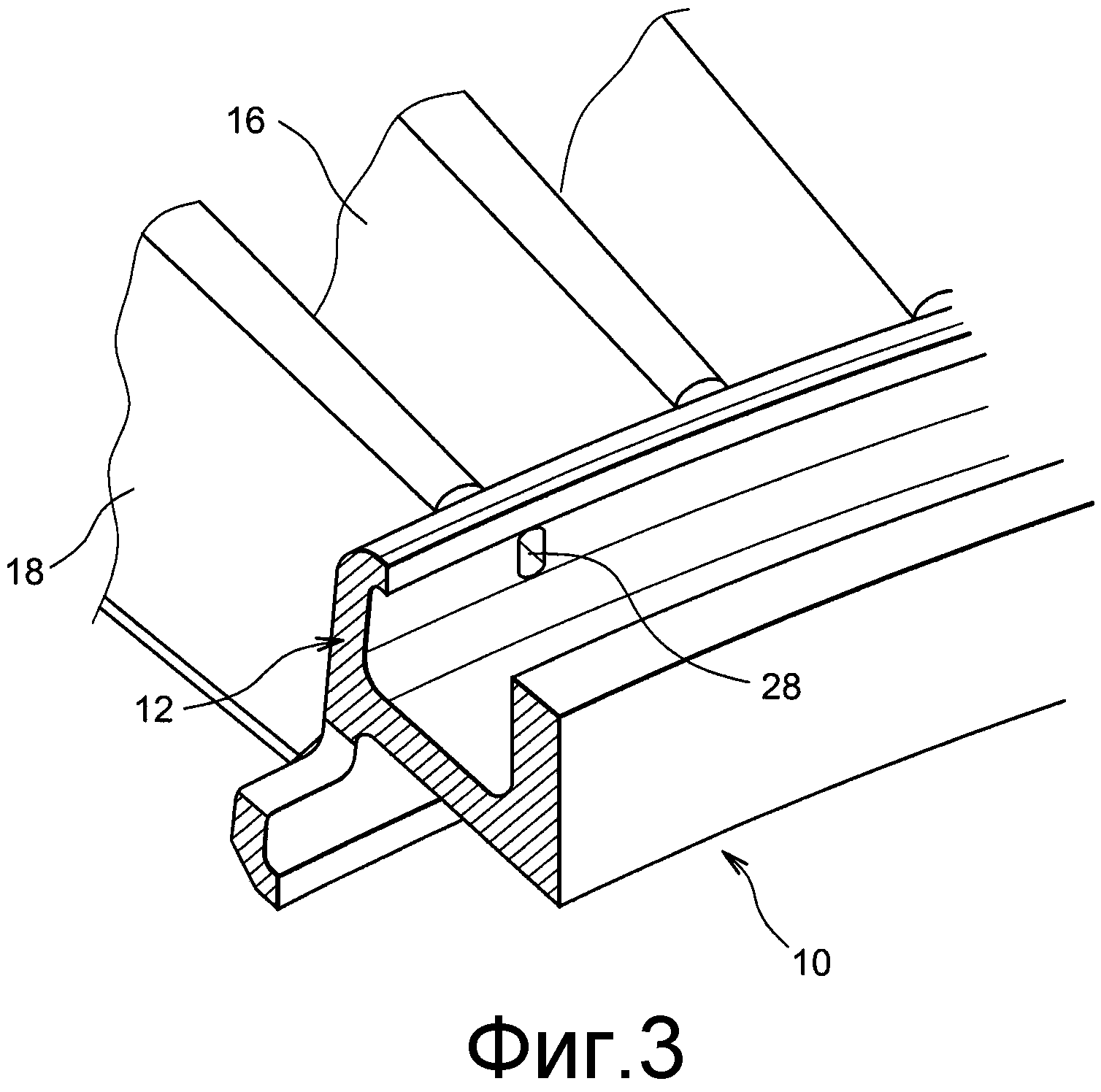
МВС-3Т — Модельный восковый состав
Модельный восковый состав МВС-3Т предназначен для изготовления моделей, используемых в литейном производстве, для литья по выплавляемым моделям (ЛВМ)черных и цветных металлов.
В зависимости от способа приготовления и области применения модельный восковый состав выпускается следующих марок:
|
Наименование |
Аналог |
Описание |
|
МВС-3Т/01 |
МВС-3А, МВС-3АЯ, МВС-3Н, Р-3Н |
Базовый состав для литья по выплавляемым моделям отливок различной категории сложности, изготовлен по стандартной технологии путем гомогенизации парафина, церезина, синтетических и природных восков. |
|
МВС-3Т/02 |
Состав для литья по выплавляемым моделям отливок особой категории сложности — ответственное, тонкостенное, сверхточное литье заготовок (колесо турбины и т.п.) в т.ч. для предприятий оборонного комплекса, изготовлен по особой технологии с улучшением реологических свойств композиции базового материала. |

Физико-химические показатели:
|
|
Наименование показателя |
норма | |
|
МВС-3Т/01 |
МВС-3Т/02 |
||
|
1 |
Внешний вид |
Брикеты, блины от желтого до коричневого цвета |
|
|
2 |
Температура каплепадения,ОС |
80-90 |
|
|
3 |
Предел прочности при статическом изгибе при 18-20ОС, кг/см2 (МПа), не менее |
50 (5,0) |
|
|
4 |
Теплоустойчивость,ОС, не менее |
37 |
|
|
5 |
Кислотное число, мг КОН на г состава, не более |
0,5 |
0,4 |
|
6 |
Массовая доля золы, %, не более |
0,03 |
|
Отличительные свойства:
— в состав материала дополнительно введен структурообразующий сополимер;
— в состав материала дополнительно введена антиокислительная присадка
Получаемые преимущества:
— сокращается расход модельной массы на 1шт. годных отливок на 10-15%;
годных отливок на 10-15%;
— уменьшается брак моделей по недоливам и различной точности;
— сокращаются потери моделей при извлечении их из пресс-форм за счет прочности композиции;
— обеспечивается стабильность технологических свойств состава;
— улучшается чистота поверхности;
— повышается трещиноустойчивость, твердость моделей.
Меры безопасности:
МВС-3Т в соответствие с ГОСТ 12.1.044 относится к горючим, пожароопасным веществам. Температура вспышки не менее 190 ºС, воспламенения не менее 220 ºС.
При работе с МВС-3Т необходимо применять индивидуальные средства защиты по ГОСТ 12.4.011, ГОСТ 12.4.103, ГОСТ 12.4013-85Е, ГОСТ 12.4010-75.
Оборудование и помещение, при работе с составом модельным восковым МВС-3Т, должны быть оборудованы общеобменной приточно-вытяжной вентиляцией и местными отсосами.
Транспортировка и хранение:
МВС-3Т транспортируют и хранят в соответствии с ГОСТ 1510.
Модельный восковой состав в Нижнем Новгороде (Составы модельно-восковые)
Различают следующие марки модельных восковых составов:СДС-13М для сыров (ТУ 38.101225-94) — представляет собой композицию глубокоочищенного нефтяного воска с полимерными добавками.
ВН-2 (ТУ 38.401210-93) — в качестве основы применяют, воск который представляет собой фракцию твердых парафиновых углеводородов, выделенных обезмасливанием смеси петролатума и целевого фильтрата, полученного в процессе обезмасливания широкой фракции гача. Для использования в сплаве СДС-13М воск ВН-2 подвергают глубокой очистке.
МВС-ЗА, МВС-3Т\01 и МВС-3Т\02 (ТУ 38.101516-76 и ТУ 0255-001-71462031-2005) МВС-3Н (аналог МВС-3А) представляет собой композицию глубокообезмасленного парафина и высокоплавкого нефтяного церезина с полимерной добавкой. Предназначен для точного стального литья по выплавляемым моделям.
МВС-15 (ТУ 38.1011044-85 и ТУ 0255-001-71462031-2005) представляет собой композицию высокоплавкого церезина с техническим парафином и полимерной добавкой. Р-3Н (аналог Р-3). Р-3,ПЦБ (ТУ 0255-001-71462031-2005). Г-1М-2 (ТУ 6-00-010-5000597-94) (аналог ВИАМ102).
Р-3Н (аналог Р-3). Р-3,ПЦБ (ТУ 0255-001-71462031-2005). Г-1М-2 (ТУ 6-00-010-5000597-94) (аналог ВИАМ102).
Модельный состав «Г-1» представляет собой сплав на основе природных восков и парафинов, который предназначен для изготовления моделей отливок в литье по выплавляемым моделям предприятий авиационной и судостроительной промышленности.
