Древесина и ее применение в народном хозяйстве. Строение дерева
В народном хозяйстве древесина применяется очень широко: в промышленном и гражданском строительстве ее используют для изготовления отдельных конструктивных элементов зданий и сооружений, а также окон, дверей, паркета и т. п. Древесина — сырье для изготовления целлюлозы, бумаги, картона, этилового спирта. При химической переработке древесины получают канифоль и скипидар; из коры дуба, ивы и лиственницы добывают дубильные вещества, употребляемые в кожевенной промышленности.
В судостроении древесину применяют при закладке судов на стапеле и спуске их на воду (кильблоки, спусковые дорожки стапеля, полозья, подбрюшины), для покрытия металлических палуб, для изоляции и оборудования провизионных кладовых, трюмов, кают и других помещений, для изготовления деревянной мебели, возведения строительных лесов, временных трапов.
Широкое применение древесины объясняется ее техническими свойствами. При относительно небольшой массе она обладает достаточной прочностью, легко обрабатывается, хорошо склеивается, а изделия из нее отличаются хорошими декоративными качествами.
Вместе с тем древесина имеет и ряд отрицательных свойств. При изменении влажности она разбухает, усыхает, растрескивается и коробится. Естественные древесные материалы подвержены загниванию и легко воспламеняются. В меньшей степени эти недостатки свойственны прессованной древесине, древесностружечным и древесноволокнистым плитам.
Растущее дерево состоит из корней, ствола и кроны. Корни всасывают из почвы влагу, питательные минеральные вещества и удерживают дерево в почве. Ветви и листья дерева образуют крону. В листьях происходит так называемый процесс фотосинтеза. Они поглощают из воздушной среды углекислый газ, который разлагается на углерод и кислород. Кислород из листьев выделяется в воздух, а углерод под влиянием солнечного света, взаимодействия с влагой и растворенными в ней минеральными веществами образует различные органические соединения, необходимые для питания дерева, которые распространяются по внутреннему слою его коры.
Ствол поддерживает крону дерева, хранит влагу, а также запасы питательных веществ, вырабатываемых листьями. Он состоит из сердцевины, древесины, камбия и коры (рис. 1.1).
Он состоит из сердцевины, древесины, камбия и коры (рис. 1.1).
Рис. 1.1. Строение ствола
1 — кора; 2 — камбий; 3 — древесина; 4 — сердцевина
Сердцевина, расположенная в центре ствола, представляет собой рыхлую мягкую ткань и занимает незначительную часть ствола. Далее располагается древесина, занимающая большую часть ствола.
Камбий охватывает древесину сплошным кольцом. Он состоит из клеток, способных к делению и росту.
Кора защищает дерево от механических повреждений и от неблагоприятных влияний окружающей среды. Внутренний слой коры (лубяной) служит для отложения запасов питательных
Древесина — основной технический материал, свойства которого неразрывно связаны с ее строением. Макроструктура (элементы строения) древесины хорошо видна на разрезах ствола невооруженным глазом. Так, на разрезах четко выделяются годичные слои, сердцевинные лучи, ядро и заболонь.
Так, на разрезах четко выделяются годичные слои, сердцевинные лучи, ядро и заболонь.
Годичные слои — концентрические круги вокруг сердцевины — хорошо видны на торцовом срезе. На радиальном срезе они просматриваются в виде продольных линий, на тангенциальном — в виде дугообразных кривых. У живого дерева каждый год вырастает один годичный слой. Рост происходит от камбиального слоя, поэтому чем ближе расположен слой к сердцевине, тем он старее. По числу годичных слоев на торцовом срезе комлевой части ствола можно определить возраст дерева. Годичный слой состоит из ранней древесины, образованной в весенний период роста, и поздней, развивавшейся в летний период. Окраска ранней древесины светлая. Поздняя древесина имеет более темную окраску и отличается большой плотностью и высокими механическими качествами.
Сердцевинные лучи сохраняют, а также проводят влагу и питательные вещества в горизонтальном направлении между годичными слоями. На торцовом срезе ствола (особенно у дуба, бука и клена) сердцевинные лучи имеют вид радиально расположенных узких полосок, на радиальном срезе — светлых или темных блестящих полосок, а на тангенциальном — темных штрихов с заостренными концами, расположенных по длине ствола.
У деревьев некоторых пород окраска периферийной и центральной частей ствола на торцовом и радиальном срезах различна. Такие породы деревьев называют ядровыми. Породы деревьев, стволы которых на срезе не имеют этого различия окраски, называют заболонными.
Центральная, темноокрашенная часть древесины, прилегающая к сердцевине, является ядром. Оно состоит из омертвевшей древесины, которая отличается повышенной стойкостью к загниванию, так как содержит значительное количество смол и дубильных веществ.
Заболонь — часть древесины раннего образования, состоящая из молодых клеток. В растущем дереве по заболони движется влага с растворенными в ней минеральными веществами. Ширина заболони зависит от возраста и места произрастания дерева. Широкую заболонь имеют сосна и дуб. По механическим свойствам заболонная древесина не отличается от древесины ядра, но менее устойчива к загниванию, так как легко впитывает влагу.
Микроструктура древесины различима только под микроскопом.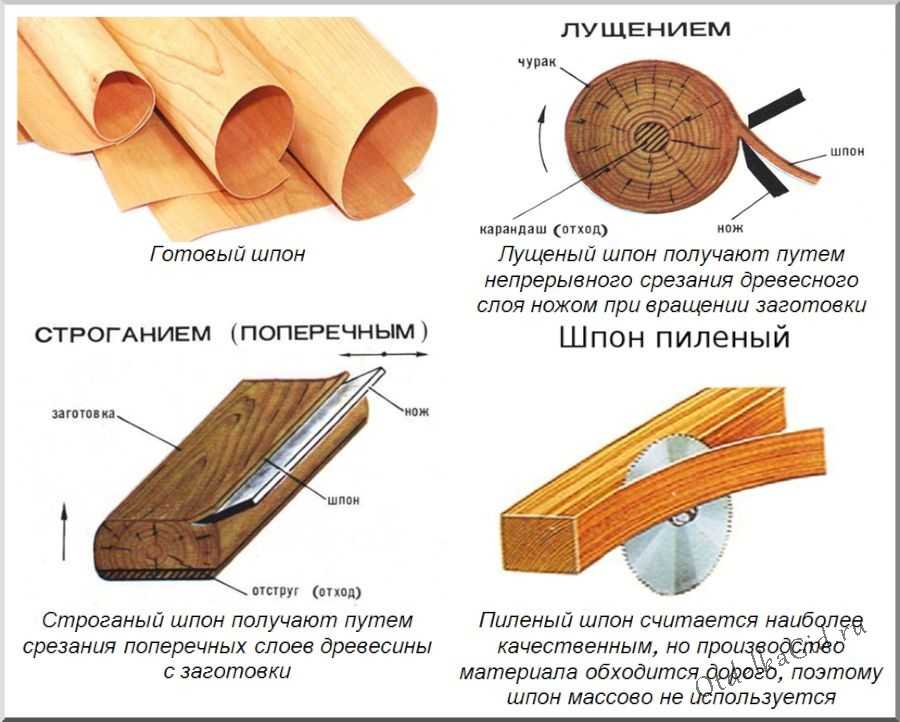 Основными элементами древесины являются клетки, древесные волокна, сосуды и трахеиды.
