плотность и содержание сухого вещества
Please use this identifier to cite or link to this item: https://elar.usfeu.ru/handle/123456789/10022
| Title: | Квалиметрия фитомассы лесных деревьев: плотность и содержание сухого вещества |
| Other Titles: | Qualimetry of forest tree biomass: density and dry matter content |
| Authors: | Усольцев, В. А. Цепордей, И. С. |
| Issue Date: | 2020 |
| Citation: | Усольцев, В. А. Квалиметрия фитомассы лесных деревьев: плотность и содержание сухого вещества = Qualimetry of forest tree biomass: density and dry matter content : монография / В. А. Усольцев, И. С. Цепордей ; Министерство науки и высшего образования Российской Федерации, Уральский государственный лесотехнический университет. – Екатеринбург, 2020. – 178 с. : ил.– Библиогр. с. 119–176. – Текст : электронный. |
| Abstract: | В необъятном море экспоненциально нарастающей информации важно вычленить и обосновать перспективные направления, научные ориентиры-маяки, ускоряющие прогресс в той или иной области науки, при минимизации потерь от дублирования и следования тривиальной методологии проб и ошибок. In the vast sea of exponentially growing information, it is important to identify and justify promising directions, scientific landmarks-beacons that accelerate progress in a particular field of science while minimizing losses from duplication and following a trivial methodology of trial and error. 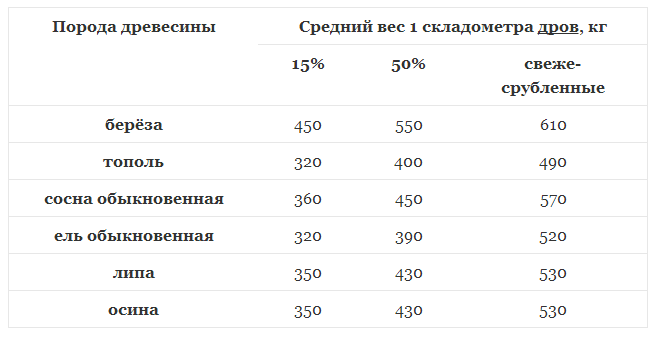 As can be seen in the proposed analysis of problems related to the assessment of qualimetric properties of wood, the number of publications on this topic has been growing catastrophically in recent years, and methods and algorithms for modeling them are steadily improving. However, in the course of solving a particular problem, a lot of uncertainties are found that require a correct solution. The proposed review is devoted to an attempt to systematize both the results obtained by different authors and the uncertainties generated by them. For professionals in the development and management of forestry and forest inventory, to develop systems for monitoring forest and environmental programs in different forest science areas. As can be seen in the proposed analysis of problems related to the assessment of qualimetric properties of wood, the number of publications on this topic has been growing catastrophically in recent years, and methods and algorithms for modeling them are steadily improving. However, in the course of solving a particular problem, a lot of uncertainties are found that require a correct solution. The proposed review is devoted to an attempt to systematize both the results obtained by different authors and the uncertainties generated by them. For professionals in the development and management of forestry and forest inventory, to develop systems for monitoring forest and environmental programs in different forest science areas. |
| Keywords: | КВАЛИМЕТРИЯ ФИТОМАССЫ ДЕРЕВА БАЗИСНАЯ ПЛОТНОСТЬ СОДЕРЖАНИЕ СУХОГО ВЕЩЕСТВА ФОРМООБРАЗОВАНИЕ СТВОЛА ДРЕВЕСИНА И КОРА ВЕТВЕЙ КАЧЕСТВО ДРЕВЕСНЫХ ВОЛОКОН ДИСТАНЦИОННОЕ ЗОНДИРОВАНИЕ РАЗВИТИЕ КРОНЫ |
| URI: | https://elar. usfeu.ru/handle/123456789/10022 usfeu.ru/handle/123456789/10022 |
| ISBN: | 978-5-94984-768-8 |
| Appears in Collections: | Монографии |
Show full item record Google Scholar
Items in DSpace are protected by copyright, with all rights reserved, unless otherwise indicated.
Древесина плотность
Плотность стволовой древесины. Величина плотности стволовой древесины зависит от ее породы, влажности и коэффициента разбухания /Ср. Все породы древесины по отношению к коэффициенту разбухания КР разделяются на две группы. К первой группе относятся породы, у которых коэффициент разбухания /СР = 0,6 (белая акация, береза, бук, граб, лиственница). Ко второй группе относятся все остальные породы, у которых /[ …]
Плотность древесины — это масса древесины, заключающаяся в единице объема, например в одном кубическом сантиметре или в одном кубическом метре. В первом случае плотность измеряется в граммах на 1 см3, во втором — в тоннах на 1 м3. [ …]
[ …]
Прочность древесины приблизительно пропорциональна ее плотности, что подтверждают проведенные испытания на определение модуля упругости древесины и наблюдения при испытании на сжатие вдоль волокон. Испытание других свойств древесины, не учитывающих разновидность пород, приводит при расчете по специальным показательным уравнениям к средним цифрам, которые могут значительно расходиться с цифрами, полученными при конкретном испытании. Фактически разница между расчетом и цифрами может доходить до 20%. Тем не менее расчетные уравнения дают возможность получить приближенные данные прочности древесины, объемный вес которой известен.[ …]
Древесина — естественный полимер, состоящий из клеток-волокон, имеющих трубчатую форму и направленных вдоль ствола. Благодаря этому древесина обладает целым рядом достоинств: высокой прочностью, упругостью, малой плотностью, а следовательно, и малым весом, низкой теплопроводностью, тонкостью к воздействию химически агрессивных сред, природной декоративностью, простотой обработки и монтажа. [ …]
[ …]
Древесина деревьев разных пород обладает различными физическими и механическими свойствами. Например, тополь хорошо проводит электричество, а сухую древесину дуба можно назвать даже диэлектриком: не так давно радиолюбители делали из нее панели простейших радиоприемников. Технические свойства даже одного дерева неодинаковы у корней (в комле) и у кроны. Плотность и прочность, например, сосны увеличивается от ядра к заболонной части (под корой) в 2 раза.[ …]
Древесина березы отличается высокой прочностью, особенно при ударных нагрузках; однородностью строения и цвета, средней плотностью и твердостью, но малостойкая против гниения.[ …]
Древесина сосны сибирской отличается от древесины сосны обыкновенной меньшей плотностью и легкостью в обработке, отчего используется, в частности, при изготовлении карандашей.[ …]
Плотность древесинного вещества — это отношение массы материала, образующего стенки клеток, к занимаемому им объему. Плотность древесинного вещества одинакова для всех пород древесины и равна 1,53 г/см3. [ …]
[ …]
Плотность элементов кроны деревьев. Плотность элементов кроны практически не изучена. В топливной щепе из элементов кроны преобладающим по объему компонентом является щепа из сучьев и ветвей, близкая по показателям плотности к стволовой древесине. Поэтому при проведении практических расчетов в первом приближении можно принять плотность элементов кроны равной плотности стволовой древесины соответствующей породы.[ …]
Часть древесины отходов отправляют на производство топливных брикетов. Они транспортабельны, удобны в применении. Их характеристика: плотность 800-1100 кг/м3, влажность [ …]
Значения плотности основных пород приведены в таблице 2. Влажность — физическое свойство древесины, характеризующееся количеством содержащейся в ней влаги. Микроструктура древесных волокон такова, что влага лучше всего проникает через торцевые поверхности.[ …]
Величина плотности при стандартной влажности определяется для различных пород древесины по табл. 6.[ …]
Величина плотности древесины зависит от ее породы и влажности. [ …]
[ …]
Неодинаковый вес древесины разных пород зависит исключительно От больш или меньшего содержания воды и различной плотности строения древесины. Дре сица старого дерева, вообще, тяжелее, чем—молодого; древесина ствола тяжелее х весины ветвей; дерево, выросшее на тощей и сухой почве, тяжелее выросшего сырой и жирной почве.[ …]