Технические показатели ТУ 0255-026-71462031-2010 Норма по ТУ Наименование показателей Г-1М-1 Г-1М-2 1. Внешний вид при 20гр.С Твердое воскообразное вещество темно-коричневого цвет 2.Предел прочности пристатическом изгибе притемпературе (19+-1)гр. С, кг/см.2, не менее 45 38 3. Теплоустойчивость, гр.С , не менее 40 40 4. Температура каплепадения 79-87 79-92 Наименование показателей Модельный состав «МВС-3А» ТУ 0255-001-71462031-2005 1.Внешний вид при 20гр.С Твердое воскообразное вещество 2.Предел прочности пристатическом изгибе при температуре (19+-1)гр. С, кг/см.2, не менее 40 3. Теплоустойчивость, гр.С , не менее 40 4.Температура каплепадения 85-95 Применение Модельные восковые составы нашли применение в различных аспектах человеческой деятельности:
Внешний вид при 20гр.С Твердое воскообразное вещество темно-коричневого цвет 2.Предел прочности пристатическом изгибе притемпературе (19+-1)гр. С, кг/см.2, не менее 45 38 3. Теплоустойчивость, гр.С , не менее 40 40 4. Температура каплепадения 79-87 79-92 Наименование показателей Модельный состав «МВС-3А» ТУ 0255-001-71462031-2005 1.Внешний вид при 20гр.С Твердое воскообразное вещество 2.Предел прочности пристатическом изгибе при температуре (19+-1)гр. С, кг/см.2, не менее 40 3. Теплоустойчивость, гр.С , не менее 40 4.Температура каплепадения 85-95 Применение Модельные восковые составы нашли применение в различных аспектах человеческой деятельности:- на предприятиях машиностроения для точного стального литья;
- на заводах тракторного и сельскохозяйственного машиностроения для литья по выплавляемым моделям в цехах точного литья;
- для защиты твердых сычужных сыров от высыхания и плесневения в период их вызревания и хранения. Подходят только глубокоочищенные воски, допущенные к контактам с пищевыми продуктами.
Если Вы решили купить модельный восковой состав оптом или в розницу в ПКФ «Нижегородхимпродукт», то можете быть уверены, что цена Вас приятно удивит. Обусловлено это тем, что являясь прямым дилером заводов, «Нижегородхимпродукт» обеспечивает прямые поставки от производителей по всей России.
Наличие прямых дилерских контрактов с производителями у ПКФ «Нижегородхимпродукт» гарантирует не только то, что цена однозначно будет конкурентоспособной, но и прямые поставки от производителя оптом и в розницу по всей России обеспечат Вам приобретение продукции без наценки!
Убедитесь сами!
ФотографииКупить модельный восковой составЗаказать93RUB ЗаказатьИли свяжитесь с продавцомСообщениеПостарайтесь кратко описать суть вашего вопроса продавцу (минимум 20 символов)ФИОТелефонE-mailОтправляя вопрос, вы соглашаетесь с пользовательским соглашением ознакоми(лся/лась) и принимаю егоВернуться назадв раздел «Составы модельно-восковые» Смотрите также товары категории «Формовочное литейное оборудование» Установки формовочные Машины формовочные Составы модельно-восковыеМодельный восковой состав мвс р-3н — Ярославский завод парафино-восковой продукции, АО Ростов (Россия)
Продукция: Модельный восковой состав МВС Р — 3Н цена купить оптом от производителя в России.
Харакетристики:
- Внешний вид: чешуйки, стружка, таблетки, монолит или брикеты от желтого до темно — коричневого цвета.
- Предназначение: предназначен для изготовления выплавляемых моделей при производстве отливок с требованиями по размерной точности в металлургической промышленности.
- Применение: Выплавка ювелирных изделий, выплавка деталей.
- Показатели качества:
СОГЛАСНО ТУ 0255 — 002 — 13933880 — 2015 С ИЗМ. 1
| Наименование показателя | Метод испытания | Норма по ТУ |
| Температура каплепадения, не менее, °С | ГОСТ 6793 | 70 |
| Предел прочности при статическом изгибе при (18 — 20)°С, кгс/см | п.4.4, 4.4.2 настоящих ТУ | 32 |
| Теплоустойчивость, °С не менее | п.4.4, 4.4.3 настоящих ТУ | 32 |
| Кислотное число, мг KOH на 1г состава, не более | ГОСТ 2488 | 4 |
| Массовая доля золы, % | ГОСТ 1461 | 0,03 |
Условия поставки: EXW, также возможны другие условия доставки
Условия оплаты: 100% предоплата, возможна частичная оплата
Производитель: Акционерное общество «Ярославский завод парафино — восковой продукции»
Перечень предлагаемой продукции:
- Парафины
- Гачи
- Воски
- Мягчители
- Церезины
- Петролатумы
- Антистаритель
- Стеарин
- Стеариновая кислота
- Пальмовый воск
- Буроугольный воск
- Модельные восковые составы
- Гидрофобные заполнители
- Масло лампадное
|
Модельные восковые составы МВС-15, МВС-3А, МВС-3Н, Р-3.
Уважаемые господа предлагаем Вам ряд восковых составов
централизованного производства, выпускаемых на нашем предприятии.
Продукт состоит из парафина и синтетических или природных восков.
Применяется для точного литья по выплавляемым моделям.
Надеемся на деловое и взаимовыгодное сотрудничество.
Качественные характеристики восковых модельных составов Р-3, МВС-3Н
приведены в таблице.
*- Данный восковой модельный состав МВС-3Н ТУ 0258-001-51570957-2002 аналогичен модельным массам МВС-3А, МВС-15, МС-В и является улучшенной, и более современной версией разработанной С –Петербургским институтом ОАО «Пластполимер»
Цены на МВС-3Н,Р-3 – договорные. .
Исходя из Ваших потребностей, наше предприятие имеет возможность изготовить модельный восковой состав с характеристиками необходимыми для вашего производства.
|
||
 Производим полиэтиленовые трубы различных диаметров и различной толщиной стенки. Расчищаем и углубляем малые реки,пруды,водемы,производственные пруды отстойники. Устанавливаем поливочные системы и системы подкорневого орошения.
Производим полиэтиленовые трубы различных диаметров и различной толщиной стенки. Расчищаем и углубляем малые реки,пруды,водемы,производственные пруды отстойники. Устанавливаем поливочные системы и системы подкорневого орошения.


Воск ПВ-200, воск ПВ-300, воск пчелиный , МВС-3А , воск буроугольный
Информация
ООО «РЕЗЕРВ НК» предлагает со склада Москвы : ПВ-200 применяется для подслоя на бумажную подложку в электронной промышленности, для заливочных составов в кабельной промышленности, для улучшения свойств полиграфических красок. ПВ-300 применяется для производства модельно- восковых составов. Воск пчелиный производственный , Срок хранения: не ограничен Условия хранения: не регламентируются Область применения: для приготовления косметических средств, полировочных мастик, водоотталкивающих пропиток для тканей, красок; при выделке кожи, а также в медицине. Воск идёт для приготовления свечей, искусственных цветов; для лепки вместо глины. При переработке из пчелиного воска можно получить два высококачественных продукта для косметики- душистый воск и липиды. Модельно-восковой состав МВС-3А является сплавом на основе природных парафинов и восков для изготовления выплавляемых моделей в процессе точного литья деталей в машиностроении.
ПВ-300 применяется для производства модельно- восковых составов. Воск пчелиный производственный , Срок хранения: не ограничен Условия хранения: не регламентируются Область применения: для приготовления косметических средств, полировочных мастик, водоотталкивающих пропиток для тканей, красок; при выделке кожи, а также в медицине. Воск идёт для приготовления свечей, искусственных цветов; для лепки вместо глины. При переработке из пчелиного воска можно получить два высококачественных продукта для косметики- душистый воск и липиды. Модельно-восковой состав МВС-3А является сплавом на основе природных парафинов и восков для изготовления выплавляемых моделей в процессе точного литья деталей в машиностроении.
Магазин «РЕЗЕРВ НК, ООО»
Другие предложения от РЕЗЕРВА НК, ООО
-
Этиленгликоль высший сорт 99, 8% в мелкой таре.

- Синтанол АЛМ-7, АЛМ-10.
- Присадки:Агидол-1;2, ДФ-11, С-300, Сигбол, С-5А, Детерсол-140, ПМА-Д, КП-2,
- Оксифос КД-6, Б, Б-1
- Рабочие жидкости РЖ-3, РЖ-8 для электро-эрозионных станков .
