧИВОСТЬ ЭКОСИСТЕМЫ — это… Что такое УСТОЙЧИВОСТЬ ЭКОСИСТЕМЫ?
- УСТОЙЧИВОСТЬ ЭКОСИСТЕМЫ
- УСТОЙЧИВОСТЬ ЭКОСИСТЕМЫ
- способность экосистемы к реакциям, пропорциональным по величине силе воздействия, которые гасят эти воздействия. При этом в экосистеме возбуждаются компенсационные (отрицательные) обратные связи, что равноценно выполнению принципа Ле Шателье. При превышении некоторой критической величины воздействия экосистема теряет устойчивость, возникают положительные обратные связи, которые могут привести к её разрушению. Син.: Живучесть экосистемы, Жизнестойкость экосистемы.
Экологический словарь, 2001
- Устойчивость экосистемы
-
способность экосистемы к реакциям, пропорциональным по величине силе воздействия, которые гасят эти воздействия. При этом в экосистеме возбуждаются компенсационные (отрицательные) обратные связи, что равноценно выполнению принципа Ле Шателье. При превышении некоторой критической величины воздействия экосистема теряет устойчивость, возникают положительные обратные связи, которые могут привести к её разрушению. Син.: Живучесть экосистемы, Жизнестойкость экосистемы.
EdwART. Словарь экологических терминов и определений, 2010
.
- УСТОЙЧИВОСТЬ УПРУГАЯ
- УТИЛИЗАЦИЯ (обезвреживание) МУСОРА И ОТХОДОВ
Смотреть что такое «УСТОЙЧИВОСТЬ ЭКОСИСТЕМЫ» в других словарях:
Устойчивость природных комплексов к техногенным нагрузкам — способность комплексов природной экосистемы противостоять антропогенным (техногенным) нагрузкам, которые нарушают их естественное функционирование. Степень подверженности экосистемы внешним факторам, нарушающим ее структуру и функционирование,… … Словарь черезвычайных ситуаций
УСТОЙЧИВОСТЬ ЭКОЛОГИЧЕСКАЯ — резистентность экологическая, способность экосистемы и ее отдельных частей противостоять колебаниям внешних факторов и сохранять свою структуру и функциональные особенности. Экологический энциклопедический словарь. Кишинев: Главная редакция… … Экологический словарь
Устойчивость окружающей среды — позволит поддерживать жизнь человека в нынешнем её виде. Снимок Blue Marble, NASA: 2001 (слева), 2002 (справа). Устойчивость окр … Википедия
Живучесть экосистемы — способность экосистемы выдерживать резкие изменения абиотической среды, массовые размножения или исчезновения отдельных видов, антропогенные нагрузки . Устойчивое (стационарное) состояние глобальной экосистемы сохраняется до тех пор, пока… … Словарь черезвычайных ситуаций
РЕЗИСТЕНТНАЯ УСТОЙЧИВОСТЬ — способность экосистемы оставаться в устойчивом состоянии под нагрузкой, ее невосприимчивость к внешнему воздействию. Экологический словарь, 2001 Резистентная устойчивость способность экосистемы оставаться в устойчивом состоянии под нагрузкой, ее… … Экологический словарь
УПРУГАЯ УСТОЙЧИВОСТЬ — способность экосистемы быстро восстанавливаться. Экологический словарь, 2001 Упругая устойчивость способность экосистемы быстро восстанавливаться. EdwART … Экологический словарь
УПОРЯДОЧЕННОСТЬ ЭКОСИСТЕМЫ — устойчивость внутренней структуры экосистемы во времени и пространстве, обусловленная устойчивостью трофических и пространственных связей между популяциями, а также устойчивым равновесием между экотопом и биоценозом. Упорядоченность экосистемы… … Экологический словарь
ЭЛАСТИЧНОСТЬ ЭКОСИСТЕМЫ — (от греч. elastikos упругий, гибкий, растяжимый), способность экосистемы менять свои динамические качества (сохраняя гомеостаз, устойчивость) в определенных пределах. См. также Валентность экологическая . Экологический энциклопедический словарь.… … Экологический словарь
Экосистема — Экосистема, или экологическая система (от др. греч. οἶκος жилище, местопребывание и σύστημα система) биологическая система, состоящая из сообщества живых организмов (биоценоз), среды их обитания (биотоп), системы связей,… … Википедия
Временные методические рекомендации по оценке на стадии ТЭО воздействия на окружающую среду (ОВОС) подземных сооружений для строительства в г. Москве — Терминология Временные методические рекомендации по оценке на стадии ТЭО воздействия на окружающую среду (ОВОС) подземных сооружений для строительства в г. Москве: 1. Безопасность экологическая состояние защищенности социальных, экономических,… … Словарь-справочник терминов нормативно-технической документации
