Тонкости микрошпильки
В зарубежных видеороликах по столярному делу мастера часто используют гвоздезабивные пистолеты для быстрого и аккуратного соединения деревянных элементов. Самым популярным инструментом для такого рода работ является шпилькозабивной пистолет.
Технические характеристики
В мире гвоздезабивных инструментов шпилькой называют такой тип гвоздя, у которого отсутствует шляпка. Наиболее часто встречаются шпильки с сечением 1.25, 0.8 и 0.64 мм. Последние еще именуются микрошпилькой. Микрошпильки загружаются в инструмент в виде кассеты (по 100 элементов в кассете). Процесс производства этого типа крепежа состоит из трех этапов. Сначала сотня тонких проволок нужного сечения склеиваются в полотно. Затем полотно рубится на пластины нужной длины. На последнем этапе одна из сторон шлифуется, вторая остается острой. Несмотря на отсутствие шляпки, микрошпильки имеют направление, и загружать в инструмент их надо ориентируясь на маркировку на кассете. Поскольку шпильки в кассете проклеены, при вхождении в материал, клей на ней разогревается и фиксируется внутри древесины.
| Шпиличник в работе | Зарядка кассеты | Шпилька 23 тип |
Существует разновидность микрошпильки — микрогвоздь. При сечении 0.64 мм он имеет шляпку 0.9 мм и лучше подходит для крепления тонких материалов до 3 мм и изделий из рыхлой древесины, например сосны. О его применении мы расскажем чуть ниже.Длина микрошпильки может варьироваться от 12 до 50 мм. При выборе нужной длины следуйте правилу: если нет ограничений по толщине материала, то длина шпильки должна быть равна двум толщинам закрепляемого элемента. Если материалы деталей сильно различаются по плотности, можно увеличить коэффициент до трех.
Шпилькозабивные пневматические пистолеты работают с давлением от 4 до 7.6 бар. Давление подбирается экспериментальным путем и зависит от плотности материала, с которым предстоит работать. Если пистолет уверенно утапливает шпильку на 4 атмосферах, то нет никакой необходимости увеличивать давление.
Существуют шпилькозабивные пистолеты с регулировкой заглубления или без нее. Регулировка осуществляется настройкой прижима носовой части (или прижимной скобы). Стандартно боек выходит из ствола и утапливает микрошпильку. Если пистолет имеет регулировку заглубления, то, отрегулировав ход носовой части, можно выровнять шпильку в плоскости с материалом.Шпилькозабивные пистолеты очень просты и не требовательны к обслуживанию. Если в вашей системе подачи воздуха не стоит масленка, следует наливать 1-2 капли не синтетического масла (веретенки) в штуцер инструмента перед началом рабочего дня. Это простое правило увеличит ресурс резиновых прокладок инструмента.
Некоторые советы при работе со шпилькозабивным пистолетом:
- Если пистолет не имеет силиконовой насадки, то во избежание вмятин от инструмента при отскоке, отпускайте курок после того, как отведете пистолет от материала.
 Так отдача от возврата поршня будет происходить не в материал.
Так отдача от возврата поршня будет происходить не в материал. - Пистолеты, не оснащенные регулировкой заглубления, всегда утапливают шпильку на 0,6-0,8 мм. Держите пистолет максимально перпендикулярно к поверхности, след от бойка будет меньше.
Крепление элементов декора микрошпилькой
Применение микрошпильки — для фиксация тонких декоративных элементов, наличников, торцевых планок, раскладок получает всё большее распространение. Если деталь имеет нагрузку на вырыв, ее предварительно приклеивают и затем фиксируют микрошпилькой. Так поступают с тяжелыми наличниками, если нет возможности опереть их на пол, или потолочными плинтусами из дерева.
Стоит заметить, что если наличник сосновый, лучше использовать микрогвоздь. При склейке деревянных заготовок требуется большое количество струбцин. Во многих случаях, первоначальная фиксация заготовок шпилькой позволяет не занимать струбцины. Поскольку сечение шпильки всего 0.64 мм, она легко пилится и шлифуется при последующей обработке заготовки.
| Крепёж дверных наличников | Крепление плинтусов | Раскладки и декоративные элементы |
Существуют области, в которых без микрошпильки приходится очень трудно. Примером может быть обшивка шпоном стены. Шпон приклеивается на двухкомпонентный клей и фиксируется микрогвоздем. Без этого приходилось бы тратить много времени в ожидании, когда клей схватится. При работе с тонкими материалами 3-5 мм также предпочтительно использовать микрогвозди. Примером такого применения может служить декоративная раскладка на дверях или резные элементы мебели.Выбираем инструмент правильно.
Микрошпилька — самый популярный тип крепежа для столярных работ. Она присутствует в линейке практически всех возможных производителей пневматических пистолетов. Различия по цвету (золотистые, бронзовые) не имеют никакого значения, поскольку в материале будет виден только срез, а он всегда стального цвета. Советуем обращать внимание только на качество, ведь качество инструмента — это качество вашей работы.Чтобы выбрать качественную микрошпильку для инструмента, нужно следить за двумя показателями. Во-первых, кассета со шпильками должна быть ровной и не иметь «волн» в плоскости. Иначе работа с таким крепежом будет часто приводить к засору в стволе инструмента. Во-вторых, микрошпилька должна быть острой — рубленой только с одной стороны, вторая должна быть отшлифована. Если оба конца рубленные — боек инструмента придется менять чаще.
Советуем обращать внимание только на качество, ведь качество инструмента — это качество вашей работы.Чтобы выбрать качественную микрошпильку для инструмента, нужно следить за двумя показателями. Во-первых, кассета со шпильками должна быть ровной и не иметь «волн» в плоскости. Иначе работа с таким крепежом будет часто приводить к засору в стволе инструмента. Во-вторых, микрошпилька должна быть острой — рубленой только с одной стороны, вторая должна быть отшлифована. Если оба конца рубленные — боек инструмента придется менять чаще.
Инструмент для работы микрошпилькой имеет несколько особо важных параметров:
- Размеры шпильки, с которой работает инструмент. Ограничение может быть не только по максимальной длине шпильки, но и по минимальной и даже по шагу изменения длины. 18 мм или 20 мм могут быть критичны для некоторых моделей.
- Наличие регулировки заглубления обеспечит максимально аккуратную работу, что важно при работе с уже окрашенными или лакированными элементами.

- Мягкая насадка на стволе инструмента позволит избежать вмятин от прижатия к материалу и уменьшит вероятность проскальзывания при работе под наклоном.
- Комбинированные модели, которые работают с микрогвоздем и микрошпилькой, более универсальны, но и след от них может быть чуть заметнее. Причина этого — устройство бойка.
- Механизм защиты от холостого выстрела спасет ваш материал от пустых лунок, если вы забудете загрузить шпильки в магазин инструмента.
Стандартным сечением для микрошпильки считается 0.64 мм. Некоторые производители указывают сечение 0.6 мм. Эта разница не критична и ваш инструмент сможет работать и такими шпильками. Иностранная маркировка микрошпильки — 23 Ga или gauge. Индекс на упаковках может содержать буквы MP, SP, Pins, Mini Pin nails. Для микрогвоздя сечение будет 0.64 и шляпка 0.64×0.9 мм. Индексные буквы на упаковке: MB, Mini Tip nails.Несмотря на то, что пневматика пока еще не столь распространена в России, как за рубежом, у нас представлены практически все мировые производители шпилькозабивных пистолетов.
Отделочные гвозди и шпильки 18Ga
Залог успешного использования любого профессионального инструмента — покупка качественных и надежных расходных материалов. От качества используемых расходных материалов зависит не только срок службы инструмента, но и безопасность персонала, использующего этот инструмент.
Компания GNGroup предлагает расходники и аксессуары для пороховых монтажных пистолетов, газовых гвоздезабивных пистолетов и пневматических гвозде и скобозабивных инструментов от ведущих производителей.
В нашем ассортименте:
• Барабанные гвозди по дереву для производства паллет и деревянного домостроения — оцинкованные и без покрытия;
• Дюбель-гвозди по бетону для пороховых и газовых монтажных пистолетов — гальванизированные и из нержавейки;
• Специализированный крепеж: гвозди с резьбой, дюбель-гвозди для монтажа пластиковых кабельканалов, усиленные гвозди для суперпрочного бетона и метала и т. д.;
д.;
• Газовые баллоны для газовых гвоздезабивных инструментов;
• Кровельные гвозди с усиленной гальванизацией для пневмопистолетов;
• Отделочные гвозди и штифты по дереву и каленые отделочные гвозди по бетону;
• Реечные гвозди по дереву: черные, оцинкованные, с усиленной гальванизацией — на бумажной или пластиковой кассетах;
• Скобы по дереву большого, среднего и малого сечения для каркасных и обивочных работ;
• Микрошпильки и микрогвозди для пневматических шпилькозабивных пистолетов с фрезеровкой тыльной стороны для увеличения ресурса пневмоинструмента.
• Саморезы в ленте для монтажа гипсокартона, ГВЛ, ДВП и других строительных материалов к дереву или металлу.
Весь товар в нашем каталоге сертифицирован и протестирован в реальных условиях на реальном оборудовании, он зарекомендовал себя на многих фабриках, производствах и строительных площадках по всей России.
Шпилька для пневмопистолета Тип-GA (Prebena)
Микрошпильки, шпильки и штифты для пневмопистолета нашли широкое
применение как в различных областях промышленного производства так и в
быту потому, что обеспечивают надежное крепление материалов, при этом
точки крепления либо не видно, либо она имеет аккуратный эстетичный вид.
Микрошпильки широко применяются при креплении различных деталей, где
требуется сделать место крепления невидимым.
Шпильки применяются при креплении более крупных деталей, где
микрошпилька уже не может обеспечить должной надежности крепления.
Штифт применяется при креплении вагонки, штапика, паркетной доски,
различных раскладок, где необходимо сочетать надежность крепления и
визуальную аккуратность места соединения.
Доставку инструмента по Украине осуществляем в течении 1-2 суток после заказа грузоперевозчиками «Ночной-Экспресс», «Новая почта», «Деливери», «Интайм», «Автолюкс», «САТ», «Гюнсел» и т. д.
Область применения:
крепление профиля, бордюров, обклада, применяется при изготовлении деревянных дверей.
- Высота шпильки — 16 — 30 мм
- Размер проволоки — 0,95х1,25 мм
- Шпилька без шляпки
Количество в упаковке:
Шпилька столярная — GA-16: — 10,000 тыс. шт в пачке;
Шпилька столярная — GA-19: — 10,000 тыс. шт в пачке;
Шпилька столярная — GA-25: — 5,000 тыс. шт в пачке;
Шпилька столярная — GA-30: — 5,000 тыс. шт в пачке.
_________________________________________________________________________________________________________
Цена: 140 грн. За пачку.
Есть в наличии.