Строение и свойства древесины влияют на качество, а не на высоту звука. Затухание звука в древесине в противоположность металлам обусловлено не только междуатомным взаимодействием, по и силами межмолекулярного взаимодействия. Поэтому энергия звука рассеивается очень быстро. Качество звука в музыкальном смысле определяется обертонами гармоник или высшей частотой гармоник, которые придают деревянным инструментам их особую привлекательность. Для достижения наилучших результатов древесина, используемая для музыкальных инструментов, должна быть однородной по строению и по содержанию влаги, свободной от дефектов или от внутренних напряжений, иначе говоря, от любых изменений в строении (однородность, плотность) или качестве, могущих вызвать искажение колебания [13].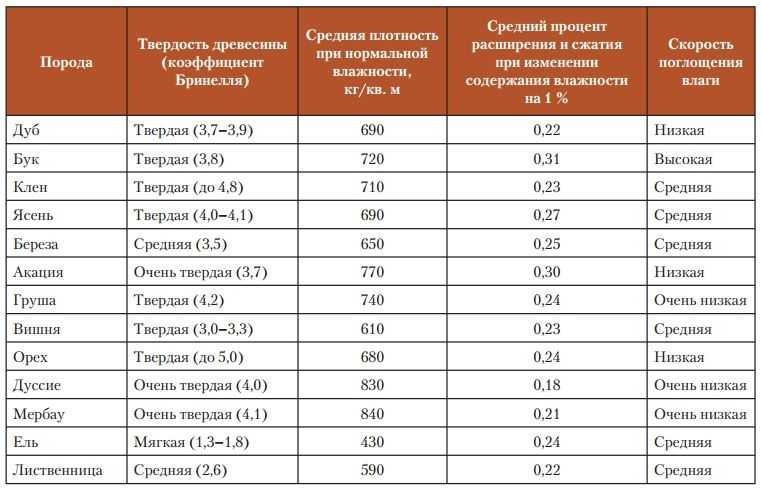 [ …]
[ …]
Зависимость прочности древесины от содержания влаги. Так как прочность и жесткость древесины частично определяются силами сцепления, связывающими молекулы, то любой агент, уменьшающий эти силы, меняет ее прочность в целом. Одним из таких агентов является вода, поэтому прочность древесины увеличивается по мере уменьшения содержания влаги не только в результате повышенной плотности, происшедшей от усушки, но также из-за присутствия вторичных валентных сил сцепления1. Так как присутствие воды в количестве, превышающем точку насыщения волокна, не изменяет характера клеточной стенки, то потеря или приобретение капиллярной (свободной) воды практически не влияет на показатели прочности древесины.[ …]
При ежегодной заготовке древесины в России на уровне 500 млн м3 общий выход ее отходов достигает 300 млн м3. Объем их использования, по данным А.Ф.Протасова и А.В.Молчанова, составляет порядка 46 млн м3, или примерно 15%. Таким образом, количество безвозвратно теряемой древесины, с учетом потерь при сплаве леса, заведомо превышает 50%, или, при ее плотности 0,6 т/м3, свыше 150 млн т/год (250 млн м3). [ …]
[ …]
При определении влияния плотности древесины на касательную составляющую силы резания Рх толщина стружки равнялась е = 0,02 мм. Была установлена линейная связь между плотностью и силой. После обработки результатов опытов получены следующие корреляционные уравнения связи: для сосны />1 = 1,04 коэффициент корреляции 0,92; для дуба Р 1,12 у, коэффициент корреляции 0,94. Полученные результаты следует учитывать при изучении процессов затупления резца и образования поверхности резания.[ …]
Хвойные породы имеют меньшую плотность древесины, а следовательно, и меньшую теплопроводность. Очевидно превосходство по теплопроводности дерева над кирпичом, поскольку кирпичные стенки толщиной 510 мм (в два кирпича) обладают такими же термоизоляционными свойствами, как и стена из деревянного бруса толщиной 100 мм. Понятно, что по стоимости эти материалы несравнимы. Кроме того, деревянные стены «накапливают» тепло и равномерно распределяют его по всему помещению. Поэтому в таком доме будет тепло даже в самый лютый мороз. Последнее обстоятельство становится решающим при выборе вида древесины для строительства деревянных домов.[ …]
Последнее обстоятельство становится решающим при выборе вида древесины для строительства деревянных домов.[ …]
При пиролизе указанных отходов древесины по известной технологии [93] образуется полидисперсный порошок с размерами частиц 0,3 — 0,7 мм. Сорбционная емкость такого нефтесорбента «Илокор» составляет 8,0 —8,8 г нефти на 1 г сорбента. Удельная поверхность сорбента, определенная методом ртутной порометрии, колеблется в пределах 2840 — 3660 мг/г. Плотность «Илокора» — 0,82 — 0,87 г/см1, насыпная масса — 82 кг/м3. Разработанный материал является экологически чистым, не оказывающим даже незначительного отрицательного влияния на все звенья экологической цепи природных экосистем, в первую очередь биологических объектов, вплоть до генетического уровня.[ …]
Путем искусственного уплотнения плотность укладки щепы повышается на 20—30%. При 25%-ном уплотнении и при тех же прочих условиях объем щепы составит: 0,35- 1,25 = 0,44 м3, а свободное пространство 1—0,44 = 0,56 м3. Однако количество кислоты определяется не только свободным объемом после заполнения котла щепой, но и объемом пор древесины, в которые впитывается варочная кислота. [ …]
[ …]
Отмечается достоверное увеличение плотности древесины по мере удаления от центра ядра к заболони в комлевой и средней частях ствола. Аналогично изменение сопротивления древесины сжатию вдоль волокон с той лишь разницей, что в области заболони отмечается незначительное увеличение технического показателя. По мере продвижения от комля к вершине происходит падение плотности, а затем вторичное ее увеличение в верхушечной части ствола.[ …]
Различную прочность заболони и ядровой древесины обычно можно объяснить различиями в плотности, содержании влаги, величине прироста, присутствии или отсутствии дефектов и экстрактивных веществ.[ …]
Это зависит от разности в сопротивлении древесины деформированию при сжатии вдоль волокон и поперек. При постановке дна поперек волокон происходит большая усадка доньев, а это отрицательно влияет на плотность доньев и их сопряжения с остовом.[ …]
Особняком стоит железная береза (В. Schmidtii Rgl), древесина которой отличается исключительной твердостью и плотностью. [ …]
[ …]
Наибольшее практическое использование находит древесина дуба, которая является важнейшим лесопильным материалом, применяемым для наиболее ответственных работ, где нужны прочность, твердость, упругость, стойкость к факторам внешней среды,— в судостроении, авиационной промышленности, транспорте, строительстве жилищ, разного рода столярных работах и т. д. Кроме того, древесина почти всех видов дубов дает прекрасное топливо, калорийность которого растет вместо с плотностью; дубовый древесный уголь отличается хорошим качеством. Наиболее твердую древесину дают виды подрода гетеробаланус, очень высокими эксплуатационными показателями характеризуется и древесина видов подрода ( иог-сия. Древесина красных американских дубов отличается от прочих розоватым или красноватым оттенком, она более мягкая. Однако в этом подроде также имеются виды с весьма ценной древесиной. Древесина видов подрода циклобалаиопсис в среднем по ¡эксплуатационным показателям гораздо хуже таковой видов умеренных областей — не очень твердая, колкая, часто коробится и повреждается насекомыми. [ …]
[ …]
Если скорость звука в воде (с) составляет 1,48 см/с, плотность воды (р) — 1г/см3, то звуковое давление будет равно 5,4 атм. Это означает, что в течение 1 с акустическое давление изменится 50 тысяч раз от -5,4 до +5,4 атм. Такое пульсирующее давление не может не оказывать положительного влияния на пропитку древесины защитными препаратами.[ …]
Таким образом, полуфабрикаты из лиственных пород древесины придают бумажной продукции ряд ценных свойств. Однако есть и недостатки при использовании лиственной древесины. Из-за повышенной плотности эта древесина в воде тонет, что исключает ее сплав. Обычный метод мокрой окорки для лиственной древесины не пригоден. Различия в химическом составе и морфологическом строении лиственных и хвойных пород древесины требуют их раздельной варки. Поэтому для лиственных пород древесины на целлюлозном заводе должен быть отдельный технологический поток варки, промывки, очистки и отбелки целлюлозы.[ …]
В сушильной практике пользуются понятием условная плотность 1и условная объемная масса), которая представляет собой отноше-г массы древесины в абсолютно сухом состоянии (кг) к ее объему ) при влажности выше предела гигроскопичности (т. е. до шки).[ …]
е. до шки).[ …]
Непосредственно образующиеся в процессе заготовки древесины и ее первичной обработки древесные отходы и щепа имеют малую плотность и низкую теплоту сгорания. Вследствие этого они как топливо малотранспортабельны, и при сжигании развивают недостаточно высокую температуру горения, что исключает их применение для ведения высокотемпературных процессов и снижает теплопроизводительность и КПД котельных установок, в которых они используются. Так, за рубежом проводятся исследования по разработке и совершенствованию производства транспортабельного, высококачественного топлива на базе древесного сырья. Применение древесных брикетов, по мнению специалистов, особенно привлекательно для бытового потребления, так как в их составе не содержится серы и других вредных элементов.[ …]
