Лигнификация, как способ приспособления растений к действию абиотических стрессовых факторов — UrFU Portal Investigación
Масштабное отчуждение сельскохозяйственных земель, их загрязнение ионами тяжелых металлов (ТМ), и как следствие, снижение урожайности и продуктивности растений – глобальные проблемы, которые необходимо решать для устойчивого развития биосферы и человечества. Распространение тяжелых металлов в среде и засоление – это неизбежный результат производства самых разных товаров бытового и производственного назначения. Концентрации поллютантов особенно высоки в местах добычи руд, металлургических производств, сельскохозяйственного производства. На загрязненных грунтах таких территорий растения вынуждены приспосабливаться к стрессорам.
При действии стрессоров в растении происходит образование АФК, что приводит к развитию окислительного взрыва. Длительное действие стрессора может сопровождаться подавлением роста растений и нарушением их жизненного цикла вследствие перераспределения ресурсов на образование компонентов антиоксидантной защиты, таких как антиоксидантное ферменты b низкомолекулярные антиоксиданты.
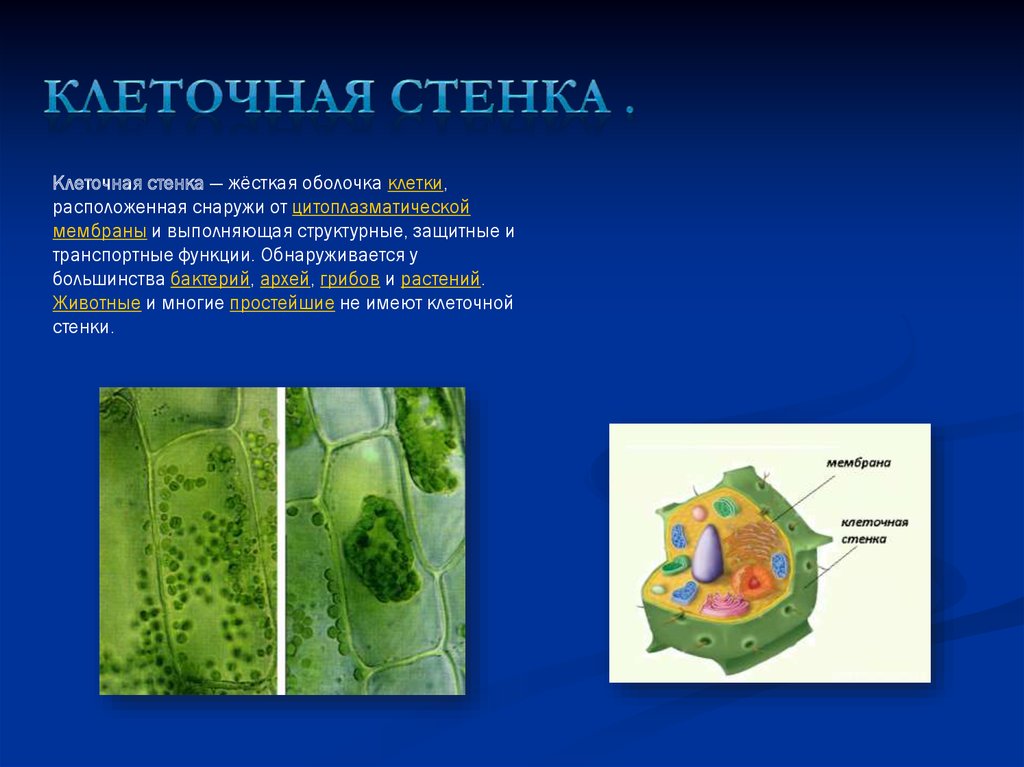
Наряду с активацией антиоксидантных систем одним из механизмов акклимации растений к избытку поллютантов в среде является их иммобилизация в клеточных стенках растений. Клеточная стенка (КС) – сложный компартмент растительной клетки, благодаря катионно-обменным свойствам способен связывать и ограничивать транспорт ионов, в том числе, ТМ, что приводит к снижению их концентрации в надземных частях растений. Таким образом, КС может выполнять в растении барьерные функции.