_________________________________________________________________________________________________________
Производитель: Германия
_________________________________________________________________________________________________________
| Название: | ЧП «ДОНПРАЙМ» | |
| Контактное лицо: | Валерий Плахотник | |
| Адрес: | пр. Таманский, 18 б, 3-й этаж, офис 34,
Донецк,
83114,
Украина Таманский, 18 б, 3-й этаж, офис 34,
Донецк,
83114,
Украина | |
| Телефон: | +380 (62) 345-06-54,
факс +380 (62) 349-78-19, стационарный +380 (50) 347-10-51,
МТС +380 (63) 589-85-65, LIFE +380 (67) 153-91-36,
КИЕВСТАР | |
| Email: | donprime@mail.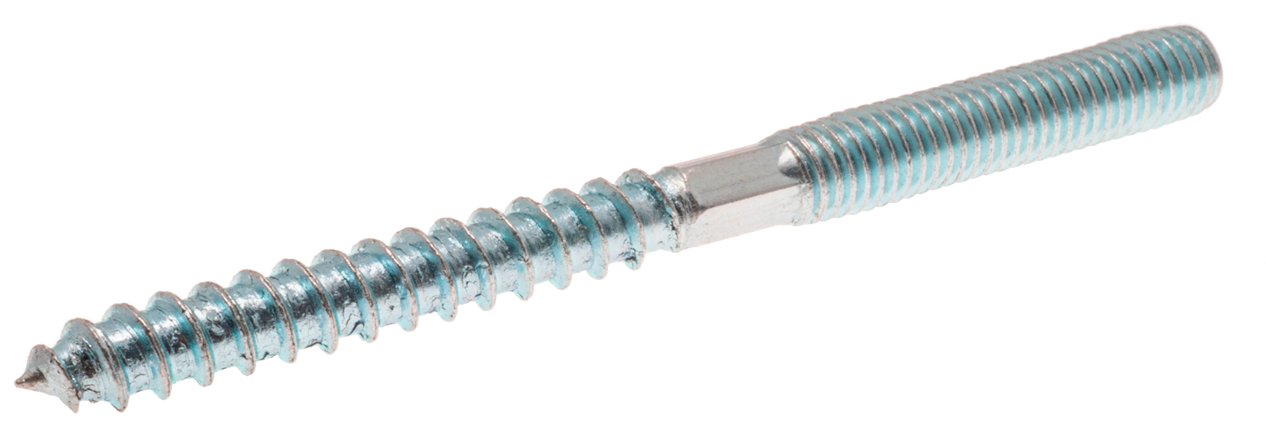 ru ru | |
| Skype: | donprime1 | |
| Сайт: | http://www.donprime.com.ua, http://www.donprime.promdex.com, http://donprime.skynell.biz, http://donprime.ua.hotlist.biz, http://donprime-vp.blogspot.com, http://donprime-pila.blogspot.com, http://donprime.uaprom.net, http://donprime.biz-kat.com.ua, http://donprime.promobud.ua, http://donprime.era.com.ua, donprime.wix.com/donprime-shlang, http://donprime.zakupka.com https://sites.google.com/site/rukavashlangi, http://donprime.stroika.biz.ua https://sites.google.com/site/lentochnyepily http://donprime.sells.com.ua http://donprime.key.ua http://blog.i.ua/user/2293293/ http://pnevmoskobupnevmogvozdi.blogspot.com http://pnevmoskobu.sells.com.ua https://sites.google.com/site/pnevmoskobupnevmogvozdi http://skobygvozdi.key.ua http://pnevmogvozdi.key.ua | |
| Время работы: | 09:00 — 17:00 |
|
Шпилька и образование дуплекса октамера ДНК d (m5C-G-m5C-G-T-G-m5C-G) в растворе.
 ЯМР-исследование.
ЯМР-исследование.Nucleic Acids Res. 1986 27 мая; 14 (10): 4187–4196.
Эта статья цитируется в других статьях в PMC.Abstract
Частично самокомплементарный октамер ДНК d (m5C-G-m5C-G-T-G-m5C-G) исследовали методом ЯМР-спектроскопии в растворе. Показано, что этот специфический фрагмент ДНК при подходящих условиях концентрации, соли и температуры предпочитает исключительно мономерную форму шпильки со стержнем из трех пар оснований типа Уотсона-Крика и петлей из двух остатков.При высокой однонитевой концентрации (8 мМ ДНК) и низкой температуре (т.е. ниже 295 К) шпилька находится в медленном равновесии со структурой B-димера. При высокой ионной силе (больше или равной 100 мМ Na +) и / или в присутствии метанола появляется третья разновидность, которая относится к Z-подобному димеру. В форме B, как и в димере Z, две центральные пары оснований образуют пары колебаний G.T с основаниями в качестве основных таутомеров.
Полный текст
Полный текст доступен в виде отсканированной копии оригинальной печатной версии. Получите копию для печати (файл PDF) полной статьи (675K) или щелкните изображение страницы ниже, чтобы просмотреть страницу за страницей. Ссылки на PubMed также доступны для Избранные ссылки .
Получите копию для печати (файл PDF) полной статьи (675K) или щелкните изображение страницы ниже, чтобы просмотреть страницу за страницей. Ссылки на PubMed также доступны для Избранные ссылки .
Избранные ссылки
Эти ссылки находятся в PubMed. Это может быть не полный список ссылок из этой статьи.
- Brown T, Kennard O, Kneale G, Rabinovich D. Структура спирали ДНК с высоким разрешением, содержащей несовпадающие пары оснований.Природа. 13 июня 1985 г .; 315 (6020): 604–606. [PubMed] [Google Scholar]
- Кеннард О. Структурные исследования фрагментов ДНК: пара оснований колебания G.T в ДНК A, B и Z; пара оснований G.A в B-ДНК. J Biomol Struct Dyn. 1985 Октябрь; 3 (2): 205–226. [PubMed] [Google Scholar]
- Патель Д. Д., Козловски С. А., Марки Л. А., Райс Дж. А., Брока С., Даллас Дж., Итакура К., Бреслауэр К. Дж.. Структура, динамика и энергетика дезоксигуанозина. образование пары оснований тимидинового колебания в самокомплементарном дуплексе d (CGTGAATTCGCG) в растворе.
 Биохимия. 2 февраля 1982 г.; 21 (3): 437–444. [PubMed] [Google Scholar]
Биохимия. 2 февраля 1982 г.; 21 (3): 437–444. [PubMed] [Google Scholar] - Tibanyenda N, De Bruin SH, Haasnoot CA, van der Marel GA, van Boom JH, Hilbers CW. Влияние несоответствий одной пары оснований на стабильность дуплекса d (T-A-T-T-A-A-T-A-T-C-A-A-G-T-T-G). d (C-A-A-C-T-T-G-A-T-A-T-T-A-A-T-A). Eur J Biochem. 1984 15 февраля; 139 (1): 19–27. [PubMed] [Google Scholar]
- Haasnoot CA, Hilbers CW. Эффективное подавление водного резонанса в 1D- и 2D-FT-1H-ЯМР-спектроскопии биополимеров в водном растворе.Биополимеры. 1983 Май; 22 (5): 1259–1266. [PubMed] [Google Scholar]
- Wemmer DE, Chou SH, Hare DR, Reid BR. Дуплекс-шпилька переходы в ДНК: ЯМР-исследования на CGCGTATACGCG. Nucleic Acids Res. 1985 24 мая; 13 (10): 3755–3772. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Марки Л.А., Блюменфельд К.С., Козловски С., Бреслауэр К.Дж. Зависимые от соли конформационные переходы в самокомплементарном дезоксидодекануклеотиде d (CGCAATTCGCG): свидетельство образования шпильки.
 Биополимеры. 1983 апр; 22 (4): 1247–1257.[PubMed] [Google Scholar]
Биополимеры. 1983 апр; 22 (4): 1247–1257.[PubMed] [Google Scholar] - Патель DJ, Hilbers CW. Протонный ядерно-магнитный резонанс исследования износа двухцепочечного d-ApTpGpCpApT в растворе h3O. Биохимия. 1975, 17 июня; 14 (12): 2651–2656. [PubMed] [Google Scholar]
- Haasnoot CA, de Bruin SH, Berendsen RG, Janssen HG, Binnendijk TJ, Hilbers CW, van der Marel GA, van Boom JH. Структура, кинетика и термодинамика фрагментов шпильки ДНК в растворе. J Biomol Struct Dyn. Октябрь 1983 г.; 1 (1): 115–129. [PubMed] [Google Scholar]
- Cornelis AG, Haasnoot JH, den Hartog JF, de Rooij M, van Boom JH, Cornelis A.Локальная дестабилизация двойной спирали ДНК парой колебаний T — T. Природа. 1979 20 сентября; 281 (5728): 235–236. [PubMed] [Google Scholar]
- Haasnoot CA, den Hartog JH, de Rooij JF, van Boom JH, Altona C. Петлевые структуры в синтетических олигодезоксинуклеотидах. Nucleic Acids Res. 1980, 11 января; 8 (1): 169–181. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Tinoco I, Jr, Uhlenbeck OC, Levine MD.
 Оценка вторичной структуры рибонуклеиновых кислот. Природа. 1971 г., 9 апреля; 230 (5293): 362–367. [PubMed] [Google Scholar]
Оценка вторичной структуры рибонуклеиновых кислот. Природа. 1971 г., 9 апреля; 230 (5293): 362–367. [PubMed] [Google Scholar] - Hilbers CW, Haasnoot CA, de Bruin SH, Joordens JJ, van der Marel GA, van Boom JH.Формирование шпильки в синтетических олигонуклеотидах. Биохимия. 1985 июль-август; 67 (7-8): 685–695. [PubMed] [Google Scholar]
Статьи из Nucleic Acids Research предоставлены здесь с любезного разрешения Oxford University Press
Произошла ошибка при настройке вашего пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.
 Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie. - Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
образование пары оснований GA в петле
Nucleic Acids Research, 1997, Vol. 25, No. 8
1546
Из остатков непарной петли в петле GTTA, T8 складывается на
своего 5′-соседа (рис. 7b), что не совсем верно для
7b), что не совсем верно для
T9. Последний направлен от петли и направлен в растворитель
(рис. 7в). Тем не менее, петлевую структуру GTTA, установленную здесь
, можно рассматривать как принадлежащую к категории I типа петли
складывания. Следовательно, результаты этого исследования согласуются с идеей
о существовании небольшого количества паттернов сворачивания для четырехчленных петель
.