Выявлена функциональная зависимость сопротивления древесины сжатию и базисной плотности. При этом отмечена обратная зависимость сопротивления древесины и диаметра растущего дерева на высоте груди. [ …]
[ …]
Нами получено, вне зависимости от стадии разрушения древесины, закономерное снижение плотности древесины фаутных деревьев в сосняках черничных до 20 и в кустарничково-сфагновых — до 25%, что закономерно приводит к значимому снижению предела сопротивления древесины сжатию.[ …]
Средние статистические показатели для некоторых пород древесины в свежесрубленном состоянии, а также при влажности древесины, равной 12%, были установлены Лабораторией лесных продуктов. Так как прочность меняется в зависимости от плотности, объемный вес древесины дается для каждой серии испытаний отдельно.[ …]
Для измерения доступности нативной целлюлозы и образцов древесины в последние годы нередко используются методы дей-терирования и тритирования [298]. Иногда для изучения пористости целлюлозных волокон применяют метод рассеяния рентгеновских лучей под малыми углами [299—301], теория которого была разработана Породой. Была показана возможность абсолютных измерений удельных поверхностей и непосредственного определения пористости в пористых телах. Изучая целлюлозные волокна методом рентгеновского рассеяния под малыми углами, Статтон [239] присоединился к взгляду Порода, считающего, что основным источником диффузного рассеяния в сухом волокне являются не кристаллиты, а микропустоты, или области низкой электронной плотности. Кривые распределения показывают на наличие в целлюлозных волокнах пустот различных размеров, которые для вискозного волокна, например, лежат в интервале 20-280 к.[ …]
Изучая целлюлозные волокна методом рентгеновского рассеяния под малыми углами, Статтон [239] присоединился к взгляду Порода, считающего, что основным источником диффузного рассеяния в сухом волокне являются не кристаллиты, а микропустоты, или области низкой электронной плотности. Кривые распределения показывают на наличие в целлюлозных волокнах пустот различных размеров, которые для вискозного волокна, например, лежат в интервале 20-280 к.[ …]
Хотя условия и место произрастании могут влиять на прочность древесины, степень различия может оказаться не больше, чем для древесины, взятой из разных частей одного и того же дерева. Фепль [7] нашел, что свежесрубленпая древесина из ветвей тяжелее древесины из ствола, что древесина из ствола тяжелее корневой древесины, и что сопротивление сжатию вдоль и поперек волокон подчинено той же зависимости. При испытаниях на сжатие вдоль волокон высушенная древесина из ствола обладает большей прочностью, чем сравниваемая древесина из ветвей. Это вызвано увеличением плотности древесины ствола за счет усушки. Корневая древесина является самой легкой, слабой и гибкой, однако это не влияет на выполнение ею своей главной функции. Древесина в верхушечной части обычно слабее древесины в комле [8].[ …]
Корневая древесина является самой легкой, слабой и гибкой, однако это не влияет на выполнение ею своей главной функции. Древесина в верхушечной части обычно слабее древесины в комле [8].[ …]
Согласно мнению Маклина 134], теплопроводность абсолютно сухой древесины пропорциональна ее плотности в соответствии с уравнением.[ …]
Движение воздуха в вертикальном направлении обусловлено гостью плотностей воздуха в штабеле и вне его. Днем нагретый 1ух, поступая в штабель, охлаждается и движется вниз. Вечером эчью остывший воздух, попадая в штабель, сохраняющий бла-фя свойству древесины аккумулировать теплоту более высокую 1ературу, нагревается н движется вверх. В связи с этим создает-текоторое реверсирование движения воздуха в штабеле.[ …]
По результатам лабораторного контроля установлено, что даже в зонах с плотностью загрязнения почвы цезием-137 1—5 Ки/км2 содержание радионуклидов в грибах, лесных ягодах, травянистой растительности, хвое и листве деревьев, а также в древесине может выходить за приделы установленных нормативов.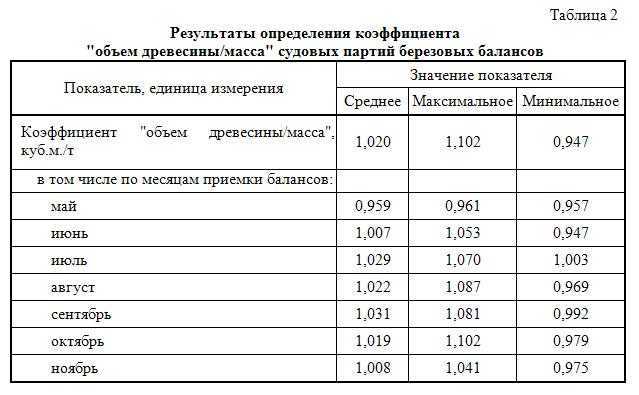 [ …]
[ …]
Если разрезать ствол не поперек, а вдоль, станут видны различные по ширине, цвету и плотности прожилки — сердцевинные лучи, пронизывающие годичные слои. По ним от камбия идут по всему стволу к ветвям и листьям питательные вещества, поступающие по корням из почвы. Рисунок, открывающийся на поверхности древесины после перерезания волокон годичных колец и питательных сердцевинных лучей, называют текстурой древесины. Чем разнообразнее этот рисунок по конфигурации и цветовым оттенкам, тем ценнее древесина как отделочный материал в столярном производстве.[ …]
Лиственница среди других хвойных пород занимает особое место. Это ядровая порода, ее древесина обладает высокой стойкостью против гниения и грибковых заболеваний. Древесина лиственницы обладает высокими физико-механическими свойствами — по плотности и прочности примерно на 30 % превосходит сосну.[ …]
Формула (3.1) является приближенной, так как влажность при точке насыщения стенок клеток и плотность древесинного вещества взяты средние.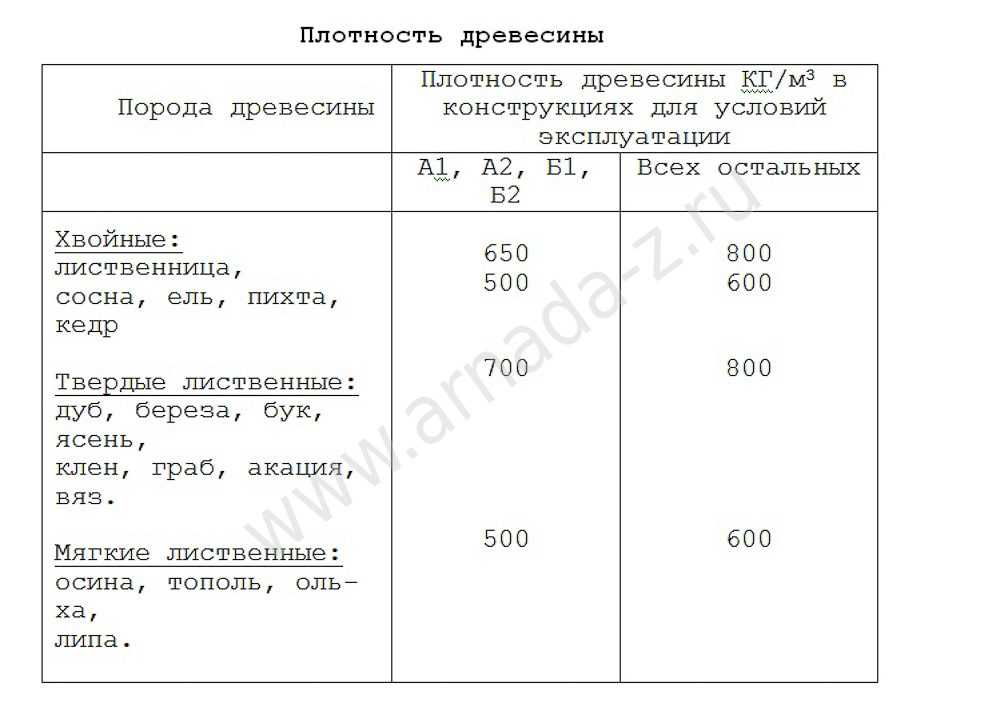 Кроме того, при выводе формулы сделано предположение, что все пустоты древесины заполняются влагой. В действительности в некоторую часть пустот вода проникнуть не может, поэтому результаты, полученные по формуле, будут превышать опытные данные. Однако для целей теплотехнических расчетов это имеет даже положительное значение — полученные результаты являются как бы предельными с точки зрения определения наихудших условий.[ …]
Кроме того, при выводе формулы сделано предположение, что все пустоты древесины заполняются влагой. В действительности в некоторую часть пустот вода проникнуть не может, поэтому результаты, полученные по формуле, будут превышать опытные данные. Однако для целей теплотехнических расчетов это имеет даже положительное значение — полученные результаты являются как бы предельными с точки зрения определения наихудших условий.[ …]
Вместе с тем необходимо отметить, что радиационная обстановка в лесах на площади 27,5 тыс.га, где имеет место все еще высокая плотность загрязнения почвы цезием-137( от 15 до 40 Кюри/кв.км), характеризуется мощностью дозы гамма-излучения от 0,7 до 2 мкЗв/с. В зоне отчуждения площади лесов, где мощность дозы составляет свыше 200 мкЗв/с, содержание радионуклидов в некоренной древесине достигает нескольких десятков тысяч Бк/кг. Особенно загрязнена кора лиственных пород (цо 150 тыс. Бк/кг). Такие участки составляют 2,2 тыс. га. На этих участках запрещены все виды лесопользования. [ …]
[ …]
В зависимости от размеров (дисперсности) частиц, их плотности, формы и скорости движения жидкости они могут всплывать (жиры, бумага, нефть, древесина и т. п.), находиться во взвешенном (суспензированном) состоянии в воде, осаждаться на дно сосуда и волочиться по дну в виде осадка (песок, куски веществ с плотностью больше 1 — металлическая окалина, каменный уголь и пр.).[ …]
Первое шлифование производят крупнозернистой шкуркой, заканчивают самой мелкозернистой. Шлифуют вдоль волокон. Но если поверхность изделия составлена из древесины разной плотности и эти составляющие имеют разное направление волокон, то шлифовка поперек волокон допускается для пород большей плотности.[ …]
При использовании рабочих водных растворов антисептика ЭОК с пониженной концентрацией токсичных ингредиентов и небольшим содержанием соды на двух предприятиях были зафиксированы случаи обрастания обработанной древесины грибами. Поэтому при антисептировании пиломатериалов целесообразно осуществлять непрерывный контроль качества рабочих растворов не только по средним показателям их плотности и pH среды, но и по концентрации основных ингредиентов препарата.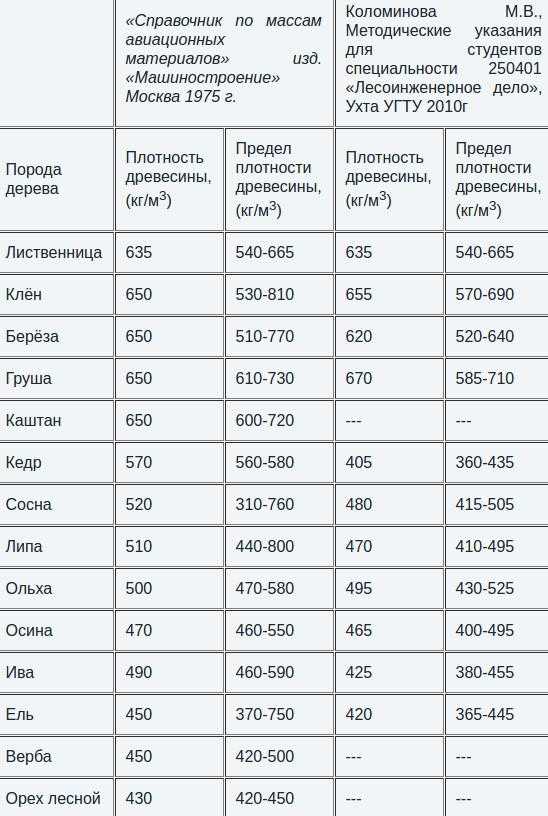 [ …]
[ …]