В ответ на действие неблагоприятных факторов внешней среды во многих растениях стимулируется биосинтез фенолов как антиоксидантов и предщественников лигнина. Эти реакции являются следствием изменения экспрессии генов PAL, C4H, 4CL и др. (Qingquan, 2018). Считается, что отложение лигнина повышает толерантность растений к ТМ за счет ограничения их подвижности в апопласте (Zeng, 2016).
Таким образом, понимание регуляции процесса лигнификации как приспособления растений к стрессовым воздействиям на разных уровнях от молекулярного (изменение экспрессии генов, кодирующих ферменты фенилпропаноидного пути) до морфофизиологического (биосинтеза лигнина в разных тканях и органах) остается актуальной научной задачей.

Цели исследования – изучение физиолого-биохимических, молекулярно-генетических и анатомо-морфологических (лигнификации клеточных стенок) механизмов устойчивости растений табака и циннии в условиях стресса, вызванного ионами меди засолением.
Задачи:
1. Определить уровень стресса в тканях корня и побега растений в условиях избытка ионов меди в среде и при засолении.
2. Оценить содержание фенолов, количество и композицию лигнина в растениях при стрессе.
3. Определить уровень экспрессии генов, кодирующих ферменты раннего этапа фенилпропаноидного пути метаболизма (ФП) и биосинтеза лигнина.
Методологической основой исследования будет комплексный подход к изучению роли лигнификации и ее регуляции в условиях абиотического стресса у растений, предусматривающий гистохимические, физиолого-биохимические и молекулярно-генетические методы исследований.
 Будет определено содержание маркеров окислительного стресса, активность ферментов антиоксидантной защиты, их локализация в тканях корня и побега, лигнификация КС, уровень экспрессии генов, кодирующих ферменты фенилпропаноидного пути и биосинтеза лигнина, а также генов ТФ, регулирующих эти процессы.
Будет определено содержание маркеров окислительного стресса, активность ферментов антиоксидантной защиты, их локализация в тканях корня и побега, лигнификация КС, уровень экспрессии генов, кодирующих ферменты фенилпропаноидного пути и биосинтеза лигнина, а также генов ТФ, регулирующих эти процессы.Научная новизна планируемого исследования. Впервые в длительном эксперименте будет изучена лигнификация тканей корня и стебля модельных растений цинии и табака в условиях действия стрессовых факторов – ионов меди и засоления. Впервые будет показано изменение экспрессии генов, кодирующих ферменты биосинтеза фенолов и лигнина (PAL, C4H, 4CL, LAC, PRX), а также белков-трансфакторов (ERF1, ERF2, MYB1 и MYB2), участвующих в регуляции экспрессии указанных генов в условиях стресса, вызванного избытком ионов меди в среде или засолением. Впервые будет определена локализация ферментов антиоксидантной защиты в разновозрастных тканях корня и стебля модельных растений, дана характеристика физиолого-биохимического статуса растений в условиях стресса и в период восстановления после стресса.
 Будут установлены коррелятивные связи между экспрессией генов, количеством лигнина, фенолов, и маркерами стресса у растений.
Будут установлены коррелятивные связи между экспрессией генов, количеством лигнина, фенолов, и маркерами стресса у растений. Ожидаемые результаты:
В 2022 году:
Будут изучены ростовые и анатомические характеристики корня и стебля модельных растений; будут выявлены те из них, которые могут рассматриваться в качестве маркеров стресса и ответных реакций на стресс, вызванный абиотическими факторами.
Будет определено общее содержание фенольных соединений в органах растений в норме и при стрессе.
Будет выявлена гистохимическая локализация пероксидаз III класса, определена их тотальная активность, количество и активность изоформ в разных органах в норме и при стрессе.
Будет проведена статистическая обработка материалов исследования с целью поиска корреляций между активностью пероксидаз III класса, содержанием лигнина и фенолов, ростом растений при действии абиотических стрессоров.