- Ортоксилол
- Масла индустриальные И-5, 8, 12, 20, 30, 40, 50 ₽1
-
Литол-24, Солидол, Циатим, ТМ-9П, ПВК, воск и др.

- ЦИАТИМ-201 фасовка в ассортименте со склада в Нижнем Новгороде
- Бутилцеллозольв ТУ Продукция завода ПАО «Казаньоргсинтез» ₽50
Твердые нефтепродукты (Воски)| Нектон Сиа
Твердые нефтепродукты (Воски)
27. 03.2012
03.2012
Наряду с парафинами, церезинами, петролатумами и вазелинами к товарным нефтепродуктам на базе твердых углеводородов относят также воски, восковые композиции и составы, широко применяемые в различных областях промышленности. Получают их путем фракционирования парафинов и церезинов, либо обезмасливанием специально подобранного парафиносодержащего сырья, а также путем компаудирования парафинов, церезинов, петролатумов и их смесей с полимерными, смоляными и другими добавками для усиления или придания композициям определенных функциональных свойств.
К числу таких восковых продуктов относят воски для терморегуляторов, для резин, для покрытия сыров, для литья по выплавляемым моделям, для прессового производства, заливочные и прошпарочные массы, защитные восковые и водно-восковые составы различного назначения.
Воски для терморегуляторов
Воски для терморегуляторов — узкие фракции парафинов, церезинов и их смесей — применяют в датчиках температуры, реагирующих на изменение объема воска при его плавлении и кристаллизации.
Синтетический церезин для терморегуляторов (ТУ 38.101261-79) выпускают трех марок. Применяют для терморегуляторов, автоматически регулирующих тепловое состояние двигателей внутреннего сгорания в интервале температур 70…93 градусов Цельсия. Получают путем глубоковакуумного фракционирования синтетического высокоплавкого церезина.
Специальные воски для датчиков температуры (ТУ 38.40102-81) представляю собой специально подобранные смеси твердых углеводородов, получаемых путем глубоковакуумного и холодного фракционирования парафинов и церезинов.
Применяют для терморегуляторов, обеспечивающих автоматическое поддержание заданных температур различных тепловых систем в интервале 15…102 °C.
Воски для резин
Эти воски применяют для защиты резин от отрицательных атмосферных воздействий, светоозонного и теплового старения, в качестве мягчителей резины.
В процессе эксплуатации резин как в статически, так ив динамически нагруженном состоянии воски, выпотевая на поверхность резин, препятствуют разрушающему воздействию света и озона, увеличивая срок годности резиновых изделий.
Защитный воск Паралайт-17 (ТУ 38.1011042-85) — композиция церезина 80 с парафином.
предназначен для защиты резины от атмосферных воздействий при эксплуатации в умеренном и тропическом климате.
Защитный воск ЗВП (ТУ 398.1011290-90) представляет собой фракцию твердых углеводородов, получаемую путем обезмасливания специально подобранной смеси гача и петролатума.
Используется при производстве шин и других резино-технических изделий.
Защитный воск ЗВ-ПФ (ТУ 38.401212-93) — композиция высокоплавкого парафина с церезином.
По эффективности защитного действия и области применения аналогичен защитному воску ЗВП.
Сплав АФ-1 (ТУ 38. 101595-81) представляет собой композицию церезинов нефтяного и синтетического с парафином.
101595-81) представляет собой композицию церезинов нефтяного и синтетического с парафином.
Применяют в производстве резино-технических изделий для защиты их от озонного растрескивания.
Мягчитель ПП для резины (ТУ 38.101225-94) применяют при изготовлении резин для кабельной промышленности.
Получают сплавлением парафина с петролатумом.
Воски для покрытия сыров
Для защиты твердых сычужных сыров от высыхания и плесневения в период их вызревания и хранения применяют глубокоочищенные воски, допущенные к контактам с пищевыми продуктами.
Восковой сплав СДС-13М для сыров (ТУ 38.101225-94), представляет собой композицию глубокоочищенного нефтяного воска с полимерными добавками. В качестве основы применяют воск ВН-2 (ТУ 38.401210-93), который представляет собой фракцию твердых парафиновых углеводородов, выделенных обезмасливанием смеси петролатума и целевого фильтрата, полученного в процессе обезмасливания широкой фракции гача. Для использования в сплаве СДС-13М воск ВН-2 подвергают глубокой очистке.
Для использования в сплаве СДС-13М воск ВН-2 подвергают глубокой очистке.
Модельные восковые составы
Модельные восковые составы применяют на предприятиях машиностроения для точного стального литья. Предложено более 200 рецептур восковых составов для стального литья, но практическое использование нашли лишь единицы. Ниже приведена информация по двум модельным восковым составам, по которым организовано централизованное производство.
Модельный восковой состав МВС-ЗА (ТУ 38.101516-76) представляет собой композицию глубокообезмасленного парафина и высокоплавкого нефтяного церезина с полимерной добавкой.
Предназначен для точного стального литья по выплавляемым моделям.
Модельный восковой состав МВС-15 (ТУ 38.1011044-85) представляет собой композицию высокоплавкого церезина с техническим парафином и полимерной добавкой.
Предназначен для литья по выплавляемым моделям в цехах точного литья заводов тракторного и сельскохозяйственного машиностроения.
Воски для прессового производства
В прессовом производстве машиностроительных предприятий автомобильной и других отраслей промышленности воски используются при изготовлении рабочих моделей и оснастки для штампов и пресс-форм.
Восковой лист ЛК-4С (ТУ 38.101300) представляет собой парафино-церезиновую композицию с полимерными добавками и красителем красного цвета.
Предназначен для изготовления модельной оснастки в прессовом производстве. Выпускают в виде пластин размером 450 х 1000 х 0,8 мм
Восковой лист ЛЖ-4 (ТУ 38.101452-80) предназначен для изготовления рабочих моделей в прессовом производстве. Имитирует листовой металл разной толщины. Выпускают в виде пластин размером 305 х 610 х (0,75-3,0) мм.
С одной стороны лист ЛЖ-4 покрыт невысыхающим клеем, что обеспечивает его прилипание к мастер-модели. Для предотвращения слипания восковые листы перекладывают антиадгезионной силиконизированной бумагой, которую перед использованием листов отделяют без нарушения клеевого слоя.
Применяют в качестве разделительного слоя при изготовлении модельной оснастки прессового производства.
Выпускают в герметично закрывающихся металлических банках.
ВОСКОВЫЕ КОМПОЗИЦИИ И СОСТАВЫ — Справочник химика 21
В состав этих продуктов, как правило, входят сложные петролатумные композиции с большим содержанием маслорастворимых ингибиторов коррозии, ПАВ и углеводородных растворителей. Продукты этой группы образуют на металле эластичные, восковые или мазеобразные мягкие пленки. Основным представителем группы МЛ-1 яьляется продукт мовиль. [c.196]Сорта церезина с высокой температурой плавления можно употреблять для пропитывания форм без смешивания с другими материалами. Однако наиболее целесообразно производить пропитку в восковых композициях, т.