Основными элементами древесины являются клетки, древесные волокна, сосуды и трахеиды.
Клетки представляют собой тонкие замкнутые оболочки, внутри которых находятся протоплазма и ядро. Клетки размножаются путем деления: ядро делится на две части и из одной клетки образуются две малые. По мере роста каждая из них делится также на две и т. д. Живые клетки служат для передачи питательных веществ друг другу и хранения запасов этих веществ на зимний период.
Древесные волокна — длинные одеревеневшие толстостенные клетки, составляющие основную часть древесины лиственных пород.
Сосуды — вытянутые тонкостенные клетки, через которые влага от корней проходит к листьям. Сосуды встречаются только в древесине лиственных пород.
Трахеиды — длинные замкнутые клетки, расположенные радиальными рядами. Они составляют основную часть древесины хвойных пород. Тонкостенные трахеиды заменяют собой сосуды, а толстостенные — волокна клетки. В древесине многих хвойных пород расположены смоляные ходы, в которых вырабатывается смола.
- Породы дерева
- Физические, механические и декоративные свойства древесины
- Пороки древесины
- Виды древесных материалов
- Пластмассы и синтетические материалы
Использование древесины в строительстве
Как стройматериал, древесина имеет свои очень важные достоинства: высокую прочность, малую теплопроводность, небольшую плотность, легко обрабатывается. При изготовлении строительных конструкций наиболее часто используются деревья хвойных пород: ель, сосна, пихта, кедр и др. Из них изготавливаются несущие конструкции. Наиболее часто применяется сосна. Деревья лиственных пород имеют меньшую устойчивость.
Дуб имеет наибольшую ценность из лиственных пород древесины. Его используют для строительства перегородок, стен, перекрытий и покрытий.
Рис. 2. Текстура сырого дуба
Дуб имеет очень прочную древесину, он очень устойчив к загниванию, но склонен к растрескиванию. Древесина дуба используется для производства паркета, для отделки в судостроении, для изготовления особенно ценных резных элементов.
Древесина дуба используется для производства паркета, для отделки в судостроении, для изготовления особенно ценных резных элементов.
Сосна применяется для устройства стен домов, оконных переплетов, пола, дверей. А также в строительстве мостов, изготовлении шпал, строительстве эстакад, изготовления фанеры.
Рис. 3. Фактура сосны
Кедр используется для изготовления декоративной фанеры.
Рис. 4. Фактура кедра
Ясень по свойствам похож на дуб, но у него древесина светлее. Она имеет высокую прочность, но тяжело поддается ручной обработке. При окраске цвет приобретает «седину», поэтому ясень используется в натуральном виде. Применяется для производства столярных изделий и паркета. Ясень хорошо сохраняется в воде и на воздухе, но загнивает при переменной влажности.
Рис. 5. Фактура ясеня
Береза дает белую древесину с желтоватым оттенком. Она прочна, имеет высокую твердость, но при повышении влажности легко гниет. Береза используется для изготовления мебели, фанеры, для ограждения, для изготовления мелких деталей инструмента и предметов домашнего обихода.
Она прочна, имеет высокую твердость, но при повышении влажности легко гниет. Береза используется для изготовления мебели, фанеры, для ограждения, для изготовления мелких деталей инструмента и предметов домашнего обихода.
Рис. 6. Фактура березы
Древесина осины обладает высокой прочностью в сухой среде, хорошо обтачивается, колется, при высыхании мало коробится и дает небольшое количество трещин. Осину используется для устройства кровель, производства фанеры, а также для сооружения временных построек. В деревне осина считается лучшим материалом для строительства сруба колодца. В бане полки и обшивку стен также предпочитают делать из осины.
Рис. 7. Фактура осины
Древесина бука имеет белый цвет с красноватым оттенком, прекрасно гнется, не загнивает в условиях с повышенной влажностью. При сушке коробится и растрескивается. Используют бук для производства паркета и фанеры, мебели. Изделия из бука обязательно красят или лакируют, так как он легко впитывает влагу.
Рис. 8. Фактура бука
Ольха в качестве стройматериала используется, в тех же условиях, как береза. Применяют ольху в отделке. Она легко поддается обработке.
Рис. 9. Фактура ольхи
Клен дает древесину, по твердости только незначительно уступающую дубу. Она используется для производства фанеры. Древесина клена применяется при имитации других пород деревьев, так как она хорошо полируется и впитывает протравы.
Рис. 10. Фактура клена
Тополь имеет мягкую древесину. Она используется для временных сооружений и в декоративных изделиях. Из нее также производят материал для покрытия пола, который бывает нескольких видов: паркетные доски, паркет штучный, ДСП, ДВП. Для производства половых покрытий используют сосну, березу, дуб, ясень, бук, клен. Для настила деревянного пола используют доски из кедра, сосны, ели, ольхи.
Рис.11. Фактура тополя
Древесина липы мягкая, легкая, однородного строения, хорошо режется, мало трескается и мало коробится. По физико-механическим свойствам близка осине. Она применяется для изготовления чертежных досок, моделей в литейном деле, деревянной посуды, карандашей, резных изделий и игрушек. Из древесины липы изготовляют тару под жидкие продукты (липовые кадки под мед), ящики для столов, шкафов, комодов, древесную стружку (для упаковки парфюмерии). Из коры липы получают мочало, из которого изготовляют рогожи, кули, веревки и т. д.