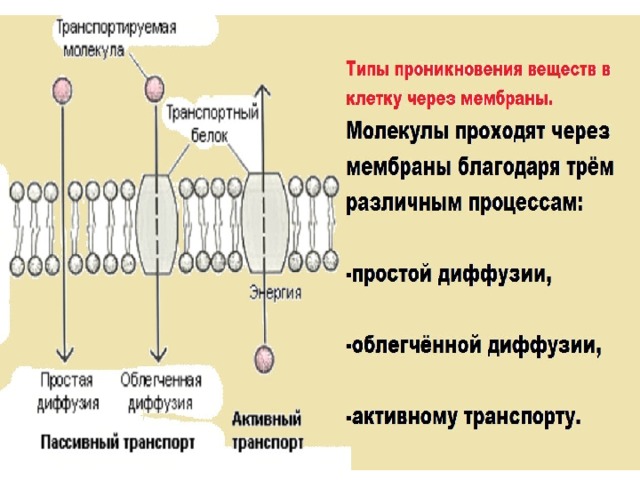
В 2023 году:
Будет определено содержание и количество продуктов фенилпропаноидного пути.
Будет выполнен анализ экспрессии генов, кодирующих ферменты фенилпропаноидного пути и трансфакторы семейства ERF и MYB, участвующие в регуляции экспрессии генов биосинтеза лигнина и его предшественников. Будет проведен сравнительный анализ содержания фенолов и лигнина в тканях и экспрессии отдельных генов, кодирующих ферменты фенилпропаноидного пути.
На основе проведенного анализа будут выявлены гены-маркеры устойчивости растений к действию абиотических факторов среды, таких как ионы ТМ и засоление.
Будет определено содержание ионов меди в тканях корней, стебля и побега растений, рассчитаны корреляции между лигнификацией, содержанием фенолов, активностью пероксидаз и количеством ионов меди.
Будет проведена статистическая обработка материалов исследования, предложена модель регуляции лигнификации тканей растений при стрессе.
| Estado | Activo |
|---|---|
| Fecha de inicio / finalización efectiva | 01/01/2022 → 31/12/2023 |
Ver todo
Ver menos
- 34.
 31.27
31.27 - 34.31.00
 31.27
31.27- Kuibyshev Research Division
Copper Stress Enhances the Lignification of Axial Organs in Zinnia elegans
Tugbaeva, A., Ermoshin, A., Wuriyanghan, H., Maleva, M., Borisova, G. & Kiseleva, I., jun. 2022, En: Horticulturae. 8, 6, 558.
Resultado de la investigación: Article › revisión exhaustiva
Acceso abierto
1 Cita (Scopus)
The Role of Secondary Metabolites and Bark Chemistry in Shaping Diversity and Abundance of Epiphytic Lichens
Paukov, A.
, Teptina, A., Ermoshin, A., Kruglova, E. & Shabardina, L., 13 may. 2022, En: Frontiers in Forests and Global Change. 5, 828211.Resultado de la investigación: Article › revisión exhaustiva
Acceso abierto
 , Teptina, A., Ermoshin, A., Kruglova, E. & Shabardina, L., 13 may. 2022, En: Frontiers in Forests and Global Change. 5, 828211.
, Teptina, A., Ermoshin, A., Kruglova, E. & Shabardina, L., 13 may. 2022, En: Frontiers in Forests and Global Change. 5, 828211.Лигнификация
Реакция на лигнин Лигнификация клеточных оболочек древесины свидетельствует о прошедшем этапе вызревания побега и его подготовке к зиме. Существует две большие группы лигнина: компонент «Ф» — фло-роглюциновая реакция, компонент «М» — реакция Меуле с перманганатом. При одревеснении побегов и подготовке их к зиме появляется сначала лигнин «М», а потом лигнин «Ф». Таким образом, положительная реакция только с перманганатом свидетельствует о неполном одревеснении побега.[ …]
Существует две большие группы лигнина: компонент «Ф» — фло-роглюциновая реакция, компонент «М» — реакция Меуле с перманганатом. При одревеснении побегов и подготовке их к зиме появляется сначала лигнин «М», а потом лигнин «Ф». Таким образом, положительная реакция только с перманганатом свидетельствует о неполном одревеснении побега.[ …]
Роль сахаров в процессе лигнификации выяснилась в последние годы. Исследованиями с меченой глюкозой древесины сосны Pinus strobus и Eucalyptus nitens /27, 28/ был подтвержден механизм образования лигнина через шикимовую (см. рис. 2) и префеновую (рис. 3) кислоты. Аналогичные эксперименты с меченой шикимовой кислотой подтвердили, что этот механизм является при лигнификации решающим. Ацетатный механизм (см. рис. 1) в процессе лигнификации не играет роли. Шикимовая кислота чрезвычайно распространена в высших растениях,и в настоящее время установлено, что шикимовая и префеновая кислоты управляют синтезом ароматических соединений в микроорганизмах и растениях. Далее было установлено, что ключевую роль при образовании лигнина играют аромаэтнеские аминокио-лоты — фенилаланин и тирозин, которые являются промежуточными продуктами метаболизма фенилпропаноидов (рис. 5). Опыты с мечеными фенилаланином и тирозином однозначно доказали, что все растения обладают способностью инкорпорировать фенилаланин в ароматической части лигнина. С другой стороны, тирозин не играет такой универсальной роли в процессе лигнификации растений. Поэтому предпочтительным оказывается механизм с участием фенилаланина, превращаемого дезаминированием в коричную кислоту (рис. 6), а не сходный механизм перехода тирозина в n-кумаровую кислоту.[ …]