Для тетрапетлей, содержащих пару оснований Уотсона – Крика (минилупы)
, было обнаружено, что положение первого неспаренного основания может
зависеть от его природы.Было высказано предположение, что если этот остаток
представляет собой пурин, он имеет тенденцию укладываться на своего 5′-соседа, тогда как остаток пиримидина
предпочтительно складывается в малую бороздку
(39). Это правило также справедливо для тетрапетлей, содержащих различные другие
типов пар оснований (24,40,41). Следовательно, можно было бы ожидать
, что второй остаток в петле GTTA будет
, расположенный в малой бороздке. Тот факт, что это не так,
Тот факт, что это не так,
можно легко понять, рассматривая геометрию пары оснований
, разрезанной на сдвиг, G-A.В этой конфигурации основание аденина имеет
, смещенное в сторону малой канавки (см. Рис. 7), что предотвращает складывание T8
в эту канавку. Следовательно, результаты, представленные здесь
, предполагают, что общее сворачивание петли, содержащей пару оснований G-A
, не определяется природой второго основания. Таким образом, подобный образец стэкинга
можно было бы ожидать для петель с пурином
в качестве второго остатка петли. Сопоставимые температуры плавления
для d (ATCCTA-GTTA-TAGGAT) и d (ATCCTA-
GAAA-TAGGAT), т.е.е. 55 и 53 ° C соответственно (23), а также
по результатам Sandusky et al. (8), подтверждают это.
Взаимодействие пары оснований G-A и замыкающего основания
пара стержня
Ряд конформационных исследований был проведен в
, в котором срезанная пара оснований G-A была обнаружена в четырехчленной петле
. Большинство этих исследований касалось петель
Большинство этих исследований касалось петель
, соответствующих консенсусной последовательности GNRA, мотив которой часто
обозначается как необычно стабильный.Однако фактическая стабильность этих
петель сильно различается, и может возникнуть вопрос относительно
, действительно ли все они содержат срезанную пару оснований G-A.
Недавно Sandusky et al. (8) наблюдали, что стабильность тетрапетлей
GNNA сильно зависит от полярности пары оснований C · G
, лежащих внизу, явление, также известное
для других петель шпильки (1). Было обнаружено, что температура плавления петли
, состоящей из C-GGCA-G, почти на 20 ° C выше на
, чем у последовательности G-GGCA-C, и был сделан вывод, что наличие
пары срезанных GA во втором случае
маловероятно.Авторы объяснили это наблюдение, предсказав
очень плохих взаимодействий стэкинга для гипотетической ситуации
срезанной пары оснований G-A, расположенной поверх пары оснований G · C,
, особенно между двумя последовательными пуринами (GG). Эти результаты
могут свидетельствовать о том, что правило, сформулированное Рейдом и
соавторами, то есть конфигурация пары оснований со сдвигом GA образуется только
, когда предыдущей парой оснований является C · G или T · A (19), также
относится к парам оснований GA в петлях.Однако представленные здесь результаты
показывают иное. Мы находим четко определенную петлевую структуру,
, включающую срезанную пару оснований G-A, поверх фрагмента A · T. Стекирующие взаимодействия
между этими двумя парами оснований определенно на
намного лучше, чем предсказанные для блока 5′-GG-3 ‘/ 3′-CA-5’
(8). В частности, стэкинг A6-G7, встречающийся для петли
A-GTTA-T, по-видимому, имеет тот же порядок, что и взаимодействие аденин-гуанин
в структуре, принятой
5′-CG-3 ‘/ Установка 3′-GA-5 ′.Сходные результаты, наблюдаемые здесь, были
, обнаруженными ранее для тетрапетли РНК GAGA, расположенной поверх
пары оснований A · U (7). К сожалению, нам не известны термодинамические или структурные данные
К сожалению, нам не известны термодинамические или структурные данные
для петель, состоящих из последовательности
T (U) -GNNA-A. Тем не менее, структура петли T-GCA-A
(9) указывает, что в такой ситуации возможны благоприятные взаимодействия стэкинга
.
В заключение, возникает картина, что стабильность петли
, содержащей срезанную пару оснований GA, относительно нечувствительна к полярности
базовой пары оснований A · T (U), в отличие от ситуации
, в которой это базовая пара C · G.Этот эффект, возможно, связан с направлением дипольного момента основания гуанина
, что более или менее противоположно таковым у тимина,
аденина и цитозина (42). Это может привести к более благоприятной ситуации
для стеков CG, AG и TG по сравнению со стеком GG
, и, следовательно, может привести к более благоприятному стэку
взаимодействий в 5′-CG-3 ‘/ 3’- GA-5 ‘, 5′-AG-3′ / 3’-TA-5 ‘и
5′-TG-3′ / 3′-AA-5 ‘единиц и менее благоприятные взаимодействия для
5′-GG -3 ′ / 3′-CA-5 ′.
БЛАГОДАРНОСТИ
Спектры ЯМРбыли записаны в Голландском национальном центре hf-ЯМР
(Неймеген, Нидерланды) при поддержке SON. Мы
благодарим J.Joordens за отличную техническую помощь.
ССЫЛКИ
1 Hilbers, C.W., Heus, H.A., Van Dongen, M.J.P. и Вейменга, С.С. (1994) В
Экштейн, Ф. и Лилли, Д.М.Дж. (ред.), Нуклеиновые кислоты и молекулярная биология.
Springer-Verlag, Берлин, Германия, Vol. 8. С. 56–104.
2 Хирао, И., Нисимура, Ю., Нарока, Т., Ватанабе, К., Арата, Ю. и Миура, К.-я.
(1989) Nucleic Acids Res., 17, 2223–2231.
3 Hirao, I., Nishimura, Y., Tagawa, Y.-i., Watanabe, K. и Миура, К.-я. (1992)
Nucleic Acids Res., 20, 3891–3896.
4 Хирао, И., Нисимура, Ю., Тагава, И.-и., Ватанабэ, К. и Миура, К.-я. (1994)
Nucleic Acids Res., 22, 576–582.
5 Heus, H.A. и Парди, А. (1991) Science, 253, 191–193.
6 Санта-Люсия, Дж., Младший, Кежек, Р. и Тернер, Д. (1992) Science, 256,
217–219.
7 Орита, М., Нисикава, Ф., Симаная, Т., Тайра, К., Эндо, Ю. и Нисикава, С.
(1993) Nucleic Acids Res., 21, 5670–5678.
8 Сандаски П., Вутен Э. У., Курочкин А. В., Кавано Т., Мандеки В.
и Zuiderweg, E. (1995) Nucleic Acids Res., 23, 4717–4725.
9 Чжу, Л., Чжоу, С.-Х., Сюй, Дж. и Рид Б. (1995) Nature Struct. Биол., 2,
1012–1017.
10 Woese, C.Р., Винкер С. и Гутелл Р. (1990) Proc. Natl. Акад. Sci. США,
87, 8467–8471.
11 Катасти, П., Гупта, Дж., Гарсия, А.Э., Ратлифф, Р., Хонг, Л., Яу, П., Мойзис, Р.К.
и Брэдбери, Э. (1994) Biochemistry, 33, 3819–3830.
12 Феррер, Н., Азорин, Ф., Вилласанте, А., Гутьеррес, К. и Abad, J.P. (1995)
J. Mol. Биол., 245, 8–21.
13 Ли, Ю., Зон, Г. и Уилсон, У. (1991) Proc. Natl. Акад. Sci. США, 88,
26–30.
14 Li, Y., Зон, Г. и Уилсон, У. (1991) Biochemistry, 30, 7566–7572.
15 Chou, S.-H., Cheng, J.W. и Рид Б. (1992) J. Mol. Биол., 228, 138–155.
16 Чжоу, С.-Х., Ченг, Дж. У., Федоров, О. и Рид Б. (1994) J. Mol. Биол.,
241, 467–479.
17 Санта-Люсия, Дж., Младший и Тернер, Д.Х. (1993) Biochemistry, 32, 12612–12623.
18 Готере Д., Конингс Д. и Гутелл Р. (1994) J. Mol. Биол., 242, 1–8.
19 Cheng, Y.K., Chou, S. -H. и Рид Б. (1992) Дж.Мол. Биол., 228,
1037–1041.
20 Уолтер, А.Э., Ву, М. и Тернер, Д. (1994) Biochemistry, 33,
11349–11354.
21 Антао, В.П. и Tinoco, I., Jr (1992) Nucleic Acids Res., 20, 819-824.
22 Джакер, Ф.М., Хеус, Х.А., Пинг, Ф.Ю., Мурс, Э. и Парди, А. (1996)
J. Mol. Биол., 264, 968–980.
Петли шпильки, состоящие из единичных остатков аденина, закрытых срезанными парами A · A и G · G, образованными триплетами ДНК AAA и GAG: структура раствора d (GTACAAAGTAC) Шпилька
Abstract
Унудакамеры ДНК GTAC AAA GTAC (11-мерный AAA) и GTAC- GAG GTAC (11-мерный GAG) были исследованы в растворе методом ЯМР-спектроскопии высокого разрешения.Оба дуплекса образуют стабильные шпильки, содержащие одиночные дезоксиаденозиновые петли, и стержни, содержащие пять пар оснований, которые замыкаются на конце петли срезанными парами A · A и G · C соответственно. Таким образом, эти молекулы содержат новые мотивы поворота петли AAA и GAG. Все протоны, включая хиральные протоны H5 ‘/ H5 ″ петлевых остатков, были отнесены с использованием NOESY, DQF-COSY и гетероядерных экспериментов. Торсионные углы основной цепи были ограничены с использованием экспериментальных данных из кросс-пиков NOE, констант связи с тремя связями 1 H- 1 H с константами связи и с четырьмя связями 1 H- 31 P с константами связи и с четырьмя связями 1 H — 31 Константы связи P.11-меры AAA и GAG образуют в растворе аналогичные структуры. Подробная структура AAA 11-мер была определена путем комбинированного использования ЯМР, геометрии расстояния и методов минимизации энергии. Эта структура демонстрирует хорошее наложение петлевой аденозиновой основы на замыкающую сдвигающую пару 5A · 7A, с наложением основания 6A на основание 5A и наложение дезоксирибозы 6A с основанием 7A. Все сахара в 11-мерной шпильке AAA принимают типичную конформацию ДНК C2′- endo , и между остатками 6A и 7A происходит резкий поворот основной цепи.Этот виток петли вызван, главным образом, изменением углов кручения фосфата основной цепи с ζ ( г — ) α ( г — ) на ζ ( г + ) α ( г + ) на повороте. Угол кручения γ остатка 7А в замыкающей сдвигающейся паре также изменяется с гош + до транс . В Pu 1 NPu 2 витков петли типов GCA, AAA и GAG химический сдвиг протона h5 ‘дезоксирибозы петли зависит от природы Pu 2 ; это отражает наложение петлевых сахаров на основу из Pu 2 и различные эффекты кольцевого тока A или G в этом положении.
Ключевые слова
Петля AAA
Петля ГАГ
пара Pu · Pu
шпилька
Структура ЯМР
Рекомендуемые статьиЦитирующие статьи (0)
Полный текстCopyright © 1996 Academic Press. Все права защищены.