В накоплении радионуклидов разными группами растений отмечены видовые и другие таксономические различия. Коэффициент перехода радионуклидов из почвы в растения увеличивается в ряду: лесные ягоды — грибы — мхи и лишайники. По уровню содержания радионуклидов в древесине при одинаковой плотности загрязнения почвы и в одинаковых лесорастительных условиях основные лесообразующие породы составляют в порядке убывания следующий условный ряд: мягколиственные породы, твердолиственные, хвойные породы. Накопление радионуклидов древесным ярусом происходит интенсивнее в молод-няках, чем в средневозрастных, приспевающих и спелых дре-востоях, а деревья лучшего класса роста накапливают 137Св больше и интенсивнее, чем угнетенные и отстающие в росте. На влажных и переувлажненных почвах этот процесс происходит гораздо интенсивнее по сравнению с автоморфными условиями местопроизрастания. Наблюдается также обратная связь между трофностью почвы и интенсивностью поступления из нее радионуклидов в лесную растительность. [ …]
[ …]
Внешний вид годичных колец зависит от плоскости, в которой их рассматривают (рис. 2). У торца бревна они имеют вид концентрических окружностей, расположенных вокруг сердцевины. В течение года вырастает только один слой, называемый годичным слоем. Рисунки на поверхности досок, получаемые от годичных колец, зависят от плоскостей, на которых они видны. На радиальной поверхности древесины годичные кольца имеют вид параллельных линий (рис. 2, б). На тангенциальной поверхности (в плоскости, касательной к годичным слоям), как правило, видны концентрические и параболические кривые. Это объясняется природой вторичного утолщения у деревьев, в связи с чем плоскость разреза пересекает несколько годичных колец. Каждое отдельное кольцо прироста не в одинаковой степени проявляется в древесине, потому что интенсивность роста, а следовательно, и плотность формирующейся древесины, неодинаковы на протяжении всего вегетационного периода. Обычно наиболее быстрое увеличение в толщину происходит в начале вегетационного сезона и заметно замедляется к его концу.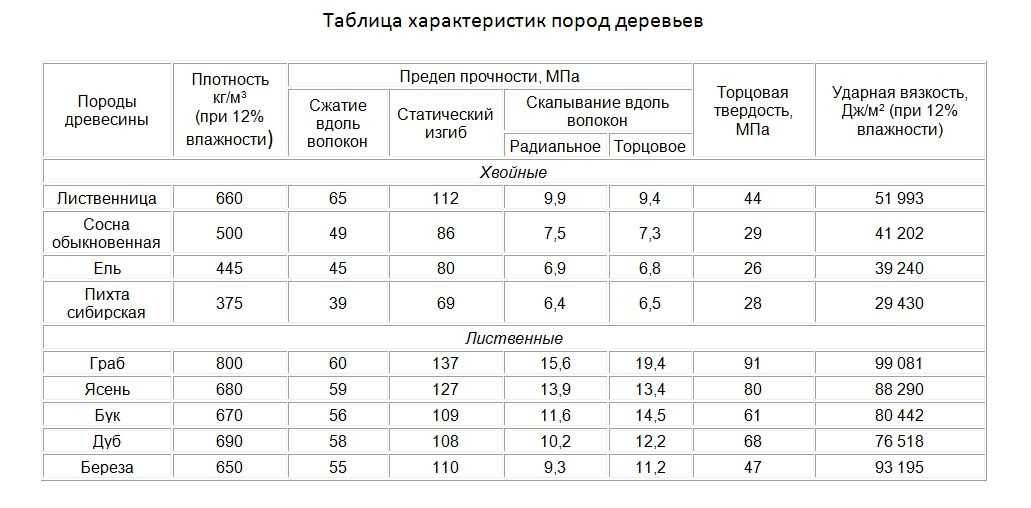 Та часть кольца, которая образуется весной, когда рост возобновляется, предназначена прежде всего для продвижения сока, поэтому она более пориста и часто обладает малой плотностью. Эта ткань носит название ранней, или весенней, древесины. Древесина, образуемая во второй половине вегетационного периода, называется поздней, или летней, древесиной. Она обычно плотнее и темнее ранней. Это можно заметить, рассматривая ее при небольшом увеличении или даже невооруженным глазом. Летняя древесина хорошо приспособлена для обеспечения прочности ствола и, по всей вероятности, не участвует в передаче сока в такой же степени, как весенняя древесина. Различие между более плотной и более темной летней древесиной данного прироста и более рыхлой и светлой весенней древесиной последующей зоны позволяет разграничивать годичные кольца.[ …]
Та часть кольца, которая образуется весной, когда рост возобновляется, предназначена прежде всего для продвижения сока, поэтому она более пориста и часто обладает малой плотностью. Эта ткань носит название ранней, или весенней, древесины. Древесина, образуемая во второй половине вегетационного периода, называется поздней, или летней, древесиной. Она обычно плотнее и темнее ранней. Это можно заметить, рассматривая ее при небольшом увеличении или даже невооруженным глазом. Летняя древесина хорошо приспособлена для обеспечения прочности ствола и, по всей вероятности, не участвует в передаче сока в такой же степени, как весенняя древесина. Различие между более плотной и более темной летней древесиной данного прироста и более рыхлой и светлой весенней древесиной последующей зоны позволяет разграничивать годичные кольца.[ …]
Объекты и методика. Исследования проводили в 1983—1986 гг. в лесах долин рек Северо-Запада европейской территории страны (ЕТС). Всего было заложено около 200 временных пробных площадей.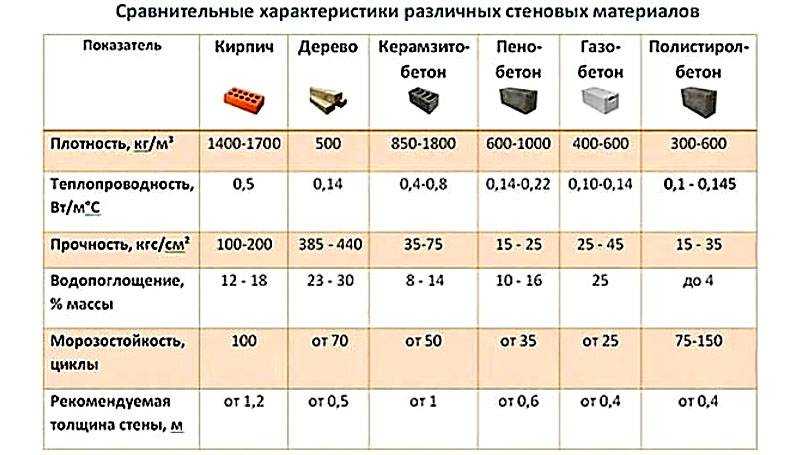 На них, кроме обычных лесотаксационных работ, измеряли освещенность под пологом нижних ярусов, делали перечет подлеска, определяли проективное покрытие доминантных видов живого напочвенного покрова, измеряли запас сухостоя и валежа, мощность подстилки и гумусового слоя, брали образцы почв и подстилки на химический анализ. В каждом фитоценозе общепринятыми методами лесной таксации и геоботаники количественно оценивали значимые синузии (ярусы) и элементы леса. По методике В. В. Загреева [12] определяли текущий прирост стволовой древесины, условную плотность и удельную теплоту сгорания древесины стволов, сухостоя, валежа, пней и подстилки.[ …]
На них, кроме обычных лесотаксационных работ, измеряли освещенность под пологом нижних ярусов, делали перечет подлеска, определяли проективное покрытие доминантных видов живого напочвенного покрова, измеряли запас сухостоя и валежа, мощность подстилки и гумусового слоя, брали образцы почв и подстилки на химический анализ. В каждом фитоценозе общепринятыми методами лесной таксации и геоботаники количественно оценивали значимые синузии (ярусы) и элементы леса. По методике В. В. Загреева [12] определяли текущий прирост стволовой древесины, условную плотность и удельную теплоту сгорания древесины стволов, сухостоя, валежа, пней и подстилки.[ …]
Лесоводственно-экологические требования следует формализовать по структурно-параметрическим изменениям лесной экосистемы, вызванным работой лесозаготовительной техники и, прежде всего, изменениям на входе в момент рубки и на выходе её в период образования типа вырубки, смыкания молодняка или на последующих этапах формирования леса. Важным элементом упомянутого методического подхода является использование выявленных связей между параметрическими показателями на входе и на выходе экосистемы. Особое значение для установления критериев рассматриваемых требований имеют входные показатели, существенно влияющие на выход экосистемы. Поэтому лесоводственно-экологические требования будут наиболее полны и надежны только тогда, когда они корректируются на выходе экосистемы на том или ином этапе формирования леса или его типа. Из-за длительности выращивания древесины после рубки и возникающими в связи с этим затруднениями в получении достоверного экспериментального материала, ограничимся лишь этапом возобновления леса или типа вырубки с учетом формирования последующих этапов леса.[ …]
Особое значение для установления критериев рассматриваемых требований имеют входные показатели, существенно влияющие на выход экосистемы. Поэтому лесоводственно-экологические требования будут наиболее полны и надежны только тогда, когда они корректируются на выходе экосистемы на том или ином этапе формирования леса или его типа. Из-за длительности выращивания древесины после рубки и возникающими в связи с этим затруднениями в получении достоверного экспериментального материала, ограничимся лишь этапом возобновления леса или типа вырубки с учетом формирования последующих этапов леса.[ …]
Плотность дерева отображения в глобальном масштабе
История изменений
09 сентября 2015 г.
Незначительные изменения были внесены в отчеты о вкладе авторов.
13 апреля 2016 г.
В раздел информации об авторе добавлена ссылка на глобальную карту плотности деревьев.
Ссылки
Pan, Y.
 et al. Большой и постоянный поглотитель углерода в лесах мира. Наука 333 , 988–993 (2011)
et al. Большой и постоянный поглотитель углерода в лесах мира. Наука 333 , 988–993 (2011)Статья ОБЪЯВЛЕНИЯ КАС Google ученый
Crowther, T.W. et al. Прогнозирование реакции биоразнообразия почвы на обезлесение: межбиомное исследование. Глоб. Изменить биол. 20 , 2983–2994 (2014)
Артикул ОБЪЯВЛЕНИЯ Google ученый
Hansen, M.C. et al. Глобальные карты изменения лесного покрова в 21 веке в высоком разрешении. Наука 342 , 850–853 (2013)
Статья ОБЪЯВЛЕНИЯ КАС Google ученый
Бонан, Г. Б. Леса и изменение климата: воздействие, обратная связь и климатические преимущества лесов. Наука 320 , 1444–1449 (2008)
Статья ОБЪЯВЛЕНИЯ КАС Google ученый
Пфайфер, М.