По физико-механическим свойствам близка осине. Она применяется для изготовления чертежных досок, моделей в литейном деле, деревянной посуды, карандашей, резных изделий и игрушек. Из древесины липы изготовляют тару под жидкие продукты (липовые кадки под мед), ящики для столов, шкафов, комодов, древесную стружку (для упаковки парфюмерии). Из коры липы получают мочало, из которого изготовляют рогожи, кули, веревки и т. д.
Наряду с достоинствами, древесина имеет свои недостатки: она гниет, быстро возгорается, коробление, растрескивание, различная плотность вдоль волокон и поперек них, много отходов при обработке.
В деревянном строительстве по сложившейся практике наиболее часто применяют сосну и ель. Они имеют ровный прямой ствол, не имеют дупел, ветвей немного. Все эти особенности облегчают изготовление строительных конструкций и сбивку стены. Благодаря большому содержанию смолы хвойная древесина очень хорошо сопротивляются гниению.
В строительстве также часто применяются лиственница и кедровая сосна, используется и осина. А из ольхи, бука, дуба и липы бревенчатое строительство ведется нечасто.
А из ольхи, бука, дуба и липы бревенчатое строительство ведется нечасто.
Продолжительность жизни бревенчатого дома зависит не только от породы дерева, но и от качества исходной древесины, на которое влияют многие фактор. В первую очередь, важно место и время заготовки и методы сушки. О качестве древесины можно судить по ее плотности. В основном плотность древесины определяется местом, где проводятся заготовки и климатическими условиями района. Более высокую плотность имеет древесина северных районов России.
Считается, что самая подходящая древесина для строительства заготавливается в период с конца октября по март месяц. Сосну и ель заготавливать надо в феврале-марте, так как древесина в это время будет более сухой, это значит, при ее обработке трещин возникнет меньше. При низких температурах окружающей среды на деревьях не появляется грибок, а при последующей обработке кора деревьев легко снимается. В результате очищенные от коры бревна почти не травмируются.
Сосна относится к ядровым хвойным деревьям. Древесина заболони сосны желтоватая, шириной до 10 см, древесина ядра значительно темнее, коричневатого цвета. В поперечном разрезе сосны ясно видны годовые кольца, благодаря резко выраженной различной окраске весенней и летней древесины. Смолистость сосны очень высока (18—20 кг на кубический метр древесины), благодаря чему обеспечивается высокая стойкость древесины против загнивания, поэтому сосна применяется в частях сооружений, подверженных атмосферным воздействиям.
Древесина заболони сосны желтоватая, шириной до 10 см, древесина ядра значительно темнее, коричневатого цвета. В поперечном разрезе сосны ясно видны годовые кольца, благодаря резко выраженной различной окраске весенней и летней древесины. Смолистость сосны очень высока (18—20 кг на кубический метр древесины), благодаря чему обеспечивается высокая стойкость древесины против загнивания, поэтому сосна применяется в частях сооружений, подверженных атмосферным воздействиям.
В северном климате наилучшее качество имеет сосна в возрасте от 80 до 140 лет. Ее ствол обладает малой сбежистостью, то есть мало меняется по длине, имеет большую высоту — до 40 м, ветви дерева растут под углом к стволу в мутовку, а в густых насаждениях ствол почти лишен ветвей. Вес воздушно-сухой сосны колеблется от 400 до 700 кг на куб м. Следует различать сосну, выросшую на низких, сырых почвах — мяндовую и на сухих песчаных почвах — рудовую. Рудовая сосна имеет более темную окраску, меньшую ширину заболони, большую плотность, поэтому ценится как строительный материал выше.
Ель также широко применяется в строительстве. Она относится к заболонным деревьям; ее ядро по цвету не отличается от заболони. Древесина ели более легкая и мягкая, имеет более светлый цвет, годовые слои не так резко выделяются. Ее смолистость значительно меньше, чем у сосны, поэтому еловую древесину не рекомендуется применять в незащищенных сооружениях. Крепость ели мало отличается от сосны, но она менее стойкая. Ствол ели прямой, ровный, ветви растут под прямым углом в мутовку. Нижние ветви, отмирая, не отпадают, а остаются в виде сучьев в стволе. У ели упругость больше, чем у сосны.
Пихта близка по качеству своей древесины к ели, но немного уступает ей в прочности, более легко колется, не содержит смолы, цвет древесины более серый, ядра не имеется. Применение в незащищенных сооружениях, ввиду малой стойкости, не рекомендуется. Ствол очень полный, ветви растут под прямым углом к стволу и, так же как и у ели не отпадают.
Лиственница считается наиболее ценным по своей крепости и стойкости материалом. По плотности, удельному весу, смолистости и крепости она превосходит сосну. Древесина имеет красноватый цвет, с узкой заболонью и резко очерченным ядром. Недостаток лиственницы — сильная трещиноватость при быстром высыхании. Лиственницу предпочтительно использовать в сооружениях, находящихся в условиях повышенной влажности, а также наряду с дубом — на прокладки и вкладыши там, где требуется повышенная прочность.
По плотности, удельному весу, смолистости и крепости она превосходит сосну. Древесина имеет красноватый цвет, с узкой заболонью и резко очерченным ядром. Недостаток лиственницы — сильная трещиноватость при быстром высыхании. Лиственницу предпочтительно использовать в сооружениях, находящихся в условиях повышенной влажности, а также наряду с дубом — на прокладки и вкладыши там, где требуется повышенная прочность.
Дуб как основной материал в массовом строительстве не употребляется, из-за его дороговизны. Благодаря большой крепости и прочности, дуб в настоящее время применяется в виде нагелей, шпонок, подушек, вкладышей, т. е. в таких частях конструкций, где обычные породы — сосна и ель — из-за незначительной их твердости и малого сопротивления скалыванию не выдержать рабочих нагрузок. Также он часто используется для изготовления деталей отделки с художественной резьбой.
В некоторых случаях дуб может заменить береза, которая обладает достаточно большой крепостью. Наиболее широкое распространение береза получила при выделке фанеры. Попытки применения березы в конструкциях показали, что в этом качестве она является недостаточно надежным материалом в ответственных сооружениях, так как она быстро загнивает.
Попытки применения березы в конструкциях показали, что в этом качестве она является недостаточно надежным материалом в ответственных сооружениях, так как она быстро загнивает.
При изготовлении мебели и предметов декоративно-прикладного искусства в основном используются те же породы древесины, как и при строительстве. Однако для отделки, конечно, применяются породы, которые обладают более высокими декоративными свойствами.
Для изготовления бижутерии рекомендуют использовать древесину следующих пород деревьев: тисс, береза, дуб, черемуха, рябина, липа, ель, яблоня, груша, осина, грецкий орех. Древесина является исходным материалов для изготовления больше тридцати тысяч изделий и продуктов.