Рекомендуемые статьи
Цитирующие статьи
Асимметричная структура пяти- и шестичленных петель шпильки ДНК
Lilley DMJ (1982) в Neidle S [Ed] Topics in nucleic acid structure, vol.2 (стр. 173–198) Mcmillan, London
Google Scholar
Kuhn H & Waser J (1981) Angew. Chem., Int. Эд. Англ. 20: 500–520
Google Scholar
van de Ven FJM & Hilbers CW (1988) Eur. J. Biochem. 178: 1–38
Google Scholar
Gralla J & Crothers DM (1973) J. Mol. Биол.73: 497–511
Google Scholar
Haasnoot CAG, de Bruin SH, Berendsen RG, Janssen HGJM, Binnendijk TJJ, Hilbers CW, van der Marel GA и van Boom JH (1983) J. Biomol. Struct. Дин. I: 115–129
Google Scholar
Uhlenbeck OC, Borer PN, Dengler B. & Tinoco I Jr (1973) J. Mol. Биол. 73: 483–496
Google Scholar
Baumann U, Frank R & Blöcker H (1986) Eur. J. Biochem. 161: 409–413
Google Scholar
Xodo LE, Manzini G, Quadrifoglio F, van der Marel G & van Boom JH (1991) Nucl. Acids Res. 19: 1505–1511
Google Scholar
Kinjo M, Hasegawa T., Nagano K, Ishikura H & Ishigami M (1986) J. Mol. Evol. 23: 320–327
Google Scholar
Дрю Х.Р. (1984) J. Mol. Биол. 176: 535–557
Google Scholar
Vournakis JN, Celantano J, Finn M, Lockard RE, Mitra T, Pavlakis G, Troutt A, van den Berg M & Wurst RM (1981) в Chirikjian JG & Papas TS [Eds] Амплификация и анализ генов , т. 2 (стр. 267–298) Elsevier North Holland, Нью-Йорк
Google Scholar
Göringer H-U, Szymkowiak C & Wagner R (1984) Eur.J. Biochem. 144: 25–34
Google Scholar
Wells RD et al. (1977) Крит. Rev. Biochem. 4: 305–340
Google Scholar
Bacolla A & Wu FY-H (1991) Nucl. Acids Res. 19: 1639–1647
Google Scholar
Szafranski P & Godson GN (1990) Gene 88: 141–147
Google Scholar
Laskowski Sr M (1980) Methods Enzymol. 65: 263–275
Google Scholar
Green MR & Roeder RG (1980) Cell 22: 231–242
Google Scholar
Baumann U, Fischer W & Spinzl M (1985) Eur. J. Biochem. 152: 645–649
Google Scholar
Sundquist WI и Klug A (1989) Nature 342: 825–829
Google Scholar
Rich A & RajBhandary UL (1976) Ann. Rev. Biochem. 45: 805–860
Google Scholar
Moras D, Comarmond MB, Fischer J, Weiss R, Thierry JC, Ebel JP & Giege R (1980) Nature 288: 669–674
Google Scholar
Wrede P, Woo NH & Rich A (1979) Proc. Natl Acad. Sci. 76: 3289–3293
Google Scholar
Baumann U, Lehmann U, Schwellnus K, van Boom JH и Kuhn H (1987) Eur. J. Biochem. 170: 267–272
Google Scholar
Haasnoot CAG, Hilbers CW, van der Marel GA, van Boom JH, Singh UC, Pattabiranam N & Kollman PA (1986) J. Biomol. Struct. Дин. 3: 843–857
Google Scholar
Blommers, MJJ, van de Ven FJM, van der Marel GA, van Boom JH & Hilbers CW (1991) Eur.J. Biochem. 201: 33–51
Google Scholar
Senior MM, Jones RA & Breslauer KJ (1988) Proc. Natl Acad. Sci. 85: 6242–6246
Google Scholar
Поток индуцирует переход от петли к β-шпильке на β-переключателе гликопротеина тромбоцитов Ibα
Abstract
Взаимодействие гликопротеина Ibα (GPIbα) с фактором фон Виллебранда (VWF) инициирует адгезию тромбоцитов к поврежденной стенке сосуда, чтобы остановить кровотечение.Основной контакт между GPIbα и VWF включает область β-переключателя, которая представляет собой петлю в нелигандованном GPIbα, но переключается на β-шпильку в сложной структуре. Парадоксально, но кровоток скорее усиливает, чем препятствует связыванию GPIbα-VWF. Мутации с усилением функции (например, M239V) в β-переключателе снижают потребность в потоке для связывания VWF, тогда как мутации с потерей функции (например, A238V) увеличивают потребность в потоке. Эти явления нельзя объяснить кристаллическими структурами или расчетами энергии. Здесь мы демонстрируем, что β-шпилька нестабильна без контакта с VWF, поскольку она переключается на петлю при моделировании свободной молекулярной динамики.Моделирование с использованием нового алгоритма молекулярной динамики потока показывает, что конформация петли нестабильна в присутствии потока, поскольку она переключается на β-шпильку даже без контакта с VWF. По сравнению с диким типом мутанту M239V легче, но труднее мутанту A238V переключиться на β-шпильку при наличии потока. Эти результаты проясняют структурную основу двух мутантов и предполагают регуляторный механизм, с помощью которого поток активирует GPIbα посредством индукции конформационного перехода петля-β-шпилька на β-переключателе, тем самым способствуя связыванию VWF.
Связывание гликопротеина Ibα (GPIbα) с фактором фон Виллебранда (VWF) инициирует многоступенчатую адгезию тромбоцитов и сигнальный каскад гемостатического процесса (1, 2). Дисфункция взаимодействия GPIbα-VWF может вызывать нарушения свертываемости крови, такие как болезнь фон Виллебранда (VWD) (3). Быстрая кинетическая скорость связывания GPIbα-VWF требуется для того, чтобы текущие тромбоциты прикрепились к поврежденной стенке сосуда. Однако, как это ни парадоксально, связывание GPIbα-VWF усиливается кровотоком, несмотря на тот факт, что увеличение потока сокращает время контакта для молекулярного взаимодействия (4).Для связывания тромбоцитов с VWF требуется минимальный поток (4, 5). Кинетические скорости связывания GPIbα-VWF и их механическое регулирование потоком могут быть изменены структурными изменениями. Мутации могут потребовать более высоких или более низких потоков для связывания тромбоцитов с VWF по сравнению с GPIbα дикого типа (WT) (6). Первый называется мутантом с усилением функции (GOF), а второй — мутантом с потерей функции (LOF). Мутации GOF G233V и M239V в природе встречаются у некоторых пациентов с VWD тромбоцитарного типа (5, 7, 8).
GPIbα состоит из гликозилированного N-концевого домена (GPIbαN), длинной муциновой ножки, трансмембранного домена и цитоплазматического хвоста (9, 10). Сайт связывания VWF находится на GPIbαN, который содержит восемь тандемных повторов, богатых лейцином (рис. 1 A ) (11). Мономерная субъединица VWF состоит из множества копий доменов типа A, B, C и D (12). Сайт связывания GPIbα находится в домене A1 (рис. 1 B , голубой) (13). Участки GPIbαN на каждом конце богатых лейцином повторов связываются, соответственно, с верхней и нижней частью домена A1 (рис.1 В ) (14, 15).
Сравнение нелигандированных (фиг.1 A ) и лигандированных (фиг.1 B ) структур GPIbαN (14, 15) показывает, что наибольшее конформационное изменение происходит в области С-концевой фланкирующей последовательности (остатки 229–240), который переключается с бесструктурной петли (рис. 1 C ; зеленый на рис. 1 A ) на β-шпильку (рис. 1 D ; красный на рис. 1 B ) , отсюда и название β-переключатель. Β-шпилька совпадает с центральным β-листом A1, образуя главную границу раздела комплекса (рис.1 В ). Почти каждый невалиновый остаток в β-переключателе был заменен валином в исследованиях мутагенеза (16). В дополнение к встречающимся в природе мутациям PT-VWD G233V и M239V, D235V и K237V также обладают фенотипом GOF (светло-голубой на рис. 1 C и D ). Напротив, K231V, Q232V и A238V (фиолетовый на рис. 1 C и D ) демонстрируют фенотип LOF.
Сравнение комплексов GPIbαN: A1 M239V (14) и WT (15) не дало немедленного понимания фенотипа GOF M239V.Расчеты энергии смоделированных мутантов также не основывались на равновесных структурах. Поскольку взаимодействие GPIbα-VWF с усилением потока является неравновесным процессом, мы предположили, что поток индуцирует динамическое конформационное изменение β-переключателя, которое способствует связыванию GPIbα-VWF. Мы разработали новый алгоритм молекулярной динамики потока всех атомов (FMD) для моделирования того, как поток влияет на WT и мутантные структуры. В отличие от алгоритмов, основанных на давлении (17) или SLLOD (18), наш алгоритм FMD генерирует поток, прикладывая силы непосредственно к молекулам воды, что легче контролировать.Мы исследовали, насколько стабильными были конформации петли и β-шпильки, насколько легко они переходили от одной к другой, может ли поток вызывать конформационные переходы и могут ли и как мутации GOF и LOF изменять регуляцию потока конформационных изменений. Результаты подтверждают нашу гипотезу и дают представление о регуляции потока связывания GPIbα-VWF. Эта работа демонстрирует с помощью вычислений, что поток жидкости может вызвать переход от неструктурированной к структурированной конформации в белке.
Результаты
Важность β-переключателя на связывание VWF подтверждается наблюдениями, что он образует основную контактную поверхность с A1, а мутации в этой области изменяют связывание с VWF. Чтобы выяснить структурные основы фенотипов этих мутантов, мы сравнили структуры комплексов GPIbαN: A1 с β-переключателем WT, M239V и A238V [вспомогательная информация (SI) Fig. S1 A – C ]. Соответствующие структуры мутантов GOF и LOF моделировались аналогичным образом на основе структуры WT.Мутации не изменяют взаимодействия β-листов с A1, которые не зависят от боковых цепей (Fig. S1 A – C ). Мы использовали бесплатное моделирование МД для исследования устойчивости и расчета энергий взаимодействия этих структур. Конформационных изменений не наблюдалось. Удивительно, независимо от их фенотипа, два мутанта демонстрируют сходные энергии взаимодействия между A1 и всем GPIbαN (рис. S1 D ) или одним β-переключателем (рис. S1 E ), которые немного ниже, чем у мутанта. WT.Таким образом, структурный анализ и расчеты энергии не дали немедленного понимания различных фенотипов этих мутантов GOF и LOF.