Очень хорошими свойствами обладает восковая композиция, применяемая для механической записи звука при изготовлении граммофонных пластинок. Такая композиция имеет следующий примерный состав (в % по весу) [c.42]
В состав твердых углеводородов входят компоненты, различающиеся по свойствам и, в частности, по температуре плавления. От соотношения углеводородов, плавящихся при разных температурах, зависят многие важнейшие эксплуатационные характеристики нефтяных парафинов, церезинов и восковых композиций, такие, как пластичность, твердость, скорость затвердевания, возможность применения в широком или узком диапазоне температур. [c.33]
Модельный восковой состав МВС-15 (ТУ 38.1011044-85) представляет собой композицию высокоплавкого церезина с техническим парафином и полимерной добавкой. [c.485]
В состав продуктов, как правило, входят сложные мыльно-полимерно-восковые (петролатумные) композиции с большим содержанием маслорастворимых ингибиторов коррозии и других ПАВ и углеводородные растворители, обеспечивающие высокие водовытесняющие, проникающие и пропитывающие свойства ПИНС этой группы. Продукты образуют на металле эластичные, восковые или мазеобразные мягкие пленки. [c.18]
Продукты образуют на металле эластичные, восковые или мазеобразные мягкие пленки. [c.18]
Состав восковой для сыров, ГОСТ 7508—55, представляет собой композицию из парафина (любой марки, кроме спичечного) и петролатума ПК. Состав должен сдаваться окрашенным органическими красителями. Применяется для покрытия сыров с целью предохранения их от усыхания и плесневения. Температура каплепадения, отсутствие запаха и вкуса и ограниченное содержание механических примесей — основные показатели качества воскового состава. [c.376]
Состав восковой № 23, МРТУ 12 Н № 85—64, представляет собой композицию из петролатума по ГОСТ 4096—62 (40 2%) и парафина спичечного по ГОСТ 784—53 (60 2%). Применяется для изготовления восковых моделей при бронзовом литье. Основными показателями качества воскового состава являются температура каплепадения и глубина проникания иглы. [c.385]
Проведенные исследования показывают, что наилучшие варианты применения кремнеорганических соединений все-таки не являются достаточно эффективными, так как не позволяют сохранить гидрофобность в течение 12 часов дождевания. Наилучшие варианты позволяют получить износостойкость только до 6 часов. В связи с этим были проведены изыскания способа повышения эффективности покрытий при дождевании. Проведенные исследования привели в конце концов к применению на силикатном стекле тонких двухслойных покрытий, состоящих из кремнеорганического подслоя и органического наружного слоя. В результате изучения большого количества различных веществ для образования наружного слоя была выбрана композиция из церезина, полиэтилена, нолиизобутилена и лакового бензина (восковой состав). Двухслойное покрытие обеспечивает увеличение износостойкости в два раза. [c.340]
Наилучшие варианты позволяют получить износостойкость только до 6 часов. В связи с этим были проведены изыскания способа повышения эффективности покрытий при дождевании. Проведенные исследования привели в конце концов к применению на силикатном стекле тонких двухслойных покрытий, состоящих из кремнеорганического подслоя и органического наружного слоя. В результате изучения большого количества различных веществ для образования наружного слоя была выбрана композиция из церезина, полиэтилена, нолиизобутилена и лакового бензина (восковой состав). Двухслойное покрытие обеспечивает увеличение износостойкости в два раза. [c.340]
Восковая композиция включает 70 % гаровакса, восковая мон-тановая композиция — 30 % (температура плавления 58,5 °С). Состав пропиточного лака (г) асфальт 50, сера 3,5, льняное масло 150, скипидар 50, бензин 200 смесь нагревают до температуры 120°С и перемешивают. Состав каучукового лака (г) каучук 50, кани-( юль 120, скипидар 300 (готовят так же, как и пропиточный лак).
 [c.267]
[c.267]Хорошо высушенная форма должна иметь совершенно белый цвет и издавать при постукивании специфический звук сухого гипса. Сухая форма быстро воспринимает пропитку и не дает трещин-Гипсовые формы пропитывают расплавленными восковыми составами с температурой плавления от 50 до 125°, в зависимости от состава пропиточной композиции предпочтительны составы, имеющие низкую температуру плавления. Формы, погружаемые в пропиточньг и состав, прогреваются до его температуры при этом состав медленно проникает в поры гипса, вытесняя воздухе. В зависимости от времени выдержки гипсовых форм в составе глубина пропитки может быть различной и считается достаточной при толщине примерно от 2 до 5 мм.- [c.39]
В зависимости от области применения, каждая из которых предъявляет строго определенные требования к качеству потребляемых продуктов, парафины и церезины должны обладать совокупностью эксплуатационных свойств, обусловленной составом и кристаллической структурой твердых углеводородов, на базе которых получены эти продукты. Знание взаимосвязи состава и свойств твердых углеводородов нефти позволяет создавать восковые продукты, представля-юцще собой смеси парафинов и церезинов (взятых в определенных соотношениях), которые обеспечивают высокое качество получаемой композиции. В ряде случаев для улучшения комплекса физико-химических и эксплуатационных свойств восковых композиций в их состав вводят добавки, повышающие стабильность против окисления, улучшающие адгезионные, оптические, физико-химические и другие свойства. [c.57]
Способность кремнийорганических соединений улучшать блеск и кроющую способность восковых композиций используют при приготовлении различных косметических препаратов, в частности губных помад, в состав которых вводится до 6—10% ПМС [12]. Губная помада представляет собой сложную композицию, включающую жиры, масла, воска, эфиры, наполнители, красители и ароматические вещества. Рецептуры отечественных и зарубежных помад примерно одинаковы, однако последние, как правило, содержат силоксаны, которые придают губной помаде специфические потребительские свойства блеск и хорошую кроющую способность. В качестве добавок за рубежом используют жидкие полиметилсилоксаны с вязкостью 200—300 мм с. В Советском Союзе для этих целей применяют полиэтилсилоксановую жидкость ПЭС-5 и полиметилсилоксановую жидкость ПМС-200А. Их вводят в расплав стандартной жировой основы помады в количестве 8—10%. Губная помада с добавкой силоксанов не оказывает на кожу раздражающего и аллергического действия. [c.271]
У одельный восковой состав МВС-ЗА (ТУ 38.101516—76) представляет собой композицию глубокообезмасленного парафина и высокоплавкого нефтяного церезина с полимерной добавкой. [c.484]
В качестве углеводородных загустителей ПИНС могут быть использованы самые разнообразные восковые составы и сплавы— для пищевой промышленности (№ 36, СКФ-15), для флег-матизаторов (СФ-3 и др.), а также воски, используемые в шинной, резинотехнической и других отраслях промышленности ОМСК-1, ОМСК-7, ЦСМ-1, паразон 5Н, ЗВ-1 и др. Технология получения и химический состав твердых углеводородов защитных восков приведены в работах [98]. Показана перспективность получения твердых углеводородов и защитных композиций на их основе из остаточных продуктов переработки западно-сибирских нефтей. Из смесей масла, петролатума, церезина, парафина с добавкой полиизобутилена и окисленного церезина (присадка МНИ-7) вырабатывают защитные смазки ВТВ-1 и ВТВ-2, используемые для защиты от коррозии электроаппаратуры и электрооборудования автомобилей семейства Жигули . Церезин или воск Совцернн с полимерными добавками служат основой для защитных восковых составов изоляционного типа, наносимых из растворителей ПСС-5, ПСС-6, ПЭВ-74. [c.145]
Состай восковой для кондитерских изделий № 36, ГОСТ 7509—55, представляет собой композицию из ларафина (любой марки, кроме спичечного), церезина и вазелинового медицинского масла в соотношении 1 1 2. [c.375]
Показатели Восковой состав № 23. МРТУ 12Н № 85-64 Композиция № 3 для пластилина, МРТУ38-1Г-2-68 Методы испытаний [c.385]
Восковой состав № 23 представляет собой композицию из парафина и пет-ролатума. [c.330]
Озокерпт широко используется и в других отраслях народного хозяйства в парфюмерии применяется специальный церезин для косметических це.ией (ВТУ 499-53) в пищевой прол1Ышленно-сти восковой состав для кондитерских изделий (ГОСТ 7509-55) в резиновой иромышленности озокеритовая композиция (ГОСТ 780-54) как мягчитель при пропитке резиновых тканей. [c.195]
Эффективное масштабируемое производство терапевтических микровезикул, полученных из мезенхимальных стволовых клеток человека
Изготовление массивов цилиндрических микролунок из ПЭГ-гидрогелей с перевернутыми пирамидальными отверстиями
Для настройки массивов цилиндрических микролунок с перевернутыми пирамидальными отверстиями использовались два процесса формования, процесс формования полидиметилсилоксана (PDMS, СИЛИКОНОВЫЙ ЭЛАСТОМЕР SYLGARD® 184, Dow Corning, Мидленд, Мичиган, США) для формирования формы с противоположной структурой PDMS и процесс формования PEG для получения окончательных массивов микролунок гидрогеля PEG.Для первого процесса формования требовалась мастер-форма из кремния (Si), которая имела ту же структуру, что и окончательная конструкция микролунки, и была изготовлена на кремниевой пластине (Waferbiz, Сеул, Корея) с использованием традиционных технологий микротехнологий, таких как мокрое травление Si для отверстия в форме перевернутой пирамиды, сухое травление для цилиндрических структур и процессы распыления и фотолитографии для индивидуальных масок мокрого травления и сухого травления. Изготовленная Si мастер-форма использовалась в первом процессе формования для формирования встречной формы из PDMS, которая имела форму перевернутой Si мастер-формы.Раствор PDMS (соотношение смолы и отвердителя в составе 10: 1) выливали на Si-матрицу и отверждали при 95 ° C в течение 2 часов. Структуры PDMS, отделенные от Si мастер-формы, использовались во втором процессе формования PEG для формирования окончательных массивов микролунок PEG-гидрогеля. Для второго процесса формования ПЭГ использовался раствор ПЭГ, который состоял из фосфатно-солевого буфера (PBS, Lonza, Базель, Швейцария), фотоинициатора (BASF, Людвигсхафен, Германия) и диметакрилата поли (этиленгликоля) 1000 (PEGDMA 1000, Polysciences, Inc., Warrington, PA, USA) в соотношении 10: 0,1: 1 соответственно, выливали на предметное стекло, предварительно покрытое 3- (триметоксисилил) пропилметакрилатом (TMSPMA. Sigma, St. Louis, MO, USA). Раствор PEG штамповался с помощью встречной формы PDMS, и УФ-индуцированное сшивание PEG проводили при 7 Вт / см 2 в течение 65 секунд (Omnicure ® S2000, Excelitas Technologies Corp., Уолтем, Массачусетс, США). После удаления противоформы PDMS изготовленные массивы микролунок с гидрогелем PEG хранили в 70% растворе этанола до использования.