ТИСС больше всего подходит для ювелирных работ. Его древесина имеет красивую текстуру, желтовато-красного цвета. При вымачивании приобретает фиолетовый цвет, хорошо полируется. Со временем древесина темнеет.
Рис. 12. Фактура тисса
БЕРЕЗА обладает твердой древесиной. Текстура имеет извилистое строение. В слегка подгнившей древесине березы много декоративных оранжевых, желтых и коричневых разводов. Особенно красивым рисунком отличается карельская береза.
Текстура имеет извилистое строение. В слегка подгнившей древесине березы много декоративных оранжевых, желтых и коричневых разводов. Особенно красивым рисунком отличается карельская береза.
Древесина ДУБА твердая и пористая, стойкая к гниению. Она окрашена в темно-бурые и светлые тона. Древесина дуба, пролежавшая долго в воде, приобретает темный до черного цвет и ее называют мореным дубом. Ее твердость при этом повышается. Окраску принимает плохо. Годовые кольца с разной плотностью создают игру света и тени.
ЧЕРЕМУХА. Древесина крепкая, одновременно пластичная и нежная. Хорошо полируется, окрашивается, мало коробится. Цвет древесины красно-бурый, имеет черные прожилки.
Рис. 13. Фактура черемухи
РЯБИНА дает белую древесину, иногда содержит черные и бурые слои. Окрашивается и полируется хорошо.
Рис. 14. Фактура рябины
ЛИПА имеет белая древесину, мало трескается, легко режется, мало коробится. В своем роде она является образцовым поделочным материалом.
В своем роде она является образцовым поделочным материалом.
Рис. 15. Фактура липы
ЕЛЬ. При высыхании древесина сильно трескается.
ЯБЛОНЯ. Красивей древесина у дикой яблони. У нее красно-бурый цвет со многими прожилками. Окрашивается хорошо. У культивированной яблони древесина менее ценна. Древесина яблони очень твердая. Благодаря этому раньше она была основным материалом для изготовления корпусов столярных рубанков.
Рис. 16. Фактура яблони
Рис. 17. Фактура лиственницы
ГРУША. Цвет древесины красно-бурый. Легко подается обработке и хорошо полируется.
Рис. 18. Фактура груши
ОСИНА. Древесина мягкая с красивой текстурой. Легко режется и хорошо окрашивается.
ГРЕЦКИЙ ОРЕХ. Цвет древесины серовато- бурый. Текстура очень красивая, особенно у корня. Хорошо полируется.
Хорошо полируется.
Рис. 19. Фактура грецкого ореха
При изготовлении бижутерии предпочтение необходимо отдавать ценным породам деревьев с красивой текстурой и богатой палитрой красок.
Благодаря особым свойствам осина считается наиболее предпочтительным материалом для отделки русской бани. Чтобы русская баня была эффективной в эксплуатации и служила долго, лучше всего применять воздушную сушку на открытом воздухе. Это метод считается естественным и экономичным.
Уже при проектировании бани следует предусмотреть, как лучше проводить ремонтные работы, обеспечить доступ к деревянным конструкциям, чтобы их обработать. Специалисты советуют сделать все возможное, чтобы дерево не имело точек соприкосновения ни с толем или рубероидом, ни с камнем. Нужно позаботиться, чтобы хорошо действовала вентиляция. В бане в большинстве случаев не устраивают принудительную вентиляцию, а создание условий для достаточно интенсивного естественного воздухообмена – это довольно сложная конструктивная задача.
Чтобы на потолке не образовывался конденсат, его нужно как следует утеплить.
Для внешней обработки древесины чаще всего применяют олифу. Чтобы защитить строительные материалы от повреждения микроорганизмами, способствующими гниению древесины, ее обрабатывают специальными антисептическими и защитными средствами.
Scope and Extent of Wood Biology
Angyalossy V, Angeles G, Pace MR, Lima AC, Dias-Leme CL, Lohmann LG, Madero-Vega C (2012) Обзор анатомии, развития и эволюции сосудистой системы лиан. Завод Ecol Divers 5(2):167–182
CrossRef Google Scholar
Аренд М., Фромм Дж. (2007) Сезонные изменения реакции развития клеток древесины тополя на засуху. Физиол дерева 27: 985–992
Перекрёстная ссылка пабмед Google Scholar
 ) Эволюция физиологии растений от целых растений к экосистемам, серия симпозиумов Линнеевского общества №. 21. Elsevier Academic Press, Лондон, стр. 273–296
) Эволюция физиологии растений от целых растений к экосистемам, серия симпозиумов Линнеевского общества №. 21. Elsevier Academic Press, Лондон, стр. 273–296CrossRef. Google Scholar
Бейкер П.Дж., Палмер Дж.Г., Д’Арриго Р. (2008) Дендрохронология Callitris intratropica в северной Австралии: структура годового кольца, хронология развития и климатические корреляции. Aust J Bot 56: 311–320
CrossRef Google Scholar
Браунинг А., Фолланд-Фойгт Ф., Бурхардт И., Ганжи О., Наусс Т., Петерс Т. (2009) Климатический контроль радиального роста Cedrela montana во влажных горных тропических лесах на юге Эквадора. Erdkunde 63:337–345