Кристаллографическое наблюдение β-переключателя в двух различных конформациях (рис. 1 A и B ) предполагает, что эта область структурно бистабильна. Когда GPIbαN был связан с A1, конформация β-шпильки была стабильной во время моделирования свободной динамики в течение 20 нс. Однако, когда β-шпилька была изолирована, она плавилась в петлю или показывала тенденцию к плавлению во всех 8 моделированиях продолжительностью 10 нс (три цикла) или 20 нс (пять циклов) (рис.S2). Это проиллюстрировано на рис. 2 A , на котором показаны снимки структуры в указанные моменты времени в репрезентативном моделировании (см. Также фильм S1). Конформационный переход от β-шпильки к петле обозначается постепенным разрывом шести водородных (H) связей, которые соединяют две ветви β-шпильки (см. Их нумерацию на рис. 1 D ). Чтобы количественно оценить этот процесс, мы построили график зависимости расстояния между каждой из шести донорно-акцепторных пар водородной связи от времени на рис.S2 для каждой из восьми симуляций и определите границу 3,5 Å, за которой водородная связь считается разорванной. Несмотря на вариации траекторий из разных моделей в результате тепловых флуктуаций, общая тенденция очевидна. Нарушение водородных связей началось на дальнем конце β-шпильки (т.е. водородной связи № 6), которая была разорвана во время уравновешивания во всех симуляциях до того, как было начато свободное моделирование МД (рис. S2). Плавление распространялось внутрь к ближнему концу β-шпильки (т.е.1), которая была сломана только в 1 из 8 заездов. Эта уменьшенная частота разрыва Н-связи с увеличением расстояния от дальнего конца β-шпильки четко показана на рис. 2 B . Это также можно увидеть, используя альтернативный показатель — частоту выживания — которая со временем снижается, как показано на рис. 2 C для средней кривой всех шести Н-связей по всем моделям (сплошная линия) или по пяти 20- нс пробегает (пунктир). Эти результаты демонстрируют, что без связывания с A1 β-шпилька нестабильна и спонтанно переходит в бесструктурную петлю.
Рисунок 1.Структуры GPIbα. Структуры нелигандированного GPIbαN ( A ) (код PDB 1QYY) (23) и GPIbαN в комплексе с VWF-A1 (голубой) ( B ) (код PDB 1SQ0) (15). Показаны N-концевая фланкирующая последовательность (сиреневый), богатые лейцином повторы (синий), С-концевая фланкирующая последовательность (охра) и остатки цистеина (шарики и палочки) GPIbαN. Конформация петли β-переключателя (остатки 227–243) в A показана зеленым, а его конформация β-шпилька в B показана красным.Конформации петли ( C ) и β-шпильки ( D ) увеличены соответственно, чтобы показать боковые цепи остатков 229–240 с цветами в соответствии с их фенотипами при мутации валина (розовый для LOF, серый для GOF. и оранжевый для остатков, которые уже являются валином в последовательности WT или остатков, фенотип которых неизвестен при мутации в валин). Боковые цепи остатка Thr-240 показаны зеленым, потому что разные экспериментальные анализы показали разные фенотипы. Конформация β-шпильки стабилизируется шестью Н-связями (отмечены желтым цветом в D ), пронумерованными 1–6 от ближнего конца β-шпильки наружу ( D ).
Рис. 2.Переход β-шпильки в петлю β-переключателя в отсутствие силы. ( A ) Моментальные снимки смоделированных изолированных структур β-переключателей в репрезентативном свободном моделировании MD, сделанные в последовательные моменты времени (указано), когда следующая ранее существовавшая водородная связь была разорвана. Моделирование началось с конформации β-шпильки (1SQ0) и закончилось через 10 нс после того, как все Н-связи (отмечены желтым) были разорваны, и β-шпилька превратилась в петлю. ( B ) Частота разрыва (количество симуляций, во время которых наблюдался разрыв, деленное на восемь, общее количество симуляций) для каждой Н-связи.( C ) Частота выживания (сумма текущих долей времени, в течение которых расстояния между всеми парами донорно-акцепторных водородных связей были ниже порогового значения 3,5 Å, деленное на исходное число шесть), усредненная по всем восьми запускам (сплошная кривая) или только пять прогонов по 20 нс (пунктирная кривая) в зависимости от времени. ( D ) Тест модели подгонки, индуцированной связыванием. Снимки трех свободных МД-моделирования комплекса GPIbαN: A1 (показаны только β-переключатель и β-цепь A1, с которой он контактирует) после того, как β-шпилька была разорвана силой, при сохранении ее взаимодействия с A1 полностью (левый столбец ) или частично (центральный столбец) нетронутыми, или заменой β-нитей петлей (взято из кода PDB 1QYY) (правый столбец).
Поскольку конформация петли кажется неблагоприятной для стыковки с A1, мы задали вопрос, является ли образование β-шпильки результатом связывания с A1 или β-шпилька преформирована до связывания A1. Первая возможность представляет собой гипотезу «соответствия, вызванного связыванием», которую мы проверили с помощью трех моделей ( SI Methods ). В первом моделировании две ветви β-шпильки в сокристаллической структуре GPIbαN: A1 сначала были разделены силой, сохраняя при этом свое взаимодействие с центральным β-листом A1 неповрежденным; затем система была подвергнута свободной динамике, чтобы увидеть, будет ли реформироваться β-шпилька.Это действительно наблюдалось всего за 0,4 нс моделирования (рис. 2 D , правый столбец и Movie S2). Результат предполагает, что связывание одной ветви петли с A1 будет индуцировать переход петля-шпилька β-переключателя. Это также является дополнительным подтверждением нашей гипотезы о том, что β-шпилька является стабильной конформацией, когда она связана с A1. Вторая симуляция аналогична первой, за исключением того, что две из четырех Н-связей, соединяющих β-переключатель с A1, также были разорваны. Три Н-связи между двумя ветвями β-переключателя преобразовались за 3 нс, но в оставшейся части этого моделирования свободной динамики за 25 нс больше не наблюдалось образования Н-связи (рис.2 D , средний столбец и фильм S3). Интересно, что две разорванные Н-связи между β-переключателем и A1 не реформируются стабильно, а две оставшиеся Н-связи стали слабее, поскольку расстояния между их донорами и акцепторами колебались выше и ниже линии отсечки, которая определяет Н-связь. больше, чем в первом моделировании (рис. S3 D и E ). Эти результаты указывают на наличие энергетического барьера, препятствующего связыванию неупорядоченного β-переключателя с A1, и предполагают, что, когда β-переключатель только частично связан с A1, движущая сила, которая индуцировала реформацию β-шпильки в первом моделировании, значительно ослабляется.В третьем моделировании структура β-шпильки в сокристалле GPIbαN: A1 была заменена петлевой структурой изолированного кристалла GPIbαN, который вообще не был связан с A1. Обратите внимание, что GPIbαN оставался связанным с A1 через другие контакты (например, через область β-пальца). Взаимодействия основной цепи между β-переключателем и A1 не наблюдалось, и не наблюдалось образования Н-связи между двумя ножками в β-шпильке при уравновешивании 10 нс и в другой свободной динамике 40 нс (Рис. 2 D , Рис. правый столбец и Movie S4).Это указывает на то, что, хотя мы не можем исключить существование пути «индуцированной подгонки», его движущая сила не обнаруживается в то время, когда выполнялась наша симуляция. На рис. S3 показаны графики изменения расстояний донорно-акцепторных пар водородной связи между двумя ножками β-переключателя (строки A-C) и между β-переключателем и A1 (строки с D по F) для трех моделей. Эти результаты не подтверждают гипотезу соответствия, вызванного связыванием.
Как можно было преформировать β-шпильку до связывания А1? Наблюдение, что для связывания тромбоцитов с VWF требуется минимальный поток, побудило нас предположить, что поток индуцирует переход от петли к β-шпильке, что способствует связыванию VWF.Чтобы проверить эту гипотезу, мы разработали новый алгоритм FMD и использовали его для проверки стабильности конформации петли в присутствии потока. Алгоритм FMD кратко описан в материалах и методах , а строгие проверки его достоверности представлены в другом месте (19). Примечательно, что при потоке изолированная петля быстро переходила к конформации β-шпильки во всех шести 6-нс независимых симуляциях для WT (Рис. S4 A – F ). На рис. 3A показаны снимки структуры WT в указанные моменты времени в репрезентативном моделировании (см. Также фильм S5).Интересно, что конформационный переход от петли к β-шпильке характеризовался теми же событиями, но следовал обратному временному ходу к переходу от β-шпильки к петле. Это обозначается постепенным образованием шести Н-связей, которые удерживают две ноги все ближе и ближе. Подобно предыдущему анализу перехода β-шпилька-петля, мы строим график зависимости расстояния между каждой из шести пар донорно-акцепторных водородных связей от времени и используем ту же отсечку 3,5 Å для определения водородной связи ( Инжир.S4). Образование водородных связей началось на ближнем конце β-шпильки (т.е. водородной связи № 1), которая была образована во всех моделях (рис. S4). Процесс распространялся наружу к дальнему концу β-шпильки (т.е. Н-связи № 6), которая не образовывалась в конце каких-либо циклов продолжительностью 6 нс, но явно демонстрировала тенденцию к образованию. Это можно увидеть на фиг. 3 D , где возрастающая частота образования Н-связи с течением времени нанесена на график для каждой из шести Н-связей (усредненные по шести прогонам). В определенный момент (например,g., 6 нс) частота образования Н-связи снижалась с увеличением расстояния от ближнего конца β-шпильки (рис. 3 G ). Это также можно показать с помощью альтернативного показателя — времени, необходимого для образования водородной связи, — которое увеличивается с увеличением расстояния от ближнего конца β-шпильки (рис. 3 H ). Эти результаты подтверждают нашу гипотезу и демонстрируют, что поток может индуцировать конформационный переход от петли к β-шпильке на β-переключателе даже без его связывания с A1.
Инжир.3.Индуцированный потоком переход изолированного β-переключателя из петли в β-шпильку. Снимки смоделированных структур β-переключателей WT ( A ), M239V ( B ) и A238V ( C ) в репрезентативных симуляциях FMD, сделанные в последовательные моменты времени (указано), когда образовывалась следующая новая водородная связь. Моделирование началось с конформаций петли (1QYY или смоделировано после 1QYY путем мутации остатка 239 или 238 на валин) и закончилось через 6 нс после образования первых пяти Н-связей, и петля почти превратилась в полную β-шпильку.Частоты образования (текущая часть времени, в течение которого расстояние между каждой парой донор / акцептор водородной связи было ниже порогового значения 3,5 Å) для указанных водородных связей усредняются по шести моделям и наносятся на график в зависимости от времени для WT ( D ) , M239V ( E ) и A238V ( F ). Некоторые кривые нечеткие, потому что они слишком близки к оси x . Конечные результаты при 6 нс показаны в G в виде гистограммы (среднее значение ± стандартная ошибка среднего для шести симуляций) для WT (светлые столбцы), M239V (заштрихованные столбцы) и A238V (сплошные столбцы).( H ) Время, необходимое для образования и сохранения каждой водородной связи в течение> 50 пс в первый раз (среднее значение ± стандартная ошибка среднего для шести симуляций) для WT (пустые столбцы), M239V (заштрихованные столбцы) и A238V (сплошные столбцы) .