dic.academic.ru
2.6. Устойчивость экосистем
Толерантность вида. Термин толерантность (от лат. tolerantia – терпение) означает выносливость вида по отношению к колебаниям какого-либо экологического фактора, или другими словами, способность организмов переносить отклонения экологических факторов среды от оптимальных для них величин. Изменения величин этих факторов для каждого организма допустимы только в определенных пределах, при которых сохраняется нормальное функционирование организма, т.е. его жизнеспособность. Допустимые пределы изменений экологических факторов среды называются границами толерантности. Разные виды организмов отличаются более широкими или более узкими границами толерантности. Чем большие пределы изменения параметров среды безболезненно выдерживает конкретный организм, тем выше толерантность, или устойчивость этого организма к изменению экологических факторов среды.
Адаптация организмов к изменению экологических факторов. Показатели устойчивости организмов в изменяющихся условиях среды обитания определяются возможностями организмов приспосабливаться (адаптироваться) к изменениям биотических и абиотических факторов. Адаптациями называются эволюционно выработанные и наследственно (генетически) закрепленные свойства организмов, обеспечивающие их нормальную жизнедеятельность при изменениях экологических факторов. Адаптационные возможности у разных видов очень сильно различаются. Например, береза хорошо растет как на сухих, так и увлажненных почвах, а сосна – только на почвах с умеренным увлажнением.
Часто важны не только пределы изменения экологических факторов, но и скорость их изменения, т.е. динамика. Не все виды способны приспособиться к быстрым изменениям условий среды. Виды, которые не могут (или не успевают) приспособиться к изменившимся условиям, вымирают и их экологические ниши в экосистемах занимают другие, более пластичные виды.
Рассмотрим основные виды адаптаций организмов к изменениям экологических факторов. Наиболее важными из них являются:
— морфологические;
— физиологические;
— поведенческие.
К морфологическим адаптациям относятся видоизменения органов, например, развитие у баобаба колючек вместо листьев, а у китов и дельфинов – плавников вместо ног. Физиологические адаптации связаны с особенностями ферментативного набора в пищеварительном тракте. Так, потребность животных во влаге удовлетворяется в пустынях путем биохимического окисления жиров, а у растений биохимические процессы фотосинтеза позволяют создавать органическое вещество из неорганических соединений. Поведенческие адаптации проявляются, например, в способах обеспечения теплообмена у птиц путем сезонных перелетов, у животных – с помощью линьки; для обеспечения пищей хищники используют приемы затаивания (в засаде), а их жертвы – защитную окраску.
Устойчивость экосистем – это способность экосистем сохранять структуру и нормальное функционирование при изменениях экологических факторов. Рассмотренные выше адаптации организмов к изменениям факторов среды обитания в определенной степени обеспечивают устойчивость экосистем, в состав которых они входят, к изменению экологических факторов среды. Однако, как и всякая более сложная система, экосистема по сравнению с отдельными видами организмов имеет более высокую степень надежности функционирования в изменяющейся среде, так как на системном уровне формируются и развиваются новые, системные механизмы обеспечения устойчивости и живучести экосистем, которые отсутствовали у отдельных видов. Такие эволюционно выработанные механизмы приспособления экосистем к изменениям среды обитания называются адаптациями экосистем.
Рассмотрим адаптации экосистем, состоящие из адаптационных механизмов двух уровней: видовой уровень и интеграционный, или системный уровень. Видовой (низший) уровень соответствует ранее рассмотренным механизмам в подразделе «Адаптации организмов к изменению экологических факторов». Системный уровень образуют приспособительные механизмы, возникающие за счет видового взаимодействия по трофическим цепям и сетям. Природа этих интеграционных, системных механизмов обеспечения устойчивости экосистем основана на круговороте веществ, который осуществляется с помощью трофических цепей.