CrossRef Google Scholar
Brienen RJW, Zuidema PA (2005) Связь роста деревьев с количеством осадков в тропических лесах Боливии: тест для шести видов с использованием анализа годичных колец. Экология 146:1–12
Экология 146:1–12
CrossRef пабмед Google Scholar
Brienen RJW, Lebrija-Trejos E, Zuidema PA, Martinez-Ramos M (2010) Анализ климата и роста мексиканского сухого лесного дерева показывает сильное влияние температуры поверхности моря и прогнозирует снижение роста в будущем. Глоб Чанг Биол 16: 2001–2012
Перекрёстная ссылка Google Scholar
Буччи С.Дж., Гольдштейн Г., Мейнцер Ф.К., Шольц Ф.Г., Франко А.С., Бустаманте М. (2004) Функциональная конвергенция в гидравлической архитектуре и водные отношения деревьев тропической саванны: от листьев до всего растения. Tree Physiol 24: 891–899
CrossRef КАС пабмед Google Scholar
Чемберс К.Дж., Хигучи Н., Шимель Дж.П. (1998) Древние деревья в Амазонии. Природа 39:135–136
Перекрёстная ссылка Google Scholar
Чав Дж. , Кумс Д., Янсен С., Льюис С.Л., Свенсон Н.Г., Занне А.Е. (2009) К мировому спектру экономики древесины. Ecol Lett 12:351–366
, Кумс Д., Янсен С., Льюис С.Л., Свенсон Н.Г., Занне А.Е. (2009) К мировому спектру экономики древесины. Ecol Lett 12:351–366
CrossRef пабмед Google Scholar
Чоат Б., Сак Л., Холбрук Н.М. (2007) Разнообразие гидравлических характеристик девяти видов Cordia , произрастающих в тропических лесах с контрастными осадками. Новый Фитол 175: 686–698
Перекрёстная ссылка пабмед Google Scholar
Чоудхури М.К., Шмитц Н., Верхейден А., Сасс-Класен У., Кудам Н., Бекман Х. (2008) Природа и периодичность годичных колец у двух видов бангладешских мангровых зарослей. IAWA J 29: 265–276
CrossRef Google Scholar
Кук Э.Р., Анчукайтис К.Дж., Бакли Б.М., Д’Арриго Р.Д., Джейкоби Г.К., Райт В.Е. (2010) Отсутствие муссонов в Азии и мегазасуха в течение последнего тысячелетия. Наука 328: 486–489
Перекрёстная ссылка КАС пабмед Google Scholar
Dezzeo N, Worbes M, Ishii I, Herrera R (2003) Годовые кольца деревьев, выявленные радиоуглеродным анализом в сезонно затопляемых лесах реки Мапире, притока нижнего течения реки Ориноко, Венесуэла. Завод Ecol 168:165–175
Завод Ecol 168:165–175
CrossRef Google Scholar
Die A, Kitin P, N’Guessan Kouame F, Bulcke JVD, Acker JV, Beeckman H (2012) Колебания камбиальной активности по отношению к осадкам приводят к образованию годовых колец и внутригодовых зон роста ксилемы и флоэмы в тик ( Tectona grandis ) в Кот-д’Ивуаре. Энн Бот 110:861–873
CrossRef КАС пабмед ПабМед Центральный Google Scholar
Domec JC, Scholz FG, Bucci SJ, Meinzer FC, Goldstein G, Villalobos-Vega R (2006)Суточные и сезонные колебания корневой ксилемной эмболии у древесных видов неотропической саванны: влияние на устьичный контроль состояния воды в растениях. Plant Cell Environment 29:26–35
CrossRef КАС пабмед Google Scholar
Dünisch O, Montoia VR, Bauch J (2003) Дендроэкологические исследования Swietenia macrophylla King и Cedrela odorata L. (Meliaceae) в центральной части Амазонки. Деревья 17:244–250
(Meliaceae) в центральной части Амазонки. Деревья 17:244–250
Google Scholar
Ennos AR (1993) Функция и формирование контрфорсов. Trends Ecol Evol 8:350–351
CrossRef КАС пабмед Google Scholar
Fan ZX, Zhang S-B, Hao GY, Slik JWF, Cao KF (2012) Признаки гидравлической проводимости предсказывают скорость роста и рост взрослых 40 азиатских тропических пород деревьев лучше, чем плотность древесины. J Ecol 100:732–741
CrossRef Google Scholar
Фихтлер Э., Ворбс М. (2012) Анатомические переменные древесины тропических деревьев и их связь с местными условиями и морфологией отдельных деревьев. IAWA J 33(2):119–140
Google Scholar
Фихтлер Э., Кларк Д.А., Уорбс М. (2003) Возраст и долгосрочный рост деревьев в старовозрастных тропических лесах на основе анализа годичных колец и C-14. Биотропика 35:306–317
Биотропика 35:306–317
CrossRef Google Scholar
Fichtler E, Helle G, Worbes M (2010) Временные ряды стабильных изотопов углерода из годичных колец тропических деревьев указывают на сигнал осадков. Дерево-кольцо Res 66: 35–49
Перекрёстная ссылка Google Scholar
Фонти П., Аркс Г., Гарсия-Гонсалес И., Эйльманн Б., Сасс-Клаассен У., Гертнер Х., Экштейн Д. (2010) Изучение глобальных изменений посредством исследования пластических реакций анатомии ксилемы в годичных кольцах деревьев. Новый Phytol 185:42–53
CrossRef пабмед Google Scholar
Фромм Дж. (2013)Развитие ксилемы у деревьев: от камбиальных делений до зрелых клеток древесины. В: Фромм Дж. (ред.) Клеточные аспекты формирования древесины, Монографии клеток растений. Springer, Берлин, стр. 3–39.