3D-культура hMSC с использованием микролунок
hMSC (PT2501, Lonza, Basel, Switzerland) культивировали в инкубаторе с 5% CO 2 при 37 ° C. Среда для выращивания представляла собой среду Игла, модифицированную Дульбекко с низким содержанием глюкозы (DMEM, Gibco, Гранд-Айленд, Нью-Йорк, США), содержащую 10% эмбриональной телячьей сыворотки (FBS, Hyclone, Logan, UT, США) или FBS без экзосом (System Biosciences, Palo Альто, Калифорния, США) и 1% антибиотиков-антимикотиков (Gibco). FBS, добавленный в нашу культуральную среду hMSC, предварительно фильтровали через 0.22-мкм мембраны для устранения большинства МВ, происходящих из FBS, в анализе. Для посева hMSC в массивы микролунок клетки трипсинизировали с использованием TrypLE Express (Gibco) и подсчитывали с помощью гемоцитометра. Затем их суспендировали в 200 мкл культуральной среды с плотностью 5 × 10 5 клеток / матрица и высевали по каплям в наборы микролунок. ЧМСК объединились в микролунках и агломерировались в виде сфероидов, которые наблюдали с помощью фазово-контрастной микроскопии через 20 мин и 1 день после посева клеток, соответственно.Затем их культивировали при 30 об / мин в орбитальном шейкере в течение 7 дней. Фибробласты мыши (NIh4T3, ATCC, Манассас, Вирджиния, США) также культивировали в среде DMEM с высоким содержанием глюкозы (Gibco), содержащей 10% FBS (Hyclone) и 1% антибиотиков-антимикотиков (Gibco), и использовали для сравнения включений цитокинов в собранных Видеоклипы с таковыми из чМСК.
Выделение MV из культуральной среды hMSC
MV были выделены из собранных сред последовательным центрифугированием при 2500 × g в течение 10 минут для удаления клеточного дебриса / апоптотических тел и при 14000 × g в течение 45 минут при 10 ° C для получения гранул MV 58,59 .
Анализ живых и мертвых hMSC-сфероидов
Жизнеспособность клеток hMSC-сфероидов оценивали с использованием набора LIVE / DEAD Viability / Cytotoxicity Kit (Invitrogen, Carlsbad, CA, USA). Сфероиды hMSC были сформированы, как описано выше, и собраны в D3, D5 и D7 в группе 3D со встряхиванием. Изображения кальцеина AM (живые клетки) и EthD-1 (мертвые клетки) получали с помощью флуоресцентного микроскопа (EVOS, Advanced Microscopy Group, Bothell, WA, США).
Гистологический анализ hMSC-сфероидов
hMSC-сфероиды фиксировали с использованием 4% (мас. / Об.) Параформальдегида (Sigma).После промывки PBS фиксированные образцы помещали в раствор яичного альбумина, разбавленного глицерином, а затем центрифугировали при 3000 об / мин в течение 5 минут с последующим стандартным протоколом заливки в парафин. Срезы погружали в ксилол (Junsei, Tokyo, Japan) для депарафинизации, а затем помещали в этанол с уменьшающейся концентрацией для регидратации. Образцы окрашивали H&E (Sigma) и M&T (Sigma) с использованием стандартных протоколов и наблюдали под микроскопом (EVOS, Advanced Microscopy Group).
Анализ кинетики роста
Количество клеток определяли количественно с использованием набора для количественного анализа ДНК (CyQUANT NF Cell Proliferation Assay Kit, Invitrogen) в соответствии с инструкциями производителя.
Массив кПЦР для характеристики hMSC-сфероидов
Суммарные РНК из hMSC экстрагировали с использованием реагента Trizol (Invitrogen) в соответствии с инструкциями производителя. 84 гена, представляющие характеристики hMSC, были количественно проанализированы с использованием коммерческого набора матриц qPCR (RT 2 Profiler TM PCR array: Human Mesenchymal Stem Cells, Qiagen, Hilden, Germany) в соответствии с инструкциями производителя.
Количественная оценка проточной цитометрии и характеристика размера MV
Метод проточной цитометрии был использован для фенотипирования и подсчета MV, как сообщалось ранее 27,28 . Выделенные MV дважды окрашивали анти-CD105 (AbD Serotec, Кидлингтон, Великобритания) и анти-аннексином V (BD Pharmingen, Сан-Хосе, Калифорния, США) 29 , чтобы подтвердить, что подсчитанные MV произошли от hMSC. Абсолютное количество MV анализировали с использованием как прямого рассеяния (FSC), так и бокового рассеяния (SSC) в логарифмическом режиме (проточный цитометр FACS Verse и программное обеспечение BD FACSuiteTM, BD Biosciences, Сан-Хосе, Калифорния, США).Стандартные шарики 0,22, 0,45, 0,88 и 1,35 мкм (Nano Fluorescent Size Standard, Spherotech, Lake Forest, IL, USA) использовали для оценки размера подсчитываемых MV; в частности, положение затвора R1 было предварительно определено с использованием стандартных шариков 1,35 мкм для обнаружения частиц размером менее 1 мкм. Счетные шарики (7 мкм, CountBrightTM Absolute Counting Beads, Thermo Fisher, Waltham, MA, USA) были использованы и введены в интервал R2 для расчета абсолютных количеств MV, следуя инструкциям производителя (1):
$$ \ frac {{ \ rm {MVs}}} {{\ rm {\ mu}} {\ rm {l}}} = \, (\ frac {total \, events} {назначено \, bead \, count}) \, \ times \, (\ frac {standard \, bead \, event} {volume \, of \, sample}) $$
Подсчитанные MV были нормализованы по количеству клеток в соответствующей культуральной группе, чтобы скорости продукции MV были соизмеримы среди групп с одинаковым стандартом.
Чтобы подтвердить, что собранные MV не были агрегатами белка, а скорее настоящими пузырьками, окруженными липидным двойным слоем, мы исследовали их структурную чувствительность с помощью Triton X-100, который обычно используется для липосомного переваривания 28,31 . Собранные MV погружали в 3% раствор Triton X-100 (Sigma), разбавленный PBS, и измеряли с помощью проточного цитометра, как описано выше.
Количественное определение белка в MV
Содержание белка в MV измеряли с помощью набора Micro BCA Protein Assay Kit (Thermo Scientific) в соответствии с инструкциями производителя.Измеренное количество белка нормализовали к количеству клеток в соответствующей культуральной группе для сравнения всех групп с одним и тем же стандартом.
Определение морфологии и размера MV с использованием TEM
MV фиксировали в течение ночи 2,5% (мас. / Об.) Глутаровым альдегидом (Sigma) в 4% (мас. / Об.) Растворе параформальдегида (Sigma) при 4 ° C. После инкубации в 1% (мас. / Об.) OsO 4 в течение 1 часа образцы дегидратировали в серии разбавителей этанола, пропускали через оксид пропилена и заливали эпоксидной смолой (Epok 812, 02–1001, Окен, Япония. ).Ультратонкие срезы (60 нм) собирали на никелевые сетки 200 меш и окрашивали в течение 10 мин в 1% уранилацетате и цитрате свинца Рейнольдса. Образцы наблюдали с помощью электронного микроскопа Hitachi HT7700 (Токио, Япония) при 80 кВ. Приблизительно 1400 MV были случайным образом выбраны из одиннадцати изображений ПЭМ, и их размеры были измерены с использованием программного обеспечения ImageJ (NIH, Bethesda, MD, USA) для анализа распределения частиц по размерам.