Наше моделирование предполагает, что для изолированного β-переключателя петля стабильна, но β-шпилька нестабильна в отсутствие потока, тогда как петля нестабильна, а β-шпилька стабильна в присутствии потока. Таким образом, поток действует как механический регулятор, который переключает структуру GPIbα между двумя конформациями, а β-переключатель служит механическим датчиком.Это согласуется с экспериментально наблюдаемым усилением потока связывания GPIbα-VWF, потому что предварительно сформированная β-шпилька должна быть более благоприятной для стыковки с VWF-A1. Поскольку мутанты GOF / LOF в области β-переключателя понижают / повышают требования к потоку для связывания GPIbα-VWF, прогнозируется, что им потребуется меньше / больше времени, чем WT, чтобы вызвать переключение с петли на β-шпильку при идентичных условиях потока. Мы проверили это предсказание с помощью моделирования ящура с мутантом GOF M239V и мутантом LOF A238V. M239V был выбран потому, что этот встречающийся в природе мутант PT-VWD расположен в середине насаждений в конформации β-шпильки (рис.1 С ). Был выбран A238V, потому что он находится рядом с M239V, что упрощает сравнение. На рис. 3 B и C показаны, соответственно, снимки структур M239V и A238V в указанные моменты времени при репрезентативном моделировании (см. Также Movie S6 и Movie S7). Шесть снимков в каждой строке рис. 3 A – C выбраны таким образом, что первый соответствует началу моделирования FMD, а оставшиеся от двух до шести снимков соответствуют, соответственно, моментам, когда с первого по пятый H- облигации были сформированы.Из значений времени в каждом столбце на рис. 3 A – C мы можем видеть, что по сравнению с WT, M239V потребовалось меньше времени, тогда как A238V потребовалось больше времени для образования той же Н-связи в большинстве случаев. Опять же, мы количественно определяем переход от петли к β-шпильке расстоянием между каждой из шести пар донорно-акцепторных водородных связей, которое отображается в зависимости от времени в строках с A по F на рис. S5 и рис. S6 для каждой из них. из шести симуляций для M239V и A238V, соответственно. Соответствующие временные зависимости частоты образования для каждой Н-связи (усредненные по шести моделям) для M239V и A238V показаны на рис.3 E и F . Очевидно, что по сравнению с WT кривые M239V смещены вверх и влево в сторону более высокой частоты и более короткого времени, тогда как кривые A238V смещены вниз и вправо в сторону более низкой частоты и более длительного времени. Мы сравниваем соответствующие частоты образования для M239V и A238V при 6 нс с частотой образования WT для каждой водородной связи (рис. 3 G ) и соответствующее время, необходимое для образования водородной связи для M239V и A238V с таковым для WT. для каждой Н-связи (рис.3 H ). Очевидно, что частота образования водородной связи выше для M239V, но ниже для A238V, тогда как время, необходимое для образования водородной связи, меньше для M239V, но больше для A238V. Рис. 3 и Рис. S4 – S6 ясно показывают, что по сравнению с WT, M239V имеет пониженную потребность в потоке для переключения петля-шпилька, тогда как M239V увеличена. Эти результаты подтверждают нашу гипотезу и дополнительно демонстрируют, что поток может индуцировать конформационный переход от петли к β-шпильке на β-переключателе даже без связывания с A1.
Продолжающийся FMD применял поток в несколько десятков метров в секунду, чтобы значительно ускорить переход от петли к β-шпильке, чтобы его можно было наблюдать в течение нескольких наносекунд времени моделирования (Рис. S7 C и D ), что на 2 порядка больше скорости кровотока артериальной крови (20). Скорость потока относительно тромбоцита над его мембраной (испытываемая β-переключателем) еще меньше. Чтобы изучить, как скорость потока влияет на конформационный переход, мы уменьшили силы сопротивления наполовину (тем самым уменьшив скорость потока) в нескольких моделях.Образование Н-связи все еще наблюдалось, хотя оно происходило с меньшей скоростью (строка G на фиг. S4 – S6).
Может ли сконцентрированная сила вызвать аналогичные конформационные изменения в β-переключателе? Если да, что более эффективно, поток или сосредоточенная сила? Чтобы ответить на эти вопросы, мы использовали моделирование управляемой молекулярной динамики (SMD), чтобы исследовать эффект оттягивания силы 200 пН на дальнем конце петли (C α атомов Val-234 и Asp-235). Это тянущее усилие было выбрано для того, чтобы моделирование SMD было сопоставимо с моделированием FMD.В FMD сила 0,7 пН была приложена к каждому из ~ 300 атомов в слое мысли 2 Å, что в сумме составляет ~ 210 пН от общей приложенной силы. Результаты моделирования SMD показаны на рис. S4 H и I и S5 H . Хотя образование водородных связей наблюдалось в этих SMD-симуляциях, они происходили с гораздо меньшей частотой и длились намного меньше времени по сравнению с тем, что наблюдалось в FMD-симуляциях. Различная эффективность потока и сконцентрированной силы для индуцирования перехода петля-β-шпилька выявляется путем изучения сил взаимодействия (скользящих среднего во времени) между водой и белком, рассчитанных на основе FMD (рис.4 A ) и SMD (рис. 4 B ). Общая сила взаимодействия разбивается на две части: одна действует на боковые цепи, а другая — на белковый каркас. В FMD, поскольку боковые цепи простираются в поле потока, эти атомы толкаются молекулами воды со стабильной положительной силой ~ 400 пН. Боковые цепи тянут позвоночник вперед, что увлекает за собой молекулы воды во внутреннем круге, что приводит к стабильной отрицательной силе примерно -200 пН.В SMD, поскольку сила прилагается непосредственно к дальнему концу петли, взаимодействие воды с белком вызывается движением атомов белка в результате удлинения петли при растяжении, которое имеет чистое значение, равное нулю. . Сила положительна на боковых цепях и отрицательна на основной цепи: обе уменьшаются почти до нуля после моделирования в течение 15 нс, когда атомы белка прекращают движение, потому что дальний конец β-переключателя вытянут почти как можно дальше вправо. Таким образом, в SMD сосредоточенная сила, приложенная к двум атомам основной цепи, распространяется в основном вдоль основной цепи.Напротив, в FMD силы сопротивления, действующие через проточную воду, распределяются по боковым цепям, а также по основной цепи. Поскольку образование β-шпильки требует движения боковых цепей, поток гораздо более эффективен, чем сосредоточенная сила, чтобы вызвать конформационный переход.
Рис. 4.Силы взаимодействия β-переключателя с окружающей водой. Силы взаимодействия между боковыми цепями, основной цепью или полным β-переключателем (показаны) и окружающей водой, рассчитанные на основе моделирования FMD ( A ) и SMD ( B ).
Обсуждение
Целью этого исследования было выяснить структурную основу мутаций GOF и LOF в области β-переключателя GPIbα. Однако сравнение равновесных структур и энергетики комплексов GPIbαN: A1 с WT и мутантными β-переключателями оказалось неубедительным. Хотя на первый взгляд это может показаться удивительным, этот результат согласуется с экспериментом. Сходные энергии взаимодействия предсказывают сходное сродство и стабильность связей VWF-A1 с WT и мутантным GPIb по GOF / LOF.Это предсказание подтверждается экспериментальным наблюдением, согласно которому по сравнению с WT мутация M239V увеличивает сродство связывания для A1 в 6 раз, но скорость отклонения мало изменяется (14). Таким образом, фенотипические различия между мутантами WT и GOF / LOF заключаются в нормах включения. Но какова структурная основа курсовой разницы?
Чтобы ответить на этот вопрос, мы рассмотрели различные конформации β-переключателя в не связанном (рис. 1 A ) и связанном (рис. 1 B ) GPIbα (15) требования к потоку для тромбоцитов, чтобы присоединиться к VWF, и изменение этого требования мутациями β-switch (5).Эти соображения привели нас к гипотезе о том, что поток увеличивает скорость связывания GPIbα-VWF, индуцируя переход от петли к β-шпильке в области β-переключателя. Важно отметить, что эта гипотеза предполагает динамическую, а не статическую структурную основу для различий в скорости между мутантами WT и GOF / LOF. Динамическая структура может быть стабильной, но требует отклонения от равновесия. Неравновесные процессы могут быть вызваны механическими возмущениями, такими как сила и поток. Действительно, недавнее исследование продемонстрировало, что сила замедляет скорость диссоциации GPIbα-VWF (необычная характеристика, называемая захватывающими связями), которая лежит в основе прокатки тромбоцитов на VWF с усилением потока и предотвращает агглютинацию тромбоцитов (21).Мутации GOF в домене A1 вдали от сайта связывания устраняют улавливающие связи, устраняют потребность в потоке для перекатывания и вызывают агглютинацию тромбоцитов. Моделирование SMD предлагает структурную основу для этого, так что сила наклоняет сложную структуру, чтобы позволить скольжение и повторное связывание на границе связывания, тем самым замедляя скорость диссоциации. Для сравнения, здесь мы предполагаем, что поток увеличивает скорость ассоциации, вызывая переход петля-шпилька в области β-переключателя. Эти два механизма дополняют друг друга в том, что они регулируют различные аспекты кинетики связывания GPIbα-VWF, которые лежат в основе различных аспектов адгезии тромбоцитов с усилением потока к VWF.Следует отметить, что, хотя наши симуляции не получили поддержки для механизма «подгонки, вызванной связыванием», этот механизм нельзя исключить из-за ограниченного времени симуляций.
Чтобы проверить нашу гипотезу, мы разработали новый алгоритм MD, названный FMD, для моделирования влияния потока жидкости на конформацию белка. Наша гипотеза была подтверждена моделированием FMD перехода петля-шпилька на β-переключателе, который наблюдался при двух скоростях потока, предполагая, что пониженная скорость продлит время, но все же вызовет переход.По сравнению с WT, мутант GOF M239V и мутант LOF A238V, как наблюдали, соответственно уменьшали и увеличивали потребность в потоке для перехода петля-шпилька, что согласуется с экспериментом. Тот факт, что эти ожидаемые различия все еще наблюдались, придает уверенность нашим результатам, несмотря на тот факт, что они были получены путем моделирования с гораздо более высокими скоростями, чем реальность, чтобы сократить время перехода от петли к шпильке до наносекундного диапазона, доступного для наших вычислительных ресурсов.