Существование биогеохимических круговоротов создает возможность для саморегуляции экосистем (или гомеостаза), что придает экосистеме устойчивость в течение длительных периодов. Например, показателем устойчивости глобальной экосистемы, связанной с круговоротом веществ, может служить следующий факт. Известно, что 93% массы тела человека составляют 4 химических элемента: кислород, углерод, водород и кальций, которые, во-первых, входят в перечень одиннадцати самых распространенных в геосферах Земли химических элементов, и, во-вторых, эти четыре элемента сами образуют более 56% массы геосфер.
Видовое разнообразие – также один из факторов устойчивости экосистем к неблагоприятным факторам среды. Разнообразие обеспечивает как бы подстраховку, дублирование устойчивости. Например, малочисленный вид при неблагоприятных условиях для другого широко представленного вида может резко увеличить свою численность и таким образом заполнить освободившееся пространство (экологическую нишу), сохранив экосистему как единое целое. Такая последовательная смена видов или замена одного биоценоза другим называется сукцессией (от лат. сукцедо – следую).
Чтобы лучше уяснить суть сукцессии в экосистеме, рассмотрим два примера:
1) известно, что после лесного пожара сначала появляются лиственные породы, а затем через 70–100 лет их сменяют хвойные;
2) в упавшем дереве сначала поселяются короеды, затем появляются пожиратели древесины, а бактерии и грибы завершают процедуру превращения упавшего дерева в гумус почвы.
Таким образом, увеличение степени разнообразия является основой того, что экосистемы с более длинными цепями питания формируют более интенсивный круговорот веществ и, следовательно, обладают повышенной устойчивостью благодаря возможностям саморегуляции (гомеостаза).
Гомеостаз. Природные экосистемы (например, лесные, степные) существуют в течение длительного времени и обладают определенной стабильностью, для поддержания которой необходима сбалансированность потоков вещества и энергии в процессах обмена между организмами и окружающей средой. Однако абсолютной стабильности в природе не бывает. Поэтому стабильность состояния природных экосистем является относительной, показателем которой может служить, например, периодически изменяющаяся численность популяций разных видов в экосистеме: численность одних видов увеличивается, других – уменьшается. Такое динамически равновесное состояние, или состояние подвижно—стабильного равновесия экосистем, называют гомеостазом (от греч. гомео – тот же; стазис – состояние).
Ключевой для понимания гомеостаза экосистем термин «подвижно-стабильное равновесие» означает, что устойчивое функционирование экосистем в изменяющихся условиях среды возможно именно вследствие того, что экосистема находится в квазиравновесном состоянии, принципиально отличающимся от понимания состояния равновесия в физике. Чтобы понять это различие, кратко рассмотрим составные части этого термина.
а) Стабильность означает, что природные экосистемы существуют в течение длительного времени и обладают определенной относительной стабильностью во времени и пространстве. Заметим, что особенностью искусственных (техногенных, созданных человеком) экосистем является то, что человек сам должен поддерживать равновесие в этих экосистемах, т.е. управлять процессами их функционирования, например, замена ила в региональных, муниципальных или производственных водоочистных сооружениях, в которых культивируются колонии бактерий, пожирающих, сорбирующих, разлагающих загрязняющие вещества в сточных водах.
б) Подвижность означает изменчивость свойств (например, численности популяций) и структуры экосистемы, т.е. совокупности видов. Последовательные изменения в состоянии равновесия в природных экосистемах отражаются в смене видов (например, в процессе сукцессии), сопровождающейся и изменениями в структуре и свойствах трофических цепей (сетей). Разнообразие видов формирует сукцессию, обеспечивая заполненность пространства жизнью и увеличивая степень замкнутости биогеохимического круговорота в экосистеме.
Следовательно, гомеостатичность – общее свойство всех экосистем, зависящее от эффективности комплекса адаптационных механизмов, действующих как на уровне отдельных видов, так и на уровне экосистемы в целом. Гомеостатичность зависит от возраста и видового разнообразия экосистем и поэтому сильно различается как у разных сообществ, так и в естественных и искусственных экосистемах.
studfiles.net