, Дисней, М., Куэйф, Т. и Маршан, Р. Наземные экосистемы из космоса: обзор продуктов наблюдения Земли для макроэкологических приложений. Глоб. Экол. Биогеогр. 21 , 603–624 (2012)Статья Google ученый
Туанму, М.-Н. & Jetz, W. Глобальный согласованный продукт 1-километрового земного покрова для моделирования биоразнообразия и экосистем. Глоб. Экол. Биогеогр.
23 , 1031–1045 (2014)Артикул Google ученый
Walker, A. P. et al. Прогнозирование долгосрочного связывания углерода в ответ на обогащение CO2: как и почему современные модели экосистем отличаются? Глоб. Биогеохим. Циклы 29 , 476–495 (2015)
Артикул ОБЪЯВЛЕНИЯ КАС Google ученый
Asner, G.P. et al. Универсальный бортовой LiDAR для картирования углерода в тропических лесах.
Экология 168 , 1147–1160 (2012)Статья ОБЪЯВЛЕНИЯ Google ученый
Fauset, S. et al. Гипердоминирование в круговороте углерода в амазонских лесах. Природа Комм. 6
Артикул ОБЪЯВЛЕНИЯ КАС Google ученый
Slik, J.W.F. et al. Экологические корреляции биомассы деревьев, площади основания, удельного веса древесины и градиентов плотности стволов в тропических лесах Борнео. Глоб. Экол. Биогеогр. 19 , 50–60 (2010)
Статья Google ученый
Литвик, Л. Р. и Остин, М. П. Конкурентные взаимодействия между видами деревьев в старовозрастных коренных лесах Новой Зеландии. Экология 82 , 2560–2573 (2001)
Статья Google ученый
Oliver, C.
D. & Larson, B.C. Forest Stand Dynamics (John Wiley & Sons, 1996)Google ученый
Ригинос, К. и Грейс, Дж. Б. Плотность деревьев саванны, травоядные и травянистое сообщество: восходящие и нисходящие эффекты. Экология 89 , 2228–2238 (2008)
Статья Google ученый
О’Нил-Данн, Дж., Макфаден, С. и Ройяр, А. Универсальный, ориентированный на производство подход к картографированию кроны деревьев с высоким разрешением в городских и пригородных ландшафтах с использованием GEOBIA и объединения данных. Дистанционный датчик 6 , 12837–12865 (2014)
Артикул ОБЪЯВЛЕНИЯ Google ученый
Гулдин, Р. В. Лесная наука и лесная политика в Америке: наведение мостов к устойчивому будущему. Для. Полис Экон. 5 , 329–337 (2003)
Статья Google ученый
Цао, С.
и др. Озеленение Китая естественным путем. Ambio 40 , 828–831 (2011)Артикул Google ученый
Oldfield, E. E. et al. Выращивание городского леса: производительность деревьев в ответ на биотическое и абиотическое управление земельными ресурсами. Восстановление Ecol . (http://dx.doi.org/10.1111/rec.12230) (2015)
Надкарни, Н. Между Землей и небом: наши тесные связи с деревьями (University of California Press, 2008)
Книга Google ученый
ter Steege, H. et al. Гипердоминирование во флоре деревьев Амазонки. Наука 342 , 1243092 (2013)
Статья Google ученый
Бонан Г.Б. и Шугарт Х.Х. Факторы окружающей среды и экологические процессы в бореальных лесах.
год. Преподобный Экол. Сист. 20 , 1–28 (1989)Статья Google ученый
Мейфройдт П. и Ламбин Э. Ф. Глобальный переход к лесному хозяйству: перспективы прекращения обезлесения. год. Преподобный Окружающая среда. Ресурс. 36 , 343–371 (2011)
Статья Google ученый
Рудель, Т. К. Национальные детерминанты обезлесения в странах Африки к югу от Сахары. Фил. Транс. Р. Соц. Лонд. Б 368 , 20120405 (2013)
Артикул Google ученый
Hengeveld, G.M. et al. Карта управления лесами европейских лесов. Экол. соц. 17 , 53 (2012)
Артикул Google ученый
Киндерманн, Г. Э., МакКаллум, И., Фриц, С. и Оберштайнер, М.
Глобальная карта запасов лесов, биомассы и углерода на основе статистики ФАО. Silva Fennica 42 , 387–396 (2008)Статья Google ученый
Stephenson, N.L. et al. Скорость накопления углерода деревьями постоянно увеличивается с размером дерева. Природа 507 , 90–93 (2014)
Статья ОБЪЯВЛЕНИЯ КАС Google ученый
Чжу, К., Вудалл, К.В., Гош, С., Гельфанд, А.Е. и Кларк, Дж.С. Двойное воздействие изменения климата: миграция и оборот лесов на протяжении всей жизни. Глоб. Изменить биол. 20 , 251–264 (2014)
Статья ОБЪЯВЛЕНИЯ Google ученый
Льюис, С. Л. и др. Надземная биомасса и структура 260 африканских тропических лесов. Фил. Транс. Р. Соц. Лонд. В 368 , 20120295 (2013)
Артикул Google ученый
Брус, Д.
Дж. и др. Статистическое картирование видов деревьев в Европе. евро. Дж. Для. Рез. 131 , 145–157 (2011)Статья Google ученый
Лесная служба Министерства сельского хозяйства США. Национальная программа инвентаризации и анализа лесов http://fia.fs.fed.us/ (2010)
Стейнванд, Р. С., Хатчинсон, Дж. А. и Снайдер, Дж. П. Картографические проекции для глобальных и континентальных наборов данных и анализ искажения пикселей, вызванного перепроецированием. Фотограмм. англ. Дистанционное зондирование 61 , 1487–1499 (1995)
Google ученый
Chavent, M., Kuentz, V., Liquet, B. & Saracco, J. ClustOfVar: пакет R для кластеризации переменных. J. Стат. ПО 50 , 1–16, http://www.jstatsoft.org/v50/i13/ (2012)
Статья Google ученый
Bartoń, K.
MuMIN: Выбор модели и усреднение модели на основе информационных критериев (AICc и т.п.) . (https://cran.r-project.org/web/packages/MuMIn/index.html) (2015)Google ученый
MacKenzie, D. I. et al. Оценка занятости и моделирование (Академическая пресса, 2005)
Google ученый