Насколько точно наши in silico симуляции изолированного β-переключателя имитируют физиологические условия? Чтобы ответить на этот вопрос, необходимо поместить β-переключатель в контекст всего GPIbα и тромбоцита.Когда плазма течет по пластинке, она прикладывает силу сдвига, чтобы сгибать длинный стержень GPIbα в направлении потока. Белковая часть U-образного GPIbαN почти симметрична относительно его длинной оси, соединяющей N- и C-концы, которая, вероятно, совпадает с направлением потока (рис. 5). Кроме того, поток оказывает сопротивление двум потенциальным N-гликанам на Asn-21 и Asn-159 (оранжевый на рис. 5 A ), расположенных на одной стороне этой оси, что, вероятно, немного отклонит эту сторону от оси. направление потока, чтобы выровнять β-переключатель с потоком, что делает его идеальным датчиком потока (рис.5 А ). Помимо заякоренных остатков 227–228 и 241–243, последовательность β-switch мало взаимодействует с остальным GPIbαN как в петле, так и в конформации β-шпильки. Следовательно, он должен быть относительно гибким, и на его форму можно легко влиять потоком. Эти аргументы изображены на рис. 5, где показаны конформации петли (зеленый) и β-шпильки (красный). Они расширяют нашу модель, включая выравнивание потока GPIbαN в качестве предварительного условия для индуцированного потоком перехода петля-шпилька на β-переключателе.Выравнивание потока GPIbαN зависит от того, как GPIbαN закреплен на ножке GPIbα, что, в свою очередь, зависит от конформации анионной последовательности (остатки 269–288). В структуре сокристалла анионная последовательность не была решена, что свидетельствует об отсутствии прямого контакта с A1 (14, 15). Эта последовательность (голубая трубка) наблюдалась в другой нелигандированной структуре GPIbαN, где она взаимодействовала с С-концевой фланкирующей последовательностью (охра) через три сульфатированных тирозина (красные палочки) и другие отрицательно заряженные остатки в анионной последовательности (рис.5) (11). Кажется разумным предположить, что взаимодействия между анионной последовательностью и GPIbαN регулируют выравнивание потока GPIbαN (рис. 5). Эта модель может объяснить экспериментальное наблюдение, что замена трех тирозинов на Phe (но не на Glu) снижает адгезию клеток, экспрессирующих мутантный GPIbα, к иммобилизованному VWF в потоке (22), что ранее было необъяснимо, поскольку эти остатки находятся далеко от VWF. -A1 сайт связывания. Более того, наша модель предполагает, что два потенциальных N-гликана на GPIbαN могут влиять на выравнивание потока GPIbαN, так что дегликозилирование GPIbα может увеличивать потребность в потоке для связывания VWF.Таким образом, модель индуцированного потоком перехода петля-шпилька поддерживается моделированием MD и объясняет усиленное потоком связывание тромбоцитов и фенотипы мутантов GOF и LOF в β-переключателе. Расширенная модель с выравниванием потока дополнительно объясняет фенотипы мутантов тирозина и обеспечивает проверяемый прогноз относительно эффекта дегликозилирования, который мотивирует будущие эксперименты.
Рис. 5.Модель для выравнивания потока GPIbαN. Прикрепленный к концу длинной ножки, N-концевой домен GPIbα простирается на ~ 60 нм от мембраны тромбоцитов, что делает его идеальным датчиком потока, когда тромбоциты циркулируют в кровотоке.В зависимости от якоря GPIbαN-стебля поток может оказывать разные силы на U-образный белок (синий) и наличие / отсутствие двух потенциальных N-гликанов (оранжевый) в Asn-21 и Asn-259 для выравнивания GPIbαN с потоком. направление (обозначено) по-разному, как показано в A и B , соответственно. Якорь опосредуется контактами анионной последовательности (голубой) с другой частью С-концевой фланкирующей последовательности (охра) через три сульфатированных тирозина (красные палочки), которые, вероятно, влияют на выравнивание GPIbαN и, в свою очередь, Область β-переключателя в поле течения.Наша модель предполагает, что выравнивание в A раскрывает β-переключатель [нарисованный в конформациях петля (зеленый) и β-шпилька (красный)] потоку потока, который вызывает переход петля-шпилька, тем самым усиливая эффект включения. скорость связывания GPIbα-VWF. Мутации, которые заменяют три сульфатированных тирозина или удаляют N-гликаны, могут изменять выравнивание GPIbαN с потоком, как показано в B , что влияет на воздействие потока на β-переключатель, тем самым изменяя усиление потока связывания GPIbα-VWF. .
Методы
Системы, используемые в нашем моделировании, включают GPIbαN в комплексе с A1 и изолированную последовательность β-переключателя. Использовали четыре структуры β-переключателя (остатки 227–243): последовательность WT в конформации β-шпильки [взято из базы данных Protein Data Base (PDB), код 1SQ0] (15), последовательность WT в конформации петли (взято из PDB code 1QYY) (23) и смоделированные последовательности M239V и A238V в конформации петли. Методы моделирования GPIbαN: A1 с этими последовательностями подробно описаны в SI Methods .Для моделирования с изолированным β-переключателем каждую последовательность плюс ион Cl — (для нейтрализации системы) вымачивали в водяной камере размером 64 × 40 × 40 Å таким образом, чтобы (который должен был образоваться) β- шпилька будет выровнена по оси x под потоком (также по направлению x ) (Рис. S7 A и B ). Эти структуры послужили отправной точкой для моделирования МД с NAMD (24) и силовым полем CHARMM22 (25). Структуры и фильмы S1 – S7 были созданы с использованием VMD (26).
Во всех моделированиях N-концевые C α атомов остатков 227 и 228, а также C-концевые C α атомов остатков 242 и 243 были зафиксированы для закрепления последовательности. Использовались периодические граничные условия. Обрезка 12 Å с плавным переключением от 10,5 Å использовалась для ван-дер-ваальсовых взаимодействий. Для полного электростатического взаимодействия использовался метод Эвальда с частицами-сеткой. Система сначала была минимизирована по энергии для 5000 шагов с фиксированными тяжелыми атомами пептида, а затем еще 5000 шагов с фиксированными только концевыми C α атомами.После повышения температуры до 300 К за 50 пс системы были подвергнуты уравновешиванию в течение 1,5 нс в ансамбле NPT с использованием поршневого метода Ланжевена для управления давлением и динамики Ланжевена для управления температурой. Коэффициент Ланжевена уменьшался каждые 0,5 нс с 5,1 до 0,1 пс -1 . Уравновешенные системы на 1,5 нс были взяты для дальнейшего моделирования. Количество независимых прогонов с различными методами МД для разных систем сведено в Таблицу S1.Для каждой системы были использованы независимые процессы минимизации, нагрева и уравновешивания для разных прогонов, чтобы гарантировать, что уравновешенные структуры не коррелированы, чтобы обеспечить значимую статистику.
Для FMD сила 0,7 или 0,35 пН прикладывается в направлении x к каждому из ∼300 атомов в слое 2 Å плоскости yz (показаны в виде больших сфер на рис. S7 A ). Эта сила ускоряет молекулы воды, которые позже текут вправо и над пептидом (рис.S7 B ). Из-за периодических граничных условий после выхода из водяного бокса на правой границе молекулы воды снова появляются в соответствующих положениях на левой границе. Когда они снова входят в слой 2 Å, к ним снова прикладывается сила. Таким образом, поток создается с возрастающей скоростью с течением времени. Скорость зависит от времени и расстояния от оси x , как показано на рис. S7 C и D . Увеличение скорости потока с увеличением времени ограничивает время моделирования, потому что скорость становится слишком большой, а температура системы также увеличивается до 350 K через 6 нс, когда 0.Применяется сила 7 пН (рассчитанная на основе кинетической энергии), что приводит к нестабильности системы при длительном моделировании. В отдельной статье (19) описан улучшенный алгоритм FMD, который позволяет проводить моделирование при постоянном среднем расходе и постоянной температуре в течение гораздо более длительного времени, что подтвердило результаты настоящего метода. Мы не наблюдали значительных различий в частоте образования водородных связей или их продолжительности между FMD и свободным моделированием MD.
Благодарности
Благодарим С.Харви, К. Шультен, З. Чен, Р. П. Макэвер, Дж. А. Лопес, Л. В. Макинтайр и Р. М. Нерем за полезное обсуждение. Вычислительные ресурсы для моделирования MD были предоставлены Лабораторией интерактивных высокопроизводительных вычислений вычислительного колледжа Технологического института Джорджии. Эта работа была поддержана грантом HL0 Национального института здоровья (C.Z.) и грантом Американской кардиологической ассоциации (J.L.).
Footnotes
- § Кому следует направлять корреспонденцию.Эл. Почта: cheng.zhu {at} bme.gatech.edu
Вклад авторов: J.L. и C.Z. спланированное исследование; J.L. провел исследование; J.L. проанализировал данные; и J.L. и C.Z. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/cgi/content/full/0801965105/DCSupplemental.
- © 2008 Национальная академия наук США
Короткая шпильочная РНК (shRNA)
Последовательности короткой шпилечной РНК (кшРНК) кодируются в ДНК-векторе, который может быть введен в клетки посредством плазмидной трансфекции или вирусной трансдукции.Поскольку кассеты экспрессии shRNA могут быть включены в вирусные векторные системы, включая лентивирус, они могут интегрироваться в геном хозяина для создания стабильных клеточных линий. Кроме того, при использовании в сочетании с одной из нескольких систем доставки вирусов они могут быть доставлены в трудно трансфицируемые первичные клетки и использованы для приложений in vivo. Основываясь на способе доставки и конструкции вектора, кшРНК на основе вектора могут обеспечивать долгосрочное (или индуцируемое) подавление генов-мишеней.
На эффективность shRNA влияют многие факторы, включая эффективность трансдукции или трансфекции, промотор, управляющий экспрессией shRNA и эпигенетические модификации (которые могут приводить к подавлению экспрессии shRNA). Кроме того, влияние каждого из этих факторов на производительность вектора может различаться в зависимости от линии клеток или типа клеток. При планировании эксперимента с использованием shRNA доступны варианты вектора, в том числе; используемый дизайн shRNA, векторные особенности (например,g., промоутер), а также следует учитывать способ доставки, исходя из требований эксперимента.
дизайн shRNA
КонструкциякшРНК обычно делится на два формата: простая кшРНК типа стержень-петля и адаптированная к микроРНК кшРНК.
Простая кшРНК типа стержень-петля
Основные кшРНК моделируются на основе микроРНК-предшественника (пре-миРНК) и клонируются в вирусные векторы, где они транскрибируются под контролем промоторов РНК-полимеразы III (Pol III).shRNA продуцируются как одноцепочечные молекулы длиной 50–70 нуклеотидов и образуют структуры петли стебля, состоящие из области из 19–29 пар оснований двухцепочечной РНК (ножки), соединенной областью одноцепочечной РНК ( петлю) и короткий свес 3 ‘. После транскрибирования кшРНК покидают ядро, расщепляются в петле нуклеазой Dicer в цитоплазме и попадают в RISC, чтобы направлять расщепление и последующую деградацию комплементарной мРНК.