Далее было установлено, что ключевую роль при образовании лигнина играют аромаэтнеские аминокио-лоты — фенилаланин и тирозин, которые являются промежуточными продуктами метаболизма фенилпропаноидов (рис. 5). Опыты с мечеными фенилаланином и тирозином однозначно доказали, что все растения обладают способностью инкорпорировать фенилаланин в ароматической части лигнина. С другой стороны, тирозин не играет такой универсальной роли в процессе лигнификации растений. Поэтому предпочтительным оказывается механизм с участием фенилаланина, превращаемого дезаминированием в коричную кислоту (рис. 6), а не сходный механизм перехода тирозина в n-кумаровую кислоту.[ …]
Вентиг [368] для оценки степени лигнификации использовал хлорирование протолигнина с определением хлорного числа, т. е. процента увеличения в весе древесной пробы при хлорировании. В то время как хлорное число солянокислотного лигнина оказалось равным 150, хлорное число еловой древесины было 47,5, сосны 47, березы 42, дуба 43,4, бука 35—39. Хлорлигнин был выделен экстракцией этанолом и содержал 29,6% хлора. Он растворялся в разбавленной щелочи, после чего до 75% хлора отщеплялось. Из этого раствора лигнин осаждался известковым молоком или хлористым кальцием в виде нерастворимой кальциевой соли, которая затрудняет отбелку крафтцеллюлозы и натронной целлюлозы растворами гиплохлорита кальция.[ …]
Хлорлигнин был выделен экстракцией этанолом и содержал 29,6% хлора. Он растворялся в разбавленной щелочи, после чего до 75% хлора отщеплялось. Из этого раствора лигнин осаждался известковым молоком или хлористым кальцием в виде нерастворимой кальциевой соли, которая затрудняет отбелку крафтцеллюлозы и натронной целлюлозы растворами гиплохлорита кальция.[ …]
В результате своих исследований по лигнификации ячменя и овса Филлипс [634] показал, что с увеличением содержания лигнина абсолютное количество целлюлозы и пентозанов скорее увеличивается, чем уменьшается и что нет указаний на то, что растение синтезирует лигнин из этих компонентов. По мнению Филлипса, лигнин образуется или из глюкозы, или из фруктозы, но в последнем случае, еще не известно, только ли фруктоза включается в синтез. Одной из первых ступеней в образовании лигнина является метилирование гидроксильных групп углеводов. По мнению Брауна и Филлипса [649], в молодом овсе содержатся относительно большие количества частично метилированных соединений, которые постепенно исчезают по мере увеличения содержания лигнина. Они не считали конифе-риловый спирт или альдегид промежуточным продуктом в фотохимическом синтезе лигнина.[ …]
Они не считали конифе-риловый спирт или альдегид промежуточным продуктом в фотохимическом синтезе лигнина.[ …]
Шуба» из активных форм кислорода на поверхности молодых корней не опасна для них, так как эта защитная система вынесена на поверхность клеток, а время жизни токсических продуктов невелико. Любопытно, что свойство эпителиальных растительных клеток генерировать активированный кислород очень напоминает поведение полиморфноядерных лейкоцитов (компонентов иммунной системы животных) во время фагоцитоза. Последние образуют токсические кислородсодержащие продукты и используют их для убивания бактерий. Активные формы кислорода играют важную роль в реакции сверхчувствительности растительной клетки (Doke, 1983а, 1983b). Они могут быть индукторами синтеза гормонов «тревоги» (алармонов), способных экспрессировать гены (Rogers et al., 1988).[ …]