Перекрёстная ссылка Google Scholar
Fromm J, Sautter I, Matties D, Kremer J, Schumacher P, Ganter C (2001) Содержание воды в ксилеме и плотность древесины елей и дубов, обнаруженные с помощью компьютерной томографии высокого разрешения. Физика растений 127:416–425
Физика растений 127:416–425
CrossRef КАС Google Scholar
Fu P-L, Jiang Y-J, Wang A-Y, Brodribb TJ, Zhang J-L, Zhu S-D, Cao K-F (2012) Гидравлические характеристики стебля и устойчивость листьев к водному стрессу согласуются с фенологией листьев покрытосеменных деревьев в Азии. тропический сухой карстовый лес. Энн Бот 110:189–199
Перекрестная ссылка пабмед ПабМед Центральный Google Scholar
Гарсия-Гонсалес И., Экштейн Д. (2003) Климатический сигнал сосудов из раннего дуба на морском участке. Tree Physiol 23: 497–504
CrossRef Google Scholar
Guy CL, Huber JL, Huber SC (1992) Сахарозофосфатсинтаза и накопление сахарозы при низкой температуре. Физика растений 100:502–508
Перекрёстная ссылка КАС Google Scholar
Hacke UG, Sperry JS, Pockman WT, Davis SD, McCulloch KA (2001) Тенденции изменения плотности и структуры древесины связаны с предотвращением разрушения ксилемы отрицательным давлением. Экология 126:457–461
Экология 126:457–461
CrossRef Google Scholar
He Z, Tang Y, Deng X, Cao M (2013) Деревья-контрфорсы в тропическом лесу диптерокарповых площадью 20 гектаров в Сишуанбаньна, юго-запад Китая. J Plant Ecol 6 (2): 187–192
Google Scholar
Hietz P, Valencia R, Wright SJ (2013) Сильные радиальные колебания плотности древесины соответствуют единой схеме в двух неотропических тропических лесах. Funct Ecol 27:684–692
CrossRef Google Scholar
Холбрук Н.М., Путц Ф.Е. (1996) Физиология тропических лиан и полуэпифитов: растения, которые взбираются вверх, и растения, которые спускаются вниз. В: Малки С.С., Чаздон Р.Л., Смит А.П. (ред.) Экофизиология растений тропических лесов. Чепмен и Холл, Нью-Йорк, стр. 363–39.3
Перекрёстная ссылка Google Scholar
Höll W (2000) Распределение, колебания и метаболизм пищевых запасов в древесине деревьев. В: Savidge R, Barnett J, Napier R (eds) Клеточная и молекулярная биология образования древесины. BIOS, Оксфорд, стр. 347–362
В: Savidge R, Barnett J, Napier R (eds) Клеточная и молекулярная биология образования древесины. BIOS, Оксфорд, стр. 347–362
Google Scholar
Хуа К., Барбетти М., Ворбс М., Хед Дж., Левченко В.А. (1999) Обзор радиоуглеродных данных из проб атмосферы и годичных колец за период 1945–1997 г. н.э. IAWA J 20:261–283
CrossRef Google Scholar
Якобсен А.Л., Пратт Р.Б., Тобин М.Ф., Хаке Ю.Г., Эверс Ф.В. (2012)Глобальный анализ длины сосудов ксилемы у древесных растений. Am J Bot 99(10):1583–1591
CrossRef пабмед Google Scholar
Johnson DM, Woodruff DR, McCulloh KA, Meinzer FC (2009) Гидравлическая проводимость листьев, измеренная на месте, снижается и восстанавливается ежедневно: гидравлика листьев, водный потенциал и устьичная проводимость у четырех умеренных и трех тропических видов деревьев. Физиол дерева 29:879–887
Физиол дерева 29:879–887
Перекрёстная ссылка КАС пабмед Google Scholar
Kallarackal J, Otieno DO, Reineking B, Jung E-Y, Schmidt MWT, Granier A, Tenhunen JD (2013) Функциональная конвергенция в использовании воды деревьями из разных географических регионов: метаанализ. Деревья 27:787–799
CrossRef Google Scholar
Кинг Д.А., Дэвис С.Дж., Тан С., Супарди Н., Нур М. (2006) Роль плотности древесины и стоимости поддержки ствола в росте и гибели тропических деревьев. Дж Экол 94:670–680
Перекрёстная ссылка Google Scholar
Lautner S (2013) Формирование древесины в условиях засухи и засоления. В: Фромм Дж. (ред.) Клеточные аспекты формирования древесины, Монографии по растительным клеткам. Springer, Берлин, стр. 187–202
CrossRef Google Scholar
Льюис С. Л., Мали Ю., Филлипс О.Л. (2004) Анализ влияния глобальных изменений на тропические леса. Philos Trans R Soc Lond B Biol Sci 359:437–462
Л., Мали Ю., Филлипс О.Л. (2004) Анализ влияния глобальных изменений на тропические леса. Philos Trans R Soc Lond B Biol Sci 359:437–462
Перекрёстная ссылка КАС пабмед ПабМед Центральный Google Scholar
Льюис С.Л., Ллойд Дж., Ситч С., Митчард Э.Т.А., Лоранс В.Ф. (2009) Изменение экологии тропических лесов: доказательства и движущие силы. Annu Rev Ecol Evol Syst 40:529–549
CrossRef Google Scholar
Либерман Д., Либерман М., Хартсхорн Г.С., Перальта Р. (1985) Темпы роста и возрастные соотношения деревьев влажных тропических лесов в Коста-Рике. J Троп Эколь 1:97–109
Перекрёстная ссылка Google Scholar
Lisi CS, Tomazello M, Botosso PC, Roig FA, Maria VRB, Ferreira-Fedele L, Voigt ARA (2008) Формирование годичных колец, периодичность радиального прироста и фенология видов деревьев из сезонного полулиственного леса на юго-востоке Бразилии. IAWA J 29:189–207
IAWA J 29:189–207
CrossRef Google Scholar
Lloyd J, Farquhar GD (2008) Влияние повышения температуры и CO 2 по физиологии деревьев тропических лесов. Philos Trans R Soc Lond B Biol Sci 363:1811–1817
CrossRef КАС пабмед ПабМед Центральный Google Scholar
Magel E (2001) Физиология камбиального роста, хранение запасов и формирование сердцевины. В: Heikkilä H, Bucher J, Sundberg B, Jarvis P, Matyssek R, Huttunen S (eds) Тенденции в исследованиях физиологии европейских лесных деревьев. Клювер, Дордрехт, стр. 19–32
Google Scholar
Мартинес-Кабрера Х.И., Джонс К.С., Эспино С., Шенк Х.Дж. (2009) Анатомия древесины и плотность древесины в кустарниках: реакция на различную засушливость на трансконтинентальных трансектах. Am J Bot 96: 1388–1398
CrossRef пабмед Google Scholar
Martinez-Ramos M, Alvarez-Buylla ER (1998) Сколько лет деревьям тропических лесов? Trends Plant Sci 3(10):400–405
Перекрёстная ссылка Google Scholar
McCulloh K, Sperry JS, Lachenbruch B, Meinzer FC, Reich PB, Voelker S (2010) Движущаяся водяная скважина: сравнение гидравлической эффективности веток и стволов хвойных, кольцевидно-пористых и диффузно-пористых саженцев из умеренных и тропические леса. Новый Phytol 86: 439–450