MacLean, M.G. et al. Требования к маркировке лесных полигонов в классификации анализа изображений на основе объектов. Междунар. J. Remote Sens. 34 , 2531–2547 (2013)
Статья ОБЪЯВЛЕНИЯ Google ученый
Ståhl, G. et al. Вывод на основе модели для оценки биомассы в выборочной съемке LiDAR в округе Хедмарк, Норвегия. Кан. Дж. Для. Рез. 41 , 96–107 (2011)
Статья Google ученый
Туанму, М.
-Н. и Джец, В. Глобальная характеристика неоднородности наземной среды обитания на основе дистанционного зондирования для моделирования биоразнообразия и экосистем. Глоб. Экол. Биогеогр . http://dx.doi.org/10.1111/geb.12365 (2015)
 et al. Большой и постоянный поглотитель углерода в лесах мира. Наука 333 , 988–993 (2011)
et al. Большой и постоянный поглотитель углерода в лесах мира. Наука 333 , 988–993 (2011)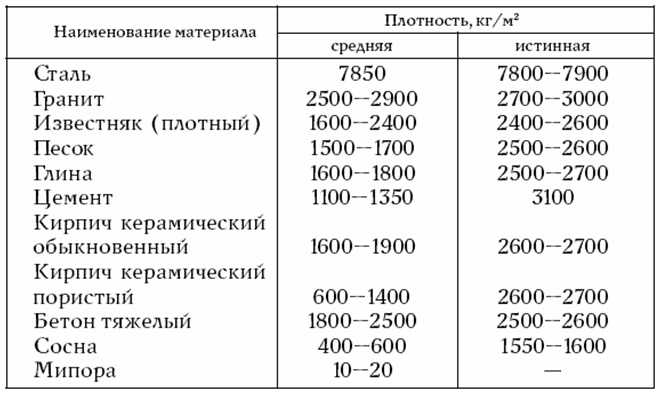 , Дисней, М., Куэйф, Т. и Маршан, Р. Наземные экосистемы из космоса: обзор продуктов наблюдения Земли для макроэкологических приложений. Глоб. Экол. Биогеогр. 21 , 603–624 (2012)
, Дисней, М., Куэйф, Т. и Маршан, Р. Наземные экосистемы из космоса: обзор продуктов наблюдения Земли для макроэкологических приложений. Глоб. Экол. Биогеогр. 21 , 603–624 (2012)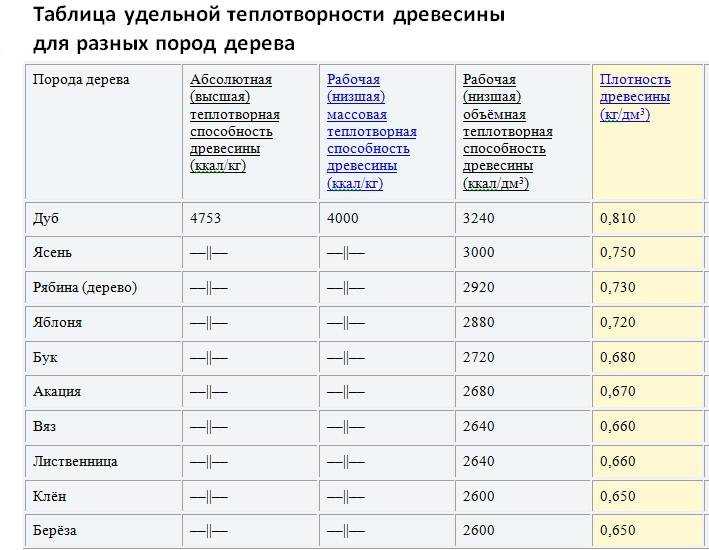 Экология 168 , 1147–1160 (2012)
Экология 168 , 1147–1160 (2012)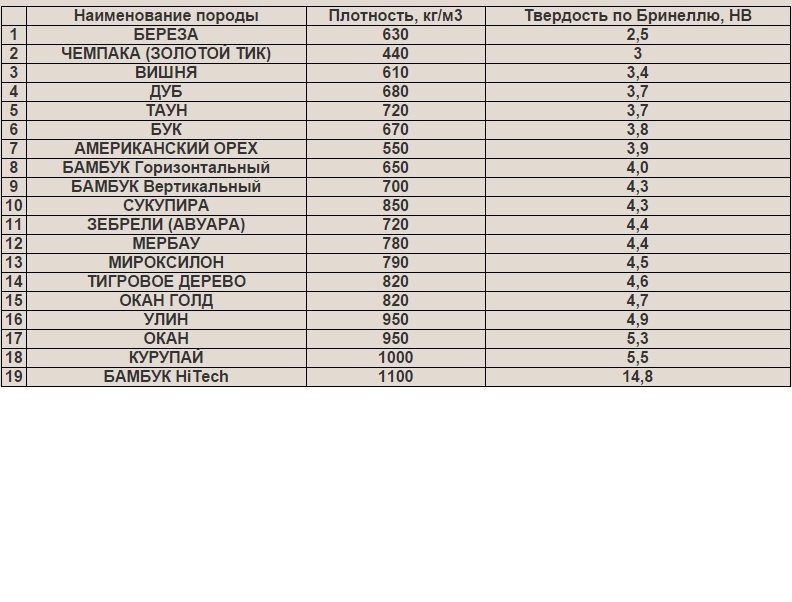 D. & Larson, B.C. Forest Stand Dynamics (John Wiley & Sons, 1996)
D. & Larson, B.C. Forest Stand Dynamics (John Wiley & Sons, 1996) и др. Озеленение Китая естественным путем. Ambio 40 , 828–831 (2011)
и др. Озеленение Китая естественным путем. Ambio 40 , 828–831 (2011) год. Преподобный Экол. Сист. 20 , 1–28 (1989)
год. Преподобный Экол. Сист. 20 , 1–28 (1989)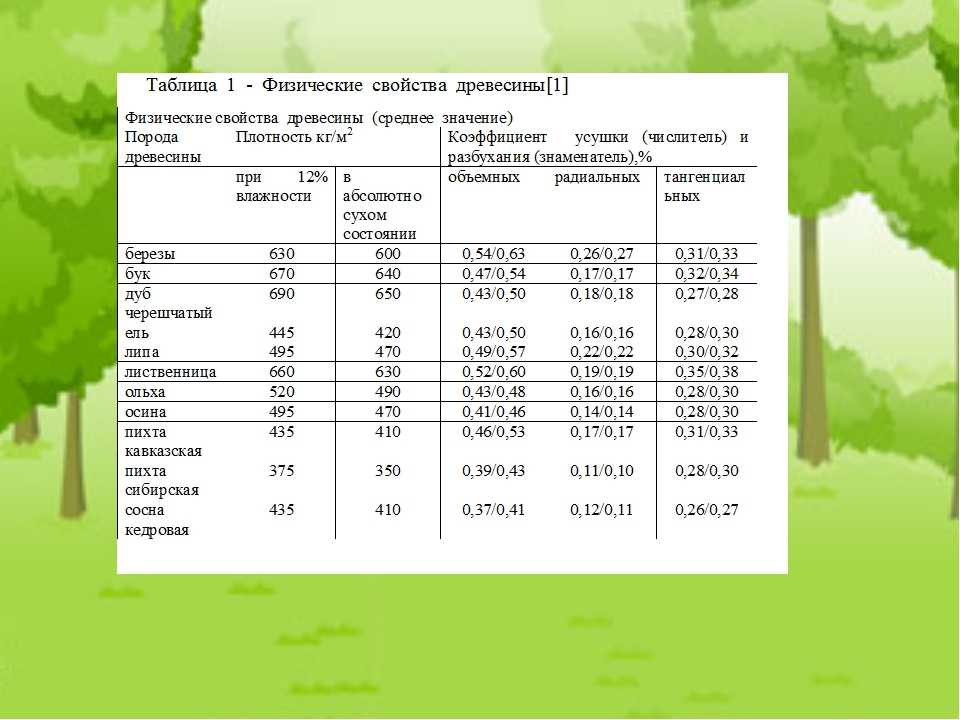 Глобальная карта запасов лесов, биомассы и углерода на основе статистики ФАО. Silva Fennica 42 , 387–396 (2008)
Глобальная карта запасов лесов, биомассы и углерода на основе статистики ФАО. Silva Fennica 42 , 387–396 (2008) Дж. и др. Статистическое картирование видов деревьев в Европе. евро. Дж. Для. Рез. 131 , 145–157 (2011)
Дж. и др. Статистическое картирование видов деревьев в Европе. евро. Дж. Для. Рез. 131 , 145–157 (2011) MuMIN: Выбор модели и усреднение модели на основе информационных критериев (AICc и т.п.) . (https://cran.r-project.org/web/packages/MuMIn/index.html) (2015)
MuMIN: Выбор модели и усреднение модели на основе информационных критериев (AICc и т.п.) . (https://cran.r-project.org/web/packages/MuMIn/index.html) (2015)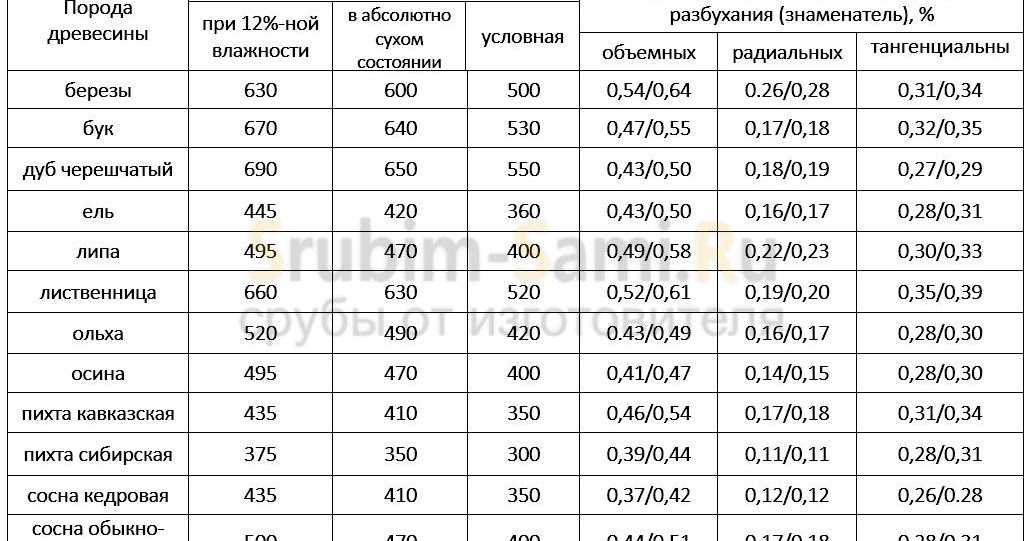 -Н. и Джец, В. Глобальная характеристика неоднородности наземной среды обитания на основе дистанционного зондирования для моделирования биоразнообразия и экосистем. Глоб. Экол. Биогеогр . http://dx.doi.org/10.1111/geb.12365 (2015)
-Н. и Джец, В. Глобальная характеристика неоднородности наземной среды обитания на основе дистанционного зондирования для моделирования биоразнообразия и экосистем. Глоб. Экол. Биогеогр . http://dx.doi.org/10.1111/geb.12365 (2015)Ссылки на скачивание
Благодарности
Мы благодарим П. Петеркинс за ее поддержку на протяжении всего исследования. Мы также благодарим Plant for the Planet за первоначальные обсуждения и сотрудничество во время исследования. Основной проект финансировался за счет грантов T.W.C. от Йельского института климата и энергетики и Британского экологического общества. Мы признаем различные источники для измерений и оценок плотности деревьев: Национальная инвентаризация лесов Канады (https://nfi.nfis.org/index.php), Лесная служба Министерства сельского хозяйства США за их Национальную инвентаризацию и анализ лесов (http:/ /fia.fs.fed.us/), Тайваньское бюро лесного хозяйства (предоставившее Национальную базу данных растительности Тайваня), DFG (Немецкий исследовательский фонд), BMBF (Федеральное министерство образования и науки Германии), Флористическое и лесное Инвентаризация Санта-Катарины (IFFSC), Национальная база данных растительности Южной Африки и чилийские исследовательские гранты FONDECYT №. 1151495. Для Европы данные о участках NFI были собраны вместе с вкладом Ж. Рондо и М. Ватеринкса, Бельгия, Т. Белуара, Франция, Х. Полли, Германия, В. Даамен и Х. Шондерворд, Нидерланды, С. Томтер, Норвегия. , Х. Вильянуэва и А. Трасобарес, Испания, Г. Кемпе, Швеция. Данные о участках природных лесов Новой Зеландии были собраны в рамках программы LUCAS для Министерства окружающей среды (Новая Зеландия) и получены из Национального банка данных обследования растительности (Новая Зеландия) (http://nvs.landcareresearch.co.nz). Мы также выражаем признательность за исследовательский проект BCI по динамике леса, который финансировался за счет грантов Национального научного фонда С. П. Хаббелла, поддержки Центра изучения тропических лесов, Смитсоновского института тропических исследований, Фонда Джона Д. и Кэтрин Т. Макартуров, Меллона. фонда, фонда Small World Institute Fund, многочисленных частных лиц, инициативы Ucross High Plains Stewardship Initiative и тяжелой работы сотен людей из 51 страны за последние два десятилетия.