Все вышеуказанное включается в понятие «вызревание побегов», под чем подразумевается не только лигнификация, но и опробковение покровов побега, заложение верхушечной почки, физиологобиохимические и структурные изменения биоколлоидов протоплазмы и накопление в клетках запасных веществ. [ …]
[ …]
Слои ксилемы лигнифицируются (древеснеют) и накапливаются в дереве, образуя основную часть ствола. В флоэме лигнификация либо вовсе отсутствует, либо происходит только в некоторых клетках; флоэма образуется из года в год снаружи слоя камбия. По мере образования новой флоэмы слой ее отодвигается все дальше и дальше к наружной стороне ствола и в конце концов образует кору.[ …]
Отложение лигнина в оболочке может происходить лишь при жизни клетки, т. е. в присутствии протопласта. Но не следует думать, что лигнификация оболочки приводит к отмиранию клетки. В древесине имеющиеся немногие живые паренхимные клетки, наполненные запасными веществами и активно участвующие в метаболизме (обмене веществ), имеют, как правило, одревесневшие оболочки. Синтез лигнина происходит в цитоплазме, а исходные вещества поступают из камбия [22]. Образование и отложение лигнина связано с нарушением работы дыхательного аппарата клетки. Процесс одревеснения сопровождается усилением дыхания (усилением поглощения кислорода). Лигнин — продукт окисления ароматических веществ, служащих катализаторами при окислительно-восстановительных процессах, со ставляющих дыхательный цикл.[ …]
Лигнин — продукт окисления ароматических веществ, служащих катализаторами при окислительно-восстановительных процессах, со ставляющих дыхательный цикл.[ …]
Ботаники широко пользовались флороглюциновой реакцией, особенно при изучении сезонного роста деревьев [77], для определения того, когда происходит лигнификация. Имеется, однако, очень мало доказательств того, что физиологический процесс лигнификации вызван первоначально присутствием вещества, известного химикам в качестве лигнина.[ …]
Значительное внимание уделялось природе ферментной системы, катализирующей полимеризацию при образовании лигнина. Модельные опыты с целым рядом фенольных производных и чистыми ферментами указывают на то, что главную роль в процессе лигнификации играет пероксидаза. Сравнительно широкое распространение ее в высших растениях подкрепляет это предположение /37/. Все работы в области лигнификации проводились на модельных системах in vitro.[ …]
Лигнификация не ограничивается клеточными стенками древесины и однолетних растений, но также наблюдается в фруктовых косточках, лубе, сердцевине и пробковых клетках; она происходит только при участии живой протоплазмы. После лигнификации клетка постепенно отмирает. Лигнин встречается в растении не в форме ориентированных пластинок или волокон, а как стройная разветвленная система. Изолированный в нерастворимой форме он еще сохраняет морфологическую структуру волокна [15]. Морфология лигнина выявляет неравномерную структуру без пространственной решетки; лигнин должен быть аморфным высокополимерным веществом. Его пермутоидные свойства объясняют его огромную реакционную способность и тот факт, что в нерастворен-ном состоянии он реагирует легко и совершенно однообразно [47]. Он никогда не встречается в природе один, а всегда в сочетании с целлюлозой или другими углеводами.[ …]
После лигнификации клетка постепенно отмирает. Лигнин встречается в растении не в форме ориентированных пластинок или волокон, а как стройная разветвленная система. Изолированный в нерастворимой форме он еще сохраняет морфологическую структуру волокна [15]. Морфология лигнина выявляет неравномерную структуру без пространственной решетки; лигнин должен быть аморфным высокополимерным веществом. Его пермутоидные свойства объясняют его огромную реакционную способность и тот факт, что в нерастворен-ном состоянии он реагирует легко и совершенно однообразно [47]. Он никогда не встречается в природе один, а всегда в сочетании с целлюлозой или другими углеводами.[ …]