Новый Phytol 86: 439–450
CrossRef Google Scholar
Meinzer FC (2003) Функциональная конвергенция реакций растений на окружающую среду. Экология 134:1–11
Перекрёстная ссылка пабмед Google Scholar
Müller-Landau HC (2004) Межвидовые и межместные различия удельного веса древесины тропических деревьев. Биотропика 36:20–32
Google Scholar
Newbery DM, Schwan S, Chuyong GB, van der Burgt XM (2009) Опорная форма дерева тропических лесов Центральной Африки Microberlinia bisulcata и его возможная роль в усвоении питательных веществ. Деревья 23:219–234
Перекрестная ссылка Google Scholar
Нок К.А., Бейкер П.Дж., Ванек В., Лейс А., Грабнер М., Буньявейчевин С., Хитц П. (2010) Долгосрочное повышение внутренней эффективности водопользования не приводит к усилению роста стволов в тропическом муссонном лесу в запад Таиланда. Глоб Чанг Биол. doi:10.1111/j.1365-2468.2010.02222.x
Глоб Чанг Биол. doi:10.1111/j.1365-2468.2010.02222.x
Google Scholar
Олсен Д.М., Динерштейн Э., Викраманаяке Э.Д., Берджесс Н.Д., Пауэлл Г.В.Н., Андервуд Э.К., Д’Амико Дж., Итуа И., Стрэнд Х.Е., Моррисон Д.С., Лукс С.Дж., Оллнатт Т.Ф., Рикеттс Т.Х., Кура И., Ламоре Д.Ф. , Веттенгель В.В., Хедао П., Кассем К.Р. (2001) Наземные экорегионы мира: новая карта жизни на Земле. Бионаука 51:933–938
Перекрёстная ссылка Google Scholar
Pandey CB, Singh L, Singh SK (2011) Неоднородность среды обитания, вызванная контрфорсами, увеличивает доступность азота в тропических лесах. Для Ecol Manage 262:1679–1685
CrossRef Google Scholar
Филлипс Н., Орен Р., Циммерманн Р. (1996) Радиальные паттерны потока сока ксилемы у недиффузных и кольцевидных пород деревьев. Окружающая среда растительных клеток 19:983–990
Перекрёстная ссылка Google Scholar
Пратт Р. Б., Якобсен А.Л., Эверса Ф.В., Дэвис С.Д. (2007) Взаимосвязь между транспортом ксилемы, биомеханикой и хранением в стеблях и корнях девяти видов Rhamnaceae калифорнийского чапараля. Новый Phytol 174:787–798
Б., Якобсен А.Л., Эверса Ф.В., Дэвис С.Д. (2007) Взаимосвязь между транспортом ксилемы, биомеханикой и хранением в стеблях и корнях девяти видов Rhamnaceae калифорнийского чапараля. Новый Phytol 174:787–798
CrossRef КАС пабмед Google Scholar
Престон К.А., Корнуэлл В.К., ДеНойер Дж.Л. (2006) Плотность древесины и особенности сосудов как отдельные корреляты экологической стратегии у 51 покрытосеменных растений побережья Калифорнии. Новый фитол 170: 807–818
Перекрёстная ссылка пабмед Google Scholar
Пумиджумнонг Н., Буаджан С. (2013) Сезонная камбиальная активность пяти видов тропических деревьев в центральном Таиланде. Деревья 27:409–417
CrossRef Google Scholar
Рана Р., Лангенфельд-Хейзер Р., Финкельдей Р., Полле А. (2009) Функциональная анатомия пяти находящихся под угрозой исчезновения тропических пород древесины семейства Dipterocarpaceae. Деревья 23: 521–529
Деревья 23: 521–529
Перекрёстная ссылка Google Scholar
Richards PW (1996) Влажный тропический лес, 2-е изд. Издательство Кембриджского университета, Кембридж
Google Scholar
Родерик М.Л., Берри С.Л. (2001) Связь плотности древесины с ростом деревьев и окружающей средой: теоретический анализ, основанный на движении воды. Новый Phytol 149:473–485
CrossRef Google Scholar
Розендал Д.А., Зуидема П.А. (2011) Дендроэкология в тропиках: обзор. Деревья 25:3–16
CrossRef Google Scholar
Rozendaal DMA, Brienen RJW, Soliz-Gamboa CC, Zuidema PA (2010) Кольцевые кольца тропических деревьев показывают предпочтительное выживание быстрорастущих молодых особей и увеличение темпов роста молодых особей с течением времени. Новый Phytol 185:759–769
CrossRef пабмед Google Scholar
Руссо С. Е., Дженкинс К.Л., Уизер С.К., Уриат М., Дункан Р.П., Кумс Д.А. (2010) Межвидовые отношения между признаками роста, смертности и ксилемы древесных видов из Новой Зеландии. Funct Ecol 24:253–262
Е., Дженкинс К.Л., Уизер С.К., Уриат М., Дункан Р.П., Кумс Д.А. (2010) Межвидовые отношения между признаками роста, смертности и ксилемы древесных видов из Новой Зеландии. Funct Ecol 24:253–262
CrossRef Google Scholar
Saldarriagga JG, Западный округ Колумбия, Tharp ML, Uhl C (1988) Долговременная хронология сукцессии лесов в верховьях Рио-Негро в Колумбии и Венесуэле. Дж. Экол. 76:938–958
перекрестная ссылка Google Scholar
Sass-Klaassen U, Couralet C, Sahle Y, Sterck FJ (2008) Можжевельник из Эфиопии содержит крупномасштабный сигнал осадков. Int J Plant Sci 169:1057–1065
CrossRef Google Scholar
Sauter JJ, Witt W (1997) Структура и функция лучей: хранение, мобилизация, транспорт. В: Ренненберг Х., Эшрих В., Циглер Х. (ред.) Деревья — вклад в современную физиологию деревьев. Академическое издательство SFB, Гаага, стр. 177–19.5
177–19.5
Google Scholar
Schöngart J, Piedade MTF, Ludwigshausen S, Worbes M (2002) Фенология и периодичность стволового роста древесных пород в пойменных лесах Амазонки. J Trop Ecol 18:1–17
CrossRef Google Scholar
Schöngart J, Piedade MTF, Wittmann F, Junk WJ, Worbes M (2005) Модели роста древесины Macrolobium acaciifolium (Benth.) Benth. (Fabaceae) в амазонских пойменных лесах черноводной и бурной воды. Экология 145:454–461
Перекрёстная ссылка пабмед Google Scholar
Шульдт Б., Лойшнер С., Брок Н., Хорна В. (2013) Изменения плотности древесины, анатомии древесины и гидравлических свойств ксилемы на пути потока от корня к побегу деревьев тропических лесов. Tree Physiol 33:161–174
CrossRef пабмед Google Scholar
Смит А. П. (1972) Укрепление тропических деревьев: описательная модель и новые гипотезы. Ам Нат 106:32–46
П. (1972) Укрепление тропических деревьев: описательная модель и новые гипотезы. Ам Нат 106:32–46
Перекрёстная ссылка Google Scholar
Sperry JS (2003) Эволюция водного транспорта и структуры ксилемы. Int J Plant Sci 164:115–127
CrossRef Google Scholar
Swaine MD, Whitmore TC (1988) Об определении групп экологических видов во влажных тропических лесах. Завод Ecol 75:81–86
CrossRef Google Scholar
Therell MD, Stahle DW, Ries LP, Shugart HH (2006) Реконструкция изменчивости осадков по годичным кольцам деревьев в Зимбабве. Клим Дин 26: 677–685