Вестерн-блоттинг-анализ апоптотических телец
Лизаты MV, клеточный дебрис и цельные hMSC для вестерн-блоттинга экстрагировали буфером RIPA, содержащим ингибиторы протеаз и ингибиторы фосфатаз (Roche, Базель, Швейцария).Концентрации белка определяли методом Брэдфорда. Лизаты (25 мкг) подвергали электрофорезу на SDS-полиакриламидных гелях и переносили на мембраны из PVDF (Millipore, Billerica, MA, USA). После блокирования мембраны инкубировали в течение ночи при 4 ° C с первичными антителами: человеческим антитромбоспондином (разведенным до 1: 500, BD Pharmingen) и человеческим анти-C3b (разведенным до 1: 500, BD Pharmingen). Затем мембраны инкубировали со вторичными антителами (разведенными до 1: 5000, Cell Signaling Technology, Danvers, MA, США).Полосы визуализировались с усиленной хемилюминесценцией (Millipore).
Приготовление и лечение IBE
Все эксперименты на животных были одобрены Комитетом по уходу и использованию животных (IACUC) Института биомедицинских исследований Samsung (SBRI, номер разрешения 20160106001) и выполнены в соответствии с требованиями Института ресурсов лабораторных животных (ILAR). ) руководящие указания. Все животные содержались в соответствии с соответствующими законами и институциональными руководящими принципами Исследовательского центра лабораторных животных (LARC; учреждение, одобренное AAALAC International, No.001003) в Медицинском центре Самсунг. Анестезию индуцировали с использованием лицевой маски самцам крыс Sprague-Dawley (SD) (возраст 7-8 недель, 250-300 г) 4% изофлураном (Hana Pharm, Gyeonggi-do, Корея) и поддерживали 1,5% изофлураном в 70 случаях. % N 2 O и 30% O 2 . Температуру тела поддерживали на уровне 37,0–37,5 ° C (ректально) с помощью грелок. Мы вызвали преходящую окклюзию средней мозговой артерии (tMCAo), используя ранее описанный метод внутрипросветной окклюзии сосудов, модифицированный в нашей лаборатории 60 .IBE получали через 3 дня после 90-минутной tMCAo. Ипсилатеральные полушария гомогенизировали в среде DMEM (150 мг / мл) на льду. После центрифугирования при 10000 × g при 4 ° C в течение 10 минут супернатанты собирали и хранили при -70 ° C. hMSC высевали в колбы T75 при плотности 6 × 10 5 клеток на колбу и культивировали при 37 ° C в 5% CO 2 в течение 24 часов. Сохраненный IBE размораживали и центрифугировали при 2500 × g в течение 10 минут для удаления мусора. После 5-кратного разбавления DMEM, IBE центрифугировали при 14000 × g в течение 45 мин при 10 ° C и фильтровали с 0.Фильтр на крышке бутылки 2 мкм для удаления МВ из экстрактов тканей. hMSC подвергались действию приготовленного IBE в течение 24 часов.
Анализы терапевтических включений, содержащихся в MV: цитокины и микро-РНК
Высокопроизводительный скрининг различных включений цитокинов в MV, связанных с терапевтической способностью, был проведен с использованием нескольких наборов набора цитокинов, таких как Proteome Profiler TM Набор человеческих цитокинов XL (R&D Systems, Миннеаполис, Миннесота, США), набор человеческих антител к ангиогенезу (Abcam, Кембридж, Великобритания) и набор человеческих антител против воспаления (Abcam), следуя инструкциям производителя.Чтобы определить ключевые игроки в нейрогенной и / или ангиогенной молекулярной передаче сигналов в MV, мы выполнили анализ экспрессии микро-РНК с использованием количественной ПЦР. Суммарные РНК IBE- и 3D-MV экстрагировали с использованием реагента Trizol (Invitrogen) в соответствии с инструкциями производителя. Уровни экспрессии микро-РНК измеряли с использованием праймеров RT «стебель-петля» и TaqMan PCR Mastermix (Applied Biosystems, Foster City, CA, USA) и нормализовали по уровням miR-16. Праймеры для микро-РНК были включены в тесты TaqMan® MicroRNA Assays (Applied Biosystems), miR-16 (Cat.№ 4427975–000391), miR-134 (№ по каталогу 4427975-001186), miR-137 (№ по каталогу 4427975-001129), miR-184 (№ по каталогу 4427975-000485), miR-210 ( Кат. № 4427975-000512), miR-150 (Кат. № 4427975-462465), miR-155 (Кат. № 4427975-002623) и miR-296 (Кат. № 4427975-002101). Относительные сравнения уровней экспрессии микро-РНК между IBE- и 3D-MV были рассчитаны с использованием сравнительного метода CT (2 — ΔΔCT ).
Анализ культуры HUVEC и образования пробирок
HUVEC были приобретены в ATCC и культивированы в покрытых 1% желатином колбах в M199 с добавлением 20% FBS, 5 ед / мл гепарина (Sigma), 3 нг / мл bFGF (Invitrogen), и антибиотики-антимикотики (Гибко).HUVEC пассажа 3–5 высевали на покрытые матригелем мкл-слайды в M199 с добавлением 1% FBS, 5 Ед / мл гепарина и антибиотиков-антимикотиков. Одновременно HUVEC обрабатывали 3 мкг / мл IBE- или 3D-MV без добавок фактора роста и сравнивали с контрольной (базальная среда) и группами, получавшими VEGF (100 нг / мл, Life Technologies, Карлсбад, Калифорния, США). ). Образование трубок HUVEC наблюдали через 6 часов после обработки, и количество петель, количество ветвей и длины ветвей были количественно определены с помощью программного обеспечения ImageJ.
Первичная культура НСК крыс
SD крысиных эмбрионов получали через 14,5 дней. Кору головного мозга собирали и погружали в DMEM / F12 (Gibco). Менинги удаляли и промывали в DMEM / F12 центрифугированием в течение 5 мин при 500 × g. Осадок коры головного мозга обрабатывали Аккутазой (Biowest, Nuaillé, Франция). Переваривание Accutase нейтрализовали добавлением DMEM / F12, содержащего 1% добавок N2, 20 нг / мл EGF, 20 нг / мл FGF и 1% антибиотиков-антимикотиков. Затем клетки распределяли пипеткой вверх и вниз несколько раз и осаждали центрифугированием.После повторного суспендирования в культуральной среде клетки переносили в колбы T25 и культивировали в инкубаторе при 37 ° C. После 5-7 дней культивирования нейросферы были пассированы для расширения NSC для будущих экспериментов.
Анализ нейрогенной дифференцировки NSC
NSC высевали при 2,5 × 10 5 клеток / мл на 24-луночные планшеты, покрытые 20 мкг / мл поли-D-лизина (Sigma) с использованием DMEM / F12 с добавлением N2 ( Gibco), 20 нг / мл bFGF (Invitrogen), 20 нг / мл EGF (Invitrogen) и антибиотики-антимикотики (Gibco).Для анализа NSC обрабатывали 3 мкг / мл IBE- или 3D-MV без добавок факторов роста. Эти экспериментальные группы сравнивали с контрольной (базальная среда) и группами, обработанными NGF (100 нг / мл, Thermo Scientific). После 4 дней культивирования нейрогенную дифференцировку NSC анализировали с использованием стандартного протокола иммуноцитохимии для кроличьего анти-Ki 67 (разведенное 1:50, Abcam) и мышиное анти-Tuj1 (разведенное 1: 100, Millipore). Вторичные антитела применяли последовательно следующим образом: меченый DyLight анти-кроличий IgG (разведенный 1: 200, 594 нм, Abcam) и меченный DyLight антимышиный IgG (разведенный 1: 200, 488 нм, Vector Laboratories, Burlingame, CA, СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ).После монтажа с использованием Vectashield TM с 1,5 мкг / мл 4′-6 ‘диамидино-2-фенилиндола (DAPI) (Vector Laboratories) образцы были визуализированы с использованием флуоресцентного микроскопа (EVOS, Advanced Microscopy Group), и положительно окрашенные НСК были количественно с использованием программного обеспечения ImageJ.
Статистический анализ
Количественный анализ проводился в более чем трех независимых экспериментах (n ≥ 3), и данные были представлены в виде среднего значения ± стандартная ошибка среднего (SEM). Различия между группами оценивали с помощью одностороннего дисперсионного анализа (ANOVA, апостериорный тест Тьюки) на уровне значимости p <0.001 (***), 0,001
Доступность данных
Все данные, полученные или проанализированные в этом исследовании, включены в эту опубликованную статью (и ее файлы с дополнительной информацией).