1151495. Для Европы данные о участках NFI были собраны вместе с вкладом Ж. Рондо и М. Ватеринкса, Бельгия, Т. Белуара, Франция, Х. Полли, Германия, В. Даамен и Х. Шондерворд, Нидерланды, С. Томтер, Норвегия. , Х. Вильянуэва и А. Трасобарес, Испания, Г. Кемпе, Швеция. Данные о участках природных лесов Новой Зеландии были собраны в рамках программы LUCAS для Министерства окружающей среды (Новая Зеландия) и получены из Национального банка данных обследования растительности (Новая Зеландия) (http://nvs.landcareresearch.co.nz). Мы также выражаем признательность за исследовательский проект BCI по динамике леса, который финансировался за счет грантов Национального научного фонда С. П. Хаббелла, поддержки Центра изучения тропических лесов, Смитсоновского института тропических исследований, Фонда Джона Д. и Кэтрин Т. Макартуров, Меллона. фонда, фонда Small World Institute Fund, многочисленных частных лиц, инициативы Ucross High Plains Stewardship Initiative и тяжелой работы сотен людей из 51 страны за последние два десятилетия.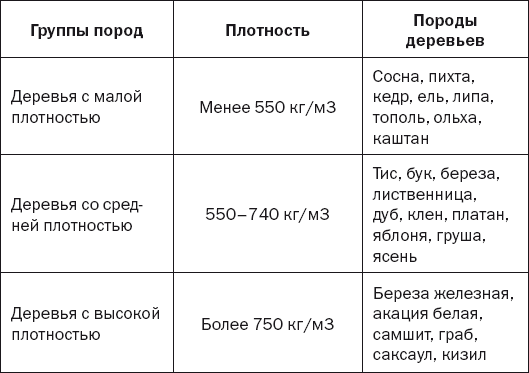 Сюжетный проект является частью Центра изучения тропических лесов, глобальной сети крупномасштабных демографических участков деревьев.
Сюжетный проект является частью Центра изучения тропических лесов, глобальной сети крупномасштабных демографических участков деревьев.
Информация об авторе
Авторы и организации
Йельская школа лесного хозяйства и экологических исследований, Йельский университет, Нью-Хейвен, 06511, Коннектикут, США
Т. В. Кроутер, Х. Б. J. R. Smith, G. Hintler, M. C. Duguid, W. Jetz, P. M. Umunay, C. W. Rowe, M. S. Ashton, P. R. Crane & M. A. Bradford
Департамент наук об окружающей среде, Университет Хельсинки, Хельсинки, 00014, Финляндия
С. М. Томас
Факультет экологии и эволюционной биологии, Йельский университет, Нью-Хейвен, 06511, Коннектикут, США
Г. Аматулли, М.-Н. Tuanmu & W. Jetz
Факультет биологических наук, Силвуд-Парк, Имперский колледж, Лондон, SL5 7PY, Великобритания
W. Jetz
Departamento de Ciencias Forestales, Universidad de La Frontera, Chile12, 08, 48
C.
SalasRedCastle Resources, Солт-Лейк-Сити, 84103, Юта, США
C. Stam
Universidade Federal do Sul da Bahia, Ferradas, 45613-204, Itabuna, Бразилия
D. Piotto
Департамент лесного хозяйства, Продовольственное сельское хозяйство и Организация Объединенных Наций, 001, Рим Италия
Р. Тавани
Operation Wallacea, Spilbsy, PE23 4EX, Lincolnshire, UK
S. Green & G. Bruce
Институт охраны природы и экологии Даррелла (DICE), Школа антропологии SAC), Кентский университет, Кентербери, ME4 4AG, Великобритания
S. Green
Центр исследований молекулярной визуализации Mircen/CEA, CNRS URA 2210, Orsay Cedex, 91401, France
S. J. Williams
Randcare Research, Lincoln, 7640, New Zealand
S. W. W.
.
WSL, Швейцарский федеральный институт исследований леса, снега и ландшафта, Бирменсдорф, 8903, Швейцария
M.
O. HuberГруппа экологических наук, Университет и исследовательский центр Вагенингена, PB, 6708, Нидерланды
Г. М. Хенгевельд и Г.-Дж. Набуурс
Центр экологии и продуктивности леса РАН, Москва, 117997, Россия
Тихонова Е.В.
P. Borchardt
Факультет ботаники и зоологии, Масариков университет, Брно, 61137, Чехия
C.-F. Ли
Южноафриканский национальный институт биоразнообразия, Исследовательский центр Кирстенбош, Клермонт, 7735, Южная Африка
L. W. Powrie
Институт наук о растениях, Ботанический сад и Центр исследований изменения климата Эшгера, Бернский университет, Берн, 3013, Швейцария
M. Fischer
Senckenberg Gesellschaft für Naturforschung, Центр исследования биоразнообразия и климата (BIK-F), Франкфурт, 60325, Германия
M. Fischer
Кафедра систематики растений, Байройтский университет, Байройт, 95447, Германия
A.
HempИнститут растениеводства им. Homeier
Исследовательская группа тропической экологии, Ланкастерский экологический центр, Ланкастерский университет, Ланкастер, LA1 4YQ, Великобритания
P. Cho
Departamento de Engenharia Florestal, Universidade Regional de Blumenau/Blumenanta 8, Cat030-000, Бразилия
A. C. Vibrans
Сино-Франч Институт Земли Системной Науки, Колледж городских и экологических наук, Пекинский университет, Пекин, 100871, Китай
S. L. Piao
 Salas
Salas O. Huber
O. Huber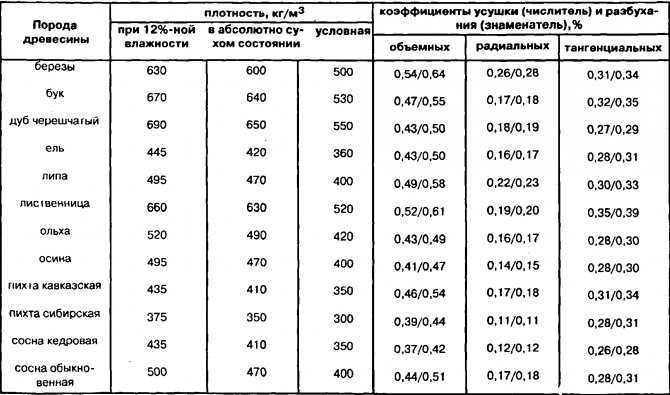 Hemp
HempАвторы
- 9299599
Автор
- 9299999
Автор
- 9
AUTHOR Crowther
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
 Р. Кови
Р. КовиПосмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
 Amatulli
AmatulliПросмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
 Green
GreenПросмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar

Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
 Cho
ChoПросмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Академия
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
 A. Bradford
A. BradfordПосмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Взносы
Исследование было задумано T.W.C и G.H. и разработан T.W.C., K.R.C. и М.А.Б. Статистический анализ проводили с помощью H.B.G., S.M.T., J.R.S., C.B., D.S.M. и T.W.C. и картирование было проведено H.B.G. и C.B. Рукопись была написана T.W.C. при участии M.A.B., PC, D.S.M., H.B.G. и C.B., с комментариями всех остальных авторов. Измерения плотности деревьев или геопространственные данные со всего мира были предоставлены K.R.C., S.M.T., M.C.D., G.A., M.N.T., WJ, C.Sa., C.St., DP, TT, S.G., G.B., S.J.W., S.K.W., M.O.H., Г.М.Х., Г.Дж.Н., Э.Т., П.Б., К.Ф.Л., Л.В.П., М.Ф., А.Х., Дж.Х., ПК, А.К.В., П.М.У., С.Л.П., К.В.Р. и М.С.А.
Автор, ответственный за переписку
Т. В. Кроутер.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют о конкурирующих финансовых интересах.
Дополнительная информация
Глобальную карту плотности деревьев можно найти по адресу http://elischolar.library.yale.edu/yale_fes_data/1/.
Расширенные данные и таблицы
Расширенные данные Рисунок 1 Гистограмма собранных измерений плотности лесных деревьев в каждом биоме по всему миру (
n = 429 775).Красная линия и синие пунктирные линии обозначают среднее значение и медиану собранных данных соответственно. Данные в каждом биоме соответствовали отрицательной биномиальной структуре ошибки.
Расширенные данные Рисунок 2. Гистограмма прогнозируемых значений плотности лесных деревьев для мест, где были собраны измерения плотности в каждом биоме по всему миру (
n = 429 775). Красная линия и синие пунктирные линии обозначают среднее значение и медиану собранных данных соответственно. Поскольку наши модели были основаны на средних значениях, большинство точек попадают или близки к средним значениям в каждом биоме.
Расширенные данные Рисунок 3. Гистограмма общих прогнозируемых значений плотности лесных деревьев для каждого пикселя в каждом биоме по всему миру (
n = 429 775).Это иллюстрирует распределение пикселей по каждому биому и подчеркивает, что наша карта учитывает систематическую ошибку выборки на графиках плотности деревьев (например, хотя у нас не было нулевых значений на наших графиках пустыни, подавляющее большинство пикселей пустыни не содержат деревьев). ).
Расширенные данные Рис. 4. Сравнение подходов к созданию глобальной карты плотности деревьев.
Первоначальная карта была создана с использованием 14 моделей на уровне биомов (биомов, описанных The Nature Conservancy http://www.nature.org) для учета крупномасштабных изменений типов наземной растительности. Благодаря нескольким тысячам измерений плотности на уровне участка в большинстве биомов этот подход обеспечил высокоточные оценки в глобальном масштабе. Однако для повышения точности в локальном масштабе мы также создали карту с использованием моделей в масштабе экорегиона.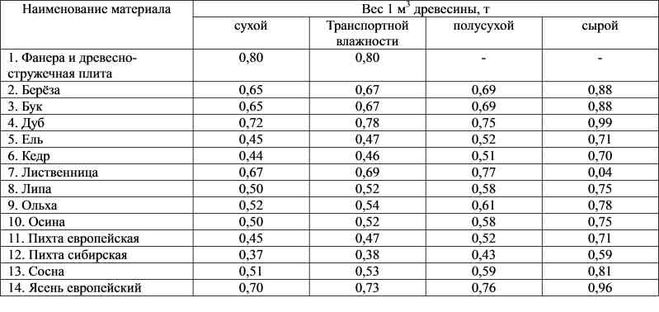 Отдельные модели были созданы в каждом из 813 глобальных экорегионов (также выделенных The Nature Conservancy для отражения типов растительности меньшего масштаба) с использованием точно такого же статистического подхода (см. Методы). Тот же 429Для построения каждой карты использовалось 775 точек данных. Карты уровня биома и уровня экорегиона дают общую оценку деревьев в 3,041 и 3,253 триллиона деревьев соответственно.