кшРНК, адаптированная к микроРНК
КшРНК, адаптированная к микроРНК, состоит из стволовой структуры кшРНК с микроРНК-подобными ошибочными спариваниями, окруженными петлей и фланкирующей последовательностью эндогенной микроРНК.адаптированные к микроРНК shRNA транскрибируются с промоторов РНК-полимеразы II (Pol II), расщепляются эндогенным ферментом Drosha РНКазы III в ядре, а затем экспортируются в цитоплазму, где они обрабатываются Dicer и загружаются в комплекс RISC. Исследования показали, что использование каркаса микроРНК, который обрабатывается как Drosha, так и Dicer, может способствовать более эффективному процессингу и снижению токсичности для РНКи in vivo.
РНК-интерференция и манипуляции
Рисунок 1.Подходы shRNA включают введение генно-инженерных вирусных векторов или векторов на основе плазмид, экспрессирующих последовательности сайленсинга, встроенные в каркас эндогенной микроРНК (1) или простую shRNA типа стержень-петля (2). Экспрессированные последовательности (1 и 2, показаны синим) входят в эндогенный путь на ранней стадии и эффективно преобразуются в мощные молекулы, подавляющие звук, с использованием механизма эндогенной микроРНК. Все эти подходы приводят к расщеплению целевой мРНК (показано пурпурным цветом) и замалчиванию генов.
Список литературы и рекомендуемая литература
- Liu, Z. et al. (1997) Систематическое сравнение относительной активности промотора / энхансера в клеточных линиях млекопитающих. Analytical Biochem. 246 (1): 150-152.
- Ramezani, A. et al. (2000) Лентивирусные векторы для повышенной экспрессии генов в гемопоэтических клетках человека. Молекулярная терапия 2 (5): 458-469.
- Brummelkamp, T.R. и другие. (2002) Система стабильной экспрессии коротких интерферирующих РНК в клетках млекопитающих. Science 296 (5567): 550-553.
- Paddison, P.J. et al. (2002) Стабильное подавление экспрессии генов с помощью РНКи в клетках млекопитающих. PNAS 99 (3): 1443-1448.
- Пол, К. и другие. (2002) Эффективная экспрессия малых интерферирующих РНК в клетках человека. Природная биотехнология 20 (5): 505-508.
- Бартель, Д.П. (2004) МикроРНК: геномика, биогенез, механизм и функция. Ячейка 116 (2): 281-297.
- Gregory, R.I. et al. (2005) Human RISC сочетает биогенез микроРНК и посттранскрипционное молчание генов.Ячейка 123 (4): 631-640.
- Ким, В. (2005) Биогенез микроРНК: скоординированное выращивание и нарезание кубиками. Обзоры природы, молекулярная клеточная биология 6 (5): 376-385.
- Silva, J.M. et al. (2005) Библиотеки shRNA второго поколения, охватывающие геномы мыши и человека. Nature Genetics 37 (11): 1281-1288.
- Siolas, D. et al. (2005) Синтетические кшРНК как мощные триггеры РНКи. Природная биотехнология 23 (2): 227-231.
- Grimm, D. et al. (2006) Смерть у мышей из-за перенасыщения клеточных путей микроРНК / коротких шпилечных РНК.Nature 441: 537-541.
- Hong, S. et al. (2007) Функциональный анализ различных промоторов в лентивирусных векторах на разных стадиях дифференцировки эмбриональных стволовых клеток мыши in vitro. Молекулярная терапия 15 (9): 1630-1639.
- Luo, B. et al. (2008) Высоко параллельная идентификация основных генов в раковых клетках. PNAS 105 (51): 20380-20385.
- McBride, J.L. et al. (2008) Искусственные miRNAs уменьшают shRNA-опосредованную токсичность в головном мозге: значение для терапевтического развития RNAi.PNAS 105 (15): 5868-5873.
- Schlabach, M.R. et al. (2008) Открытие гена пролиферации рака с помощью функциональной геномики. Наука 319 (5863): 620-624.
- Silva, J.M. et al. (2008) Профилирование основных генов в клетках молочной железы человека с помощью множественного скрининга РНКи. Наука 319 (5863): 617-620.
- Zender, L. et al. (2008) Онкогеномный скрининг in vivo RNAi выявляет опухолевые супрессоры при раке печени. Ячейка 135 (5): 852-864.
- Gumireddy, K. et al.(2009) KLF17 является негативным регулятором эпителиально-мезенхимального перехода и метастазирования при раке молочной железы. Природа клеточной биологии 11 (11): 1297-1304.
- Maier, B. et al. (2009). Широкомасштабный функциональный скрининг РНКи показывает роль CK2 в циркадных часах млекопитающих. Гены и развитие 23: 708-718.
- Mullenders, J. et al. (2009) Кандидаты в биомаркеры ответа на экспериментальное противораковое лекарство, идентифицированные с помощью крупномасштабного генетического скрининга РНК-интерференции. Клинические исследования рака 15 (18): 5811-5819.
- Peng, J. et al. (2009) Jarid2 / Jumonji координирует контроль ферментативной активности PRC2 и занятость целевого гена в плюрипотентных клетках. Cell 139 (7): 1290-1302.
- Йунг, М.Л. и другие. (2009) Скрининг короткой шпильочной РНК по всему геному Т-клеток jurkat на предмет белков человека, способствующих продуктивной репликации ВИЧ-1. Журнал биологической химии 284 (29): 19463-19473.
- Beer, S. et al. (2010) Низкий уровень цитотоксичности shRNA может способствовать развитию гепатоцеллюлярной карциномы, индуцированной MYC, у взрослых мышей.Молекулярная терапия 18 (1): 161-170.
- Du, W. et al. (2010) Цитоплазматический комплекс FANCA-FANCC взаимодействует и стабилизирует цитоплазматический лейкозный белок нуклеофозмина (NPMc). Журнал биологической химии 285 (48): 37436-37444.
- Rato, S. et al. (2010) Новые мишени для нокдауна ВИЧ-1, идентифицированные с помощью библиотеки shRNA, обогащенной киназами / фосфатазами, с использованием долгосрочного итеративного скрининга на Т-клетках Jurkat. PLoS One 5 (2): e9276.
- Смолен, Г.А. и другие. (2010) Полногеномный скрининг РНКи выявляет множественные RSK-зависимые регуляторы клеточной миграции.Гены и развитие 24 (23): 2654-2665
- Fellmann, C. et al. (2011) Функциональная идентификация оптимизированных триггеров РНКи с использованием массивно-параллельного сенсорного анализа. Молекулярная ячейка 41 (6): 733-746.
- Montgomery, R.L. et al. (2011) Терапевтическое ингибирование miR-208a улучшает сердечную функцию и выживаемость при сердечной недостаточности. Тираж 124 (14): 1537-1547.
- Pan, Q. et al. (2011) Нарушение пути микроРНК из-за широко используемых лентивирусных библиотек shRNA ограничивает применение для скрининга факторов хозяина, участвующих в инфицировании вирусом гепатита С.FEBS Letters 585: 1025-1030.
- Ying, M. et al. (2011) Kruppel-подобное семейство фактора транскрипции 9, фактора транскрипции, ассоциированного с дифференцировкой, подавляет передачу сигналов Notch2 и ингибирует стволовые клетки, инициирующие глиобластому. Стволовые клетки 29 (1): 20-31.
Какой реагент shRNA лучше всего подходит для вас?
Используйте приведенную ниже таблицу, чтобы помочь вам выбрать подходящие реагенты shRNA для ваших экспериментальных нужд.
| SMART вектор шРНК | ГИПЗ | TRIPZ | TRC | |
|---|---|---|---|---|
| Виды | Человек, Мышь, Крыса | Человек, Мышь | Человек | Человек, Мышь |
| Промоутер | Выбор из 7 конститутивных * и 4 индуцибельных промоторов | CMV Pol II человека | TRE-min-CMV | U6 Pol III |
| Магистраль вектора | Лентивирал | Лентивирал | Лентивирал | Лентивирал |
| Формат стержня-петли | Адаптированная к микроРНК | Адаптированная к микроРНК | Адаптированная к микроРНК | Простой |
| Флуоресцентный репортерный ген | GFP / RFP | GFP | ЗП | |
| Гарантированное глушение ** | ||||
| РНКи in vivo | ||||
| Создание стабильных клеточных линий | ||||
| Рекомендовано для первичных и неделящихся клеток | ||||
| Индуцируемая экспрессия | ||||
| Доступность библиотеки полного генома | Человек, Мышь, Крыса | Человек, Мышь | Человек | Человек |
| Форматы | Бактериальный запас глицерина Высокотитровые лентивирусные частицы Семейства генов и пути пути | Бактериальный глицериновый запас Формат библиотеки с матрицами Семейства и пути генов Лентивирусные частицы с высоким титром Стартовый набор shRNA | Бактериальный глицериновый запас Формат библиотеки с матрицами Семейства генов и пути Стартовый набор shRNA | Бактериальный глицериновый запас Формат матричной библиотеки Семейства и пути генов |
* Некоторые варианты промоутеров могут быть доступны только в виде индивидуальных продуктов или по запросу.
** Для лентивирусных shРНК SMARTvector, GIPZ и TRIPZ по крайней мере одна из трех конструкций гарантированно снижает уровни целевой мРНК на 70% или более при использовании в сочетании с соответствующим образом подобранным нецелевым и положительным контролями.
shRNA контролирует
Критически важный для любого эксперимента по подавлению гена, контроль shRNA обеспечивает точную интерпретацию для получения надежных, воспроизводимых результатов
Массивные библиотеки shRNA
Поставляется в виде глицерина в 96-луночном формате; Эти библиотеки идеальны для групповых форматов скрининга для выяснения активности генов и изучения конкретных путей или семейств генов.
Объединенные библиотеки shRNA
Объединенный лентивирусный скрининг может быть проведен для идентификации генов, регулирующих клеточные ответы и сигнальные пути, или для обнаружения новых функций генов. Объединенные библиотеки скрининга могут состоять от 50 до многих тысяч. В отличие от дорогостоящих автоматизированных методов, которые требуются для скрининга с использованием индивидуально расположенных реагентов, объединенные скрининговые библиотеки позволяют исследователю преобразовывать и скринировать популяцию клеток в нескольких чашках для культур тканей.
Объединенные библиотеки лентивирусных shRNA SMARTvector
Объединенные скрининговые библиотеки с высоким титром конститутивных лентивирусных кшРНК SMARTvector для предварительно определенных библиотек генов у человека, мыши и крысы. Выберите из семи промоутеров и трех вариантов репортеров, чтобы выбрать лучший формат для своего эксперимента.
Объединенные библиотеки индуцибельной лентивирусной shRNA с вектором SMART
Объединенные скрининговые библиотеки с высоким титром индуцируемых лентивирусных кшРНК SMARTvector для предварительно определенных библиотек генов у человека, мыши и крысы.Выберите из четырех промоутеров и двух вариантов репортера, чтобы выбрать лучший формат для своего эксперимента.