Численное моделирование разрушения хрупких твердых тел при нагрузке I / III
В данном исследовании использовался численный анализ для проведения большого числа расчетов численной модели на основе новой модели полукруглого изгиба (SCB).Вместо существующих методов численных расчетов ( J -интегральный), для расчета параметров трещин смешанного режима использовался метод M -integral ( K I , K II и K III ), исследовать влияние геометрии и параметров материала на поведение разрушения при смешанном режиме нагружения I / III и прогнозировать траекторию разрушения. Результаты показали, что ограничения в области применения параметра смешения M e в предыдущих исследованиях могут быть в определенной степени преодолены.Диапазон M e был установлен для различных коэффициентов Пуассона и может использоваться в качестве эталона для реальных испытаний материалов. Траектория моделирования хорошо согласуется с экспериментально полученной траекторией разрушения, и предложенный метод может быть использован для моделирования траектории разрыва при смешанном режиме нагружения I / III.
1. Введение
Механика разрушения фокусируется на механических свойствах материалов и конструкций с трещинами. Трещины могут возникать в самом материале или возникать во время производства, и наличие и рост этих трещин снижает несущую способность конструкции или даже вызывает ее разрушение.
Полукруглый изгиб (SCB) — это классический тест на механику разрушения, который широко используется для исследования твердых материалов из-за присущих ему благоприятных свойств, таких как простота, минимальные требования к механической обработке, удобство тестирования и легкость достижения разрушения при растяжении, как сообщает Махинда и Куруппу [1]. Образец SCB был предложен Чонгом [2–4] и постепенно улучшался. Кроме того, он широко использовался для определения параметра разрушения K IC , и его потенциальное применение в испытании на разрушение в смешанном режиме было исследовано [5–9].Исследование образца SCB в основном включает испытания на вязкость разрушения и определение параметров разрушения [10–28], обсуждение и пересмотр ограничений критериев расширения [11, 13, 16, 25, 29, 30] и численное прогнозирование разрушения. путь [14, 20, 23, 27, 31–33]. Из-за ограничений формы образца, вышеупомянутые исследования были сосредоточены на параметрах разрушения режима I, режима II и смешанного режима I / II. Режим сдвига вне плоскости (режим III) не был так тщательно исследован, хотя он существует в реальных ситуациях.Кроме того, существующие методы испытаний сильно зависят от машины и испытательного оборудования и в основном ориентированы на металлические материалы, керамику и другие искусственные материалы [34–38].
Поскольку инженерные аварии, такие как разрушение горных пород и разрыв покрытия, происходят часто, исследование режима III вызвало большой интерес. Более того, наблюдения показали, что плоские трещины имеют тенденцию переориентироваться в наклонные плоскости во время распространения и могут расти в условиях смешанного режима I / III [39, 40].Поэтому были предложены методы испытаний и образцы в вышеупомянутых условиях для горных пород, бетона, асфальта и других материалов. Берто и др. применили критерий плотности энергии деформации (SED) к чистым режимам и смешанным режимам I / III условий нагружения [41–45]. Алиха и др. [46–49] предложили новую конфигурацию испытаний, названную ENDB образцом, для исследования поведения разрушения в смешанном режиме I / III. Кроме того, эта конфигурация может обеспечить произвольную композицию режима I и режима III.Примечательно, однако, что в эксперименте необходимо контролировать больше переменных. Linul et al. [50, 52] провели эксперименты с пеноматериалами и получили данные о вязкости разрушения для смешанных режимов I / II и I / III. Впоследствии Пирмохаммад [53] предложил новый тип образца SCB, поведение разрушения которого в смешанном режиме I / III легче исследовать, используя предложенные образцы с наклонной трещиной, и дополнительно исследовал поведение разрушения асфальтобетона при низкой температуре.
В предыдущих исследованиях [49, 53] изменение геометрических параметров и параметров материала оказало значительное влияние на смешанный режим I / II нагружения образцов SCB.Однако лишь несколько исследований изучали, влияют ли вышеупомянутые условия на поведение разрушения в режиме I / III. В этом исследовании был использован относительно точный метод численного анализа для исследования возможности реализации смешанного режима I / III нагружения и влияния геометрических параметров образца и коэффициента Пуассона на эти условия нагружения. Наконец, траектория трещины была смоделирована и проанализирована на основе параметров трещины.
2. Трехмерный численный анализ
В предыдущих численных исследованиях [41, 53, 54] параметры трещин рассчитывались напрямую с использованием интегрального метода J в программе Abaqus.Однако, по словам Ли, интеграл J больше подходит для задач разрушения чистой моды I [55]. Для разрушения в смешанном режиме наиболее точным методом расчета параметров трещины является уравнение эквивалентной области для интеграла взаимодействия ( M -интеграл) [56, 57]. Макроструктурная модель SCB была создана в Abaqus; Затем сетка была перестроена с использованием технологии адаптивной сетки в franc3D, и была вставлена нижняя наклонная трещина. Наконец, с помощью интеграла M были получены различные значения K I , K II и K III .
Принципиальная схема полукруглой формы показана на рисунке 1; Радиус образца SCB ( R ), толщина образца ( t ) и половина диапазона нагрузки ( S ) в испытании SCB были определены путем моделирования в Abaqus. На рисунке 2 (а) показана сеточная модель Abaqus с ячейками C3D10. На рисунке 2 (b) показана сеточная модель после определения длины трещины ( a ) и угла трещины ( α ). На рисунке 3 показано распределение элементов и типы вершин трещин.Отдельный элемент был вставлен в вершину трещины, и интегральная оценка консервации была проведена вокруг двух колец элементов на фронте трещины. Для уменьшения локальных дискретных ошибок использовались внутреннее кольцо 15-узлового сингулярного клиновидного элемента и внешнее кольцо 20-узлового шестигранного элемента с симметричными сетками. Примечательно, что параметры разрушения ( K I , K II , и K III ), рассчитанные в зоне контакта с поверхностью, не являются надежными, поскольку условие локальной плоской деформации больше не поддерживается. .Элемент на вершине трещины может быть сильно деформирован, как показано на рисунке 2 (c), и симметрия сетки может быть потеряна. Кроме того, на рис. 2 (c) показано, что переходная область в вершине трещины имеет пирамидальные элементы, а глобальная область представляет собой элемент тетраэдрического типа.
В созданных моделях радиус образца SCB ( R ), толщина образца ( т ) и приложенная нагрузка ( P ) являются инвариантами, соответственно, 75 мм, 32 мм и 40,3437 кН.Геометрические параметры и параметры материала, такие как отношение половины пролета точки нагружения ( S / R ), отношение длины трещины ( a / R ), угол трещины ( α ) и коэффициент Пуассона, варьировались в следующие диапазоны: S / R = {0,4, 0,6, 0,8}, a / R = {0,2, 0,26, 0,3}, α = {0, 5, 10, 15, 20 , 25, 30, 35, 40, 45, 48, 50, 52, 55, 60} и = {0,1, 0,2, 0,3, 0,4}. Когда α составляет 0 °, это разрушение чистой моды I.При увеличении α можно получить трещину смешанного типа I / III.
Параметры трещины ( K I , K II и K III ) могут быть непосредственно извлечены с помощью следующих выражений. Для линейного анализа мы можем добавить два действительных решения, и результатом будет действительное решение, как показано ниже:
Давайте рассмотрим решения с угловой меткой (1) как результаты Abaqus и решения с угловой меткой (2) как решения, которые мы можем выбрать.Их можно подставить в выражение для интеграла J следующим образом:
Согласно теореме взаимности Бетти, имеет место следующее соотношение:
Собрав члены, можно получить следующее соотношение: с
Кончик трещины Скорость высвобождения энергии может быть определена из интеграла закрытия трещины Ирвина для мелкомасштабной текучести в предположении условий плоской деформации следующим образом:
Подставляя в выражение для скорости выделения энергии, мы получаем следующее соотношение:
Приравнивая два определения для M -Integral, мы можем получить следующее соотношение: где
Мы использовали результаты Abaqus для решения (1) и выбрали три простых вспомогательных решения (2a), (2b) и (2c).В таблице 1 представлены значения асимптотических решений для трещин чистой моды.
| ||||||||||||||||||||||||||||
Из аналитических выражений для полей фронта трещины можно получить следующее:
Подставив значение K (2) , представленный в Таблице 1, в уравнении (9), мы можем получить три уравнения для неизвестных K (1) , как показано ниже: где — коэффициент Пуассона, — интегральная кривая от конца трещина, — дельта Кронекера, q — функция, которая равна единице на вершине трещины и нулю на границе области интегрирования и может интерпретироваться как виртуальное расширение трещины, является значением функции q вдоль фронта трещины, а L — длина цилиндрической области вдоль фронта трещины.это решение, предоставленное результатами Abaqus.