CrossRef Google Scholar
Therell MD, Stahle DW, Mukelabai MA, Shugart HH (2007) Возраст и динамика радиального роста Pterocarpus angolensis на юге Африки. Для Ecol Manage 244:24–31
CrossRef Google Scholar
Tromp J, Ovaa IC (1973) Весенняя мобилизация белкового азота в коре яблони. Physiol Plant 29:1–5
Physiol Plant 29:1–5
CrossRef КАС Google Scholar
Tyree MT (2003) Гидравлические ограничения производительности деревьев: транспирация, накопление углерода и рост деревьев. Деревья 17:95–100
Google Scholar
Tyree MT, Zimmermann MH (2002) Структура ксилемы и восхождение сока, Rev edn. Спрингер, Берлин/Гейдельберг/Нью-Йорк
Google Scholar
Verheyden A, De Ridder F, Schmitz N, Beeckman H, Koedam N (2005) Временные ряды плотности сосудов в кенийских мангровых деревьях с высоким разрешением показывают связь с климатом. Новый Phytol 167:425–435
CrossRef пабмед Google Scholar
Wheeler EA, Baas P, Rodgers SA (2007) Вариации анатомии древесины двудольных растений: глобальный анализ на основе базы данных InsideWood. ИАВА Дж 28:229–258
Перекрестная ссылка Google Scholar
Wiemann MC, Williamson GB (1988) Экстремальные радиальные изменения удельного веса древесины у некоторых пионеров тропиков. Wood Fiber Sci 20:344–349
Wood Fiber Sci 20:344–349
Google Scholar
Wiemann MC, Williamson GB (1989) Радиальные градиенты удельного веса древесины некоторых тропических и умеренных деревьев. Лесоведение 35:197–210
Google Scholar
Wils THG, Robertson I, Eshetu Z, Touchan R, Sass-Klaassen U, Koprowski M (2010) Crossdating Juniperus procera из Северного Гондара. Эфиопские деревья. дои: 10.1007/s00468-010-0475-0
Google Scholar
Worbes M (2002) Сто лет исследований годичных колец в тропиках – краткая история и взгляд на будущие проблемы. Дендрохронология 20(1–2):217–231
CrossRef Google Scholar
Zanne AE, Westoby M, Falster DS, Ackerly DD, Loarie SR, Arnold SEJ, Coomes DA (2010)Структура древесины покрытосеменных растений: глобальные закономерности в анатомии сосудов и их связь с плотностью древесины и потенциальной проводимостью. Am J Bot 97: 207–215
Am J Bot 97: 207–215
CrossRef пабмед Google Scholar
Zheng J, Martinez-Cabrera HI (2013) Анатомические корреляции древесины с теоретической проводимостью и плотностью древесины в Китае: эволюционные доказательства функциональной дифференциации осевой и радиальной паренхимы. Энн Бот 112:927–935
Перекрестная ссылка пабмед ПабМед Центральный Google Scholar
Zhu S-D, Song J-J, Li R-H, Ye Q (2013) Гидравлика растений и фотосинтез 34 древесных видов из разных сукцессионных стадий субтропических лесов. Plant Cell Environ 36:879–891
CrossRef КАС пабмед Google Scholar
Zufferey V, Cochard H, Ameglio T, Spring JL, Viret O (2011)Суточные циклы образования и восстановления эмболии в черешках виноградной лозы (Vitis vinifera cv. Chasselas). J Exp Бот 62: 3885–3894
Перекрёстная ссылка КАС пабмед ПабМед Центральный Google Scholar
Звинецкий М. А., Холбрук Н.М. (2009) Противостояние демону Максвелла: биофизика восстановления ксилемной эмболии. Trends Plant Sci 14:530–534
А., Холбрук Н.М. (2009) Противостояние демону Максвелла: биофизика восстановления ксилемной эмболии. Trends Plant Sci 14:530–534
CrossRef КАС пабмед Google Scholar
Летопись лесоведения | Цели и область применения
Annals of Forest Science — международный исследовательский журнал, посвященный междисциплинарным исследованиям лесов и древесины в меняющемся мире . Это один из журналов Французского национального института сельского хозяйства, продовольствия и окружающей среды ( INRAE ), государственного некоммерческого исследовательского учреждения.
Область применения Annals of Forest Science охватывает следующие области исследований:
- Биология деревьев и связанных с ними организмов
- Динамика лесов и экосистемные процессы под воздействием факторов окружающей среды или управления (экология, генетика)
- Риски и нарушения, влияющие на лесные экосистемы (биология, экология, экономика)
- Леса как движущая сила устойчивой, многооборотной и инклюзивной биоэкономики (разведение деревьев, лесопользование и продуктивность, экосистемные услуги, лесоводство)
- Науки о древесине (взаимосвязь между структурой древесины и функционированием дерева, а также между управлением лесным хозяйством или окружающей средой и свойствами древесины)
Редколлегия приветствует:
- Новые идеи или методы измерения или мониторинга любое свойство, представляющее интерес для биологии деревьев, динамики лесных экосистем, древесины
- Документы на основе моделей посвящены цепочке лесохозяйственной деятельности, включая исследования по учету углерода, экосистемные услуги, количественную оценку взаимосвязей между управлением лесным хозяйством и свойствами конечной продукции
- Исследования по экономическим и социальным вопросам лесопользования в условиях изменения климата и лесной биоэкономики
- Обзорные статьи на актуальные темы в рамках журнала
- Документы с данными , представляющие базы данных, доступные более широкому сообществу
- Авторские статьи , представляющие аргументы по открытым исследовательским вопросам
Редколлегия не поощряет:
- Локальные тематические исследования на основе одностендовых экспериментов или небольших выборок в местных условиях, если рассматриваемые вопросы не являются инновационными и своевременными
- Чисто технологические статьи представляющие приложения без более широкой научной перспективы
- Статьи по дереву, посвященные чисто технологическим вопросам , если только тема не является очень инновационной или не связана с биологией деревьев, экологией леса или управлением лесным хозяйством
Подать рукопись
Официальный журнал
- Редакционная коллегия
- Домашняя страница ИНРАЭ
- Связаться с редакцией
- Цели и объем
- Рекомендации по отправке
- Флаер «Анналы лесоведения»
- Флаер с данными
- Дебаты
- Портал журналов ИНРАЭ, издаваемых Springer Nature
- Содержание анналов лесоведения до 2022 г.