Отдельные модели были созданы в каждом из 813 глобальных экорегионов (также выделенных The Nature Conservancy для отражения типов растительности меньшего масштаба) с использованием точно такого же статистического подхода (см. Методы). Тот же 429Для построения каждой карты использовалось 775 точек данных. Карты уровня биома и уровня экорегиона дают общую оценку деревьев в 3,041 и 3,253 триллиона деревьев соответственно.
Полноразмерная таблица
Дополнительная информация
Дополнительная таблица 1
Сводная таблица, показывающая количество оценок участков и общее количество деревьев (с 95% доверительный интервал) в биоме и глобальном масштабе. (XLSX, 15 КБ)
Дополнительная таблица 2
В этой таблице показано количество деревьев и плотность деревьев для стран мира, рассчитанные с использованием двух независимых подходов (модели на уровне биома и экорегиона) и базы данных глобальных административных районов, версия 2. 7 (http://gadm.org/). (XLSX 53 КБ)
7 (http://gadm.org/). (XLSX 53 КБ)
Слайды PowerPoint
Слайд PowerPoint к рис. 1
Слайд PowerPoint к рис. 2
PowerPoint Slide для рис. 3
PowerPoint Slide для рис. 4
PowerPoint Slide для рис. 5
Права и добычи
Перепечатки и разрешения
ОБЛАДА. Можешь ли ты получить?
Билл Кук, Расширение Мичиганского государственного университета —
Плотность леса является важным показателем, который сильно влияет на здоровье леса, состав растений, структуру насаждений и экологическую функцию. Плотность леса легко измерить, и ею можно управлять.
Насаждения сосны красной, где кроны слишком малы для восстановления насаждения. Во многих лесонасаждениях количество света является наиболее ограничивающим экологическим фактором, особенно для регенерации деревьев и других растений. Полог леса определяет, сколько света достигает каждого дерева и лесной подстилки. Характеристики навеса часто сильно различаются. Управление лесным хозяйством манипулирует характеристиками полога и проникновением света, среди прочего, для достижения конкретных целей. Вот несколько советов от Университета штата Мичиган о том, как управлять густотой леса.
Полог леса определяет, сколько света достигает каждого дерева и лесной подстилки. Характеристики навеса часто сильно различаются. Управление лесным хозяйством манипулирует характеристиками полога и проникновением света, среди прочего, для достижения конкретных целей. Вот несколько советов от Университета штата Мичиган о том, как управлять густотой леса.
Свет необходим для запуска процесса фотосинтеза. Фотосинтез производит сахар, который хранит химическую энергию, преобразованную из энергии света. Энергия расходуется на дыхание, чтобы растения и животные могли жить, расти и размножаться. Эти химические реакции питают жизнь, какой мы ее знаем.
Все растения, включая деревья, имеют разные требования к свету. Некоторые виды предпочитают полный солнечный свет. Другие виды лучше чувствуют себя в полутени. Стратегии получения света сильно различаются. В следующий раз, когда вы окажетесь в лесу, найдите время, чтобы увидеть, какие виды деревьев восстанавливаются, если таковые имеются. Кроме того, обратите внимание на свет, падающий на лесную подстилку, а затем следите за некоторыми из этих мест в течение года, чтобы увидеть, что происходит.
Кроме того, обратите внимание на свет, падающий на лесную подстилку, а затем следите за некоторыми из этих мест в течение года, чтобы увидеть, что происходит.
Весенняя флора проводит большую часть своего жизненного цикла при полном солнечном свете между зимней оттепелью и распусканием листвы среди деревьев. После распускания листьев низкорослые растения должны адаптироваться к тому, что осталось, впасть в спячку на годы или вымереть. Эта длительная борьба за свет в лесу влияет на ключевые экологические процессы, такие как сукцессия и нарушение.
Годовые кольца деревьев дают представление об условиях освещения в прошлом. Широкие кольца предполагают больше света и больший рост. Сужающиеся кольца показывают меньший рост и, возможно, меньше света. Замедление роста вызывает стресс у деревьев и повышенную уязвимость к насекомым и патогенам.
Лесоводы выражают запасы леса с помощью меры, называемой «базовая площадь» (BA). По определению, BA — это площадь поперечного сечения всех деревьев на высоте 4,5 фута от земли, часто деревьев диаметром не менее пяти дюймов на данном акре.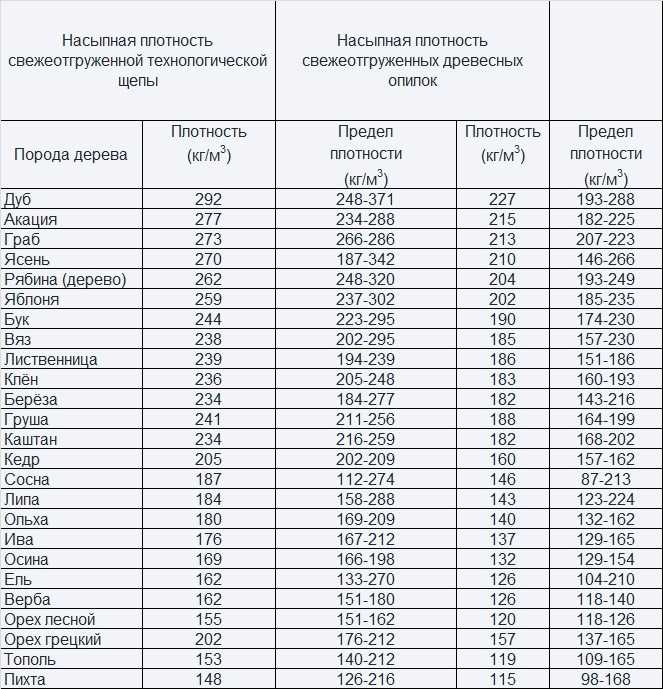 BA измеряется в количестве квадратных футов на акр (конечно, есть метрические эквиваленты). Оптимальная БА для леса будет зависеть от типа леса, но обычно варьируется от 70 до 150 квадратных футов на акр для типов леса в верховьях Великих озер.
BA измеряется в количестве квадратных футов на акр (конечно, есть метрические эквиваленты). Оптимальная БА для леса будет зависеть от типа леса, но обычно варьируется от 70 до 150 квадратных футов на акр для типов леса в верховьях Великих озер.
Поскольку леса могут быть довольно сложными, существуют разные рекомендации по плотности насаждения для различных условий, таких как состав пород, возраст, средний диаметр и т. д. Северные лиственные породы (сахарный клен, липа, желтая береза, болиголов) требуют самого высокого уровня навыков. и опыт управления. Северные лиственные леса являются наиболее распространенными типами лесов в северных лесах, и с каждым десятилетием их становится все больше.
В качестве альтернативы осина сравнительно проста. На самом деле виды осины восстанавливаются после «катастрофических нарушений», таких как пожар, продувка или сплошная рубка. Осины довольно нетерпимы к тени. Редко бывает экономически эффективным прореживание сортов осины. Жизненные стратегии осины и северных лиственных пород полярны.
Красная сосна обладает еще одним набором свойств, реагирующих на свет. Будь то естественная регенерация или посадка, красная сосна в какой-то момент «закрывает полог». Рост начинает замедляться. Нижние ветки отмирают и со временем опадают. Как только деревья приближаются к своему генетическому максимуму высоты, усыхание кроны представляет собой проблему. В то время как здоровые насаждения красной сосны хорошо реагируют на прореживание, хитрость заключается в том, чтобы проредить эти насаждения до того, как размер кроны уменьшится слишком сильно.
Если крона среднего дерева занимает менее 10-15 процентов высоты дерева, то эти деревья не смогут вырастить большие кроны. Прореживание деревьев с маленькой кроной не приведет к росту новых ветвей ниже существующей кроны, потому что красная сосна не имеет такой возможности, в отличие от клена или дуба. Прореживание может просто привести к большему разрыву ветром.
Насаждения северного белого кедра могут достигать удивительно высокой плотности, превышающей 300 квадратных футов на акр. Саженец кедра в этих очень тенистых условиях может едва выжить. Тем не менее, если его выпустить на свет, этот саженец начнет агрессивно расти. Кедр относится к меньшинству пород деревьев с такой эластичностью света.
Слишком густой сосновый древостой, где рост почти прекратился и деревья находятся в состоянии стресса.
Деревья, хотя и преобладают, не являются единственными растениями в лесу. Другие растения должны быть приспособлены к световым условиям, которые обеспечивает полог леса. Под взрослым осинником света больше, чем под взрослым кедром. Подземные растения будут соответственно различаться.
По мере появления открытых пространств, будь то естественным образом или благодаря руководству, подполье реагирует предсказуемо. Сочетание наземных растений, кустарников, молодых деревьев и взрослых деревьев называется структурой насаждения. Структурой насаждения, помимо других характеристик, можно управлять, контролируя уровень освещенности с помощью управления лесным хозяйством.
Одним из немногих недостатков управления светом является потенциальное вторжение экзотических видов растений. Кустарники, такие как крушина, осенняя маслина, японский барбарис и жимолость, занимают подлесок быстрее, чем большинство местных видов. Некоторым экзотическим наземным растениям, таким как чесночная горчица, может даже не понадобиться много света, но они постепенно займут тенистую лесную подстилку. Эти экзотические захватчики сокращают естественное разнообразие, подрывают качество среды обитания, ухудшают экологические услуги и значительно изменяют ход естественной сукцессии. Местные растения труднее выращивать.
Освобождение лесов редко приводит к положительным результатам. Почти все наши леса сильно пострадали от прошлого. В некотором смысле они не являются «естественными». Неспособность управлять уровнем освещенности и другими экологическими параметрами неизбежно приводит к серьезному снижению жизнеспособности и устойчивости леса.
Защита леса, восстановление и предоставление продуктов и услуг поставлены под угрозу из-за доброкачественного пренебрежения.