K I , K II и K III могут быть выражены безразмерными параметрами Y I , Y II и Y III , а именно:
Для проверки того, что метод численного моделирования может точно реализовать ГРП в смешанном режиме I / III, K I , K II и K III вершины трещины по толщине образца были приняты после обработки нормализации, как показано на рисунке 4.Здесь K n ( n = I, II, III) — параметр разрушения, а K Im — максимальное значение режима I. Отношение длин трещин ( a / R ) модели составляет 0,26, половина отношения точки нагружения ( S / R ) составляет 0,8, а угол трещины ( α, ) составляет 0 °, 20 °, 40 ° и 60 °. ° соответственно.
Рисунок 4 (а) показывает, что, когда α было 0 °, разрушение произошло в условиях чистой моды I; K I по направлению толщины образца оставалась примерно неизменной в определенном диапазоне (0.1 < z / t <0,9) и уменьшилась на свободной поверхности, что согласуется с результатами численного моделирования, представленными Алихой и Сагхафи [54]. Кроме того, значение z / t было принято равным 0,5 для значения K I образца.
Рисунки 4 (b) –4 (d) показывают, что по мере увеличения α эффекты мод I и III преобладали в определенном диапазоне (0,2 < z / t <0,8) вдоль направления толщины. образца.Доля моды II была мала, и ею можно было пренебречь, когда z / t было 0,5, что согласуется с ранее опубликованными результатами численного анализа [46–49]. Учитывая, что расчет зоны контакта поверхности K I , K II и K III не является надежным, и согласно традиционному определению поля сингулярных напряжений вблизи свободной поверхности, Широко признано, что поле напряжений в вершине трещины отличается, как это обсуждали Бажант и Эстенсоро [58].Следовательно, K I , K II и K III близко к свободной поверхности не следует рассматривать, и этот численный метод действителен и осуществим для испытания разрушения смешанного режима I / III. . Примечательно, что при смешанном режиме нагружения I / III вершина трещины по толщине образца полностью соответствует точке, где z / t составляет 0,5. Поэтому в последующих расчетах эта точка рассматривалась как значение K I , K II и K III образца.
Параметр смешивания M e может использоваться для описания относительных вкладов режима I и режима III, как выражено уравнением (15). В условиях чистой моды I значение M e равно единице и уменьшается по мере увеличения вклада режима III:
Для количественной оценки роли режима I и режима III в численном моделировании , мы предлагаем подставить уравнение (16) вместо уравнения (15), а параметр | M e | может изменяться от 0 до 1, что соответствует условиям нагрузки чистого режима III и режима I соответственно:
3.Результаты и обсуждение
3.1. Результаты численных расчетов и анализ
Отношение длины трещины ( a / R ) осталось неизменным на уровне 0,267 и a = 20 мм. Влияние угла трещины ( α ), половинного отношения пролета точки нагружения ( S / R ) и коэффициента Пуассона для Y I , Y III и | M e | были исследованы. На рисунках 5 и 6 показаны варианты Y I , Y III и | M e |, с углом наклона трещины ( α ) для различных значений S / R и в образце SCB.
Из анализа изображений были сделаны следующие выводы: (a) Y III оставалось равным нулю только тогда, когда α было 0 °. Для того же α , увеличение S / R и продвижение Y I . Но для Y III увеличение S / R привело к его уменьшению и увеличению соответственно. (Б) Для чистой моды I ( α = 0 °) увеличение способствовал вкладу режима I, и скорость роста имела тенденцию к увеличению.(c) С увеличением α , после входа в смешанный режим I / III нагружения, значения Y I постепенно уменьшались, и | Y III | сначала увеличился, а затем уменьшился. Кроме того, α , соответствующий | Y III | max медленно увеличивался с увеличением S / R . Чувствительность Y I к изменениям уменьшалась по мере увеличения α , а чувствительность Y III была максимальной при | Y III | макс .(d) Увеличение стоимости S / R оказало лишь незначительное влияние на | M e | значения, когда было 0,1 образцов SCB. При изменении диапазона от 0,2 до 0,4 увеличение S / R , очевидно, привело к уменьшению | M e |. Кроме того, по мере увеличения α нисходящий тренд стал более очевидным.
Половина коэффициента пролета точки нагружения ( S / R ) осталась неизменной и составила 0.8, S = 60 мм, и влияние угла трещины ( α ), отношения длины трещины ( a / R ) и коэффициента Пуассона () на Y I , Y III и | M e | были исследованы. На рисунках 7 и 8 показаны варианты Y I , Y III и | M e | с углом наклона трещины ( α ) для различных значений a / R и в образце SCB.На рисунке 9 показан | M e | мин. получается с заменой на / R и.
На основе анализа изображений были сделаны следующие выводы: (a) Для того же α увеличение на / R и привело к значениям Y I . Но для Y III увеличение на / R и привело к его уменьшению и увеличению соответственно.(b) Значение α , соответствующее | Y III | макс. стабилизировано примерно при 49 °. (C) Для значений | M e |, увеличение на / R привело к продвижению M e ; когда значение a / R было относительно небольшим, влияние на M e было меньше при том же α .(d) Значение α было 60 °, что соответствует | M e | мин. всех образцов SCB. Как и , / R стал меньше, тем меньше | M e | мин. Было получено значение , минимальное значение составило 0,39. В этом случае вклад модели III был явно выше по сравнению с вкладом модели I. Кроме того, для меньшего / R оказал лишь незначительное влияние на | M e | мин ; однако, когда / R был больше, | M e | мин был более чувствителен к вариациям.
Следует отметить, что правило, описанное в предыдущем разделе, не описывается.
3.2. Моделирование траектории разрушения
С использованием K I , K II и K III значений вершины трещины и критерия максимального растягивающего напряжения (MTS), три группы моделей ( S / R = 0,8; a / R = 0,267; α = 0 °, 30 °, 45 ° и 60 °) были получены для пути разрушения и роста трещины, как показано на рисунках 10 и 11.Для облегчения сравнения режим отказа после переворота изображения по горизонтали хорошо согласуется с предыдущими результатами [53].
При смешанном режиме нагружения I / III морфология роста поверхности трещины отличалась от плоского разрушения. Рисунок 12 (а) показывает, что, когда возникла трещина в плоскости (режим I, режим II и смешанный режим I / II), поверхность трещины оставалась в той же плоскости, несмотря на определенный прогиб. Однако на Рисунке 12 (b) показано, что, когда произошло разрушение в смешанном режиме I / III, поверхность трещины была скручена и отклонена от направления поверхности трещины, образуя угол, называемый углом перегиба.
Учитывая направление угла излома трещины (), компоненты нормального напряжения и касательного напряжения на новой плоскости могут быть получены путем преобразования координат следующим образом: где x , y и z — это исходные оси и x ′, y ′ и z ′ являются осями плоскости трещины после прогиба. Коэффициент интенсивности преобразованного напряжения может быть выражен следующим образом:
Соотношение между скоростью высвобождения механической энергии и углом выражается следующим образом:
Путем совмещения вкладов режима III можно выразить соответствующий коэффициент интенсивности преобразованного напряжения. следующее.
Подставляя уравнение (18) в уравнение (19), можно получить выражение скорости высвобождения механической энергии, изменяющейся в зависимости от угла, для смешанного режима I / III. На рисунке 13 показана нормализация скорости выделения механической энергии вместе с изменением угла перегиба трещины ().
Как видно на графике, в случае чистого режима I трещиноватость ( K III / K I = 0) стала максимальной при угле перегиба () 0 °.Следовательно, трещина чистой моды I распространялась в направлении оригинала, когда угол перегиба составлял 0 °. При добавлении наложения вклада моды III ( K III / K I = 0,25), как показано на рисунке 13, угол излома трещины отклонился от исходной плоскости трещины на угол, который может быть рассматривается как поправка за счет режима III. Наконец, трещина двигалась вверх в направлении, перпендикулярном максимальному главному растяжению, и переходила от смешанного режима I / III к чистому режиму I.Результаты численного моделирования, представленные на рисунке 10, показывают, что поверхность растяжения постепенно возвращается в плоскость с изогнутой поверхности вблизи точки нагружения и, наконец, разрушается вблизи точки нагружения. Это явление согласуется с тем, что поверхность излома делится на три области [53]. В таблице 2 представлен угол перегиба от 0,2 до 0,8 на фронте трещины при различных α , извлеченных из Franc3D. Параметры образца SCB: S / R = 0.8, a / R = 0,267 и = 0,2. Кроме того, было замечено, что угол перегиба трещины связан с α , а большее значение α привело к большему углу перегиба.