Целлюлоза в клеточных стенках волокон древесины и других растений тесно связана со своими спутниками — лигнином, гемицеллюлозами, экстрактивными веществами и пр. С ходом лигнификации разделение древесины на составные части все более затрудняется. Лигнин в древесной ткани находится в очень тесной связи с целлюлозой и может даже частично проникать внутрь микрофибрилл целлюлозы. Часть гемицеллюлоз очень тесно ассоциирована с целлюлозой (по-видимому, совместно ориентирована с ней). Эти тесно ассоциированные с целлюлозой фракции гемицеллюлоз часто называют целлюлозанами (см. стр. 91). Целлюлозаны практически не могут быть удалены из древесины без заметного повреждения самой целлюлозы. Поэтому в препаратах выделенной из древесинны целлюлозы всегда содержатся пентозаны (ксилан).[ …]
Часть гемицеллюлоз очень тесно ассоциирована с целлюлозой (по-видимому, совместно ориентирована с ней). Эти тесно ассоциированные с целлюлозой фракции гемицеллюлоз часто называют целлюлозанами (см. стр. 91). Целлюлозаны практически не могут быть удалены из древесины без заметного повреждения самой целлюлозы. Поэтому в препаратах выделенной из древесинны целлюлозы всегда содержатся пентозаны (ксилан).[ …]
Недавно было обнаружено, что в растениях встречается целый ряд подобных эфиров коричной и хинной кислот, моносахаридов и белковых веществ. Однако неизвестно, капой из этих эфиров играет наиболее важную роль в процессе лигнификации. Эль—Басиуни и сотр. /19/ разделили эти эфиры на растворимые и нерастворимые в этиловом спирте. По-видимому, нерастворимые эфиры имеют непосредственное отношение к образованию лигнина.[ …]
Еще в конце прошлого века Класон высказал гениальную догадк том, что в основе лигнина лежит конифериловый спирт.[ …]
О роли цитокннинов в регуляции активности камбия известно немного, но работа, проведенная на изолированных отрезках стеблей гороха, показала, что эти гормоны также могут стимулировать деления клеток в камбии и усиливать лигнификацию развивающихся клеток ксилемы. Обработка растений этиленом и абсцизовой кислотой влияет на активность камбия, по пока нет данных об участии этих веществ в естественном процессе регуляции деления камбиальных клеток и дифференцировки проводящей ткани.[ …]
Вязкость целлюлозы доводится до нужного уровня в процессе отбелки гипохлоритом. При этом решающее значение имеет характер предварительной обработки целлюлозы. Режим отбелки гипохлоритом для получения заданных показателей определяется степенью де-лигнификации поступающей целлюлозы и ее. вязкостью. Если предшествующие операции, начиная с варки и кончая облагораживанием, проведены правильно, доведение вязкости до соответствующего значения не представляет трудностей. В противном случае получение требуемой вязкости целлюлозы либо вообще невозможно, если целлюлоза сильно разрушена, либо сопровождается большой потерей волокна и снижением содержания а-целлюлозы.[ …]
Еще недавно любая схема отбелки начиналась со ступени хлорирования с последующим щелочением. В результате взаимодействия хлора с лигнином образуются хлорпроизводные лигнина, которые растворяются при последующем щелочении. Целью этих обработок является только де-лигнификация, так как в результате их проведения наблюдается лишь небольшое повышение белизны. Подобно хлорированию и щелочению обработки кислородом и озоном используются преимущественно для делигни-фикации в начале схемы отбелки.[ …]
Целью этих обработок является только де-лигнификация, так как в результате их проведения наблюдается лишь небольшое повышение белизны. Подобно хлорированию и щелочению обработки кислородом и озоном используются преимущественно для делигни-фикации в начале схемы отбелки.[ …]
Приемлемыми методами для определения целлюлозы считают такие методы, которые обеспечивают делигнификацию одревесневшей ткани при минимальном гидролитическом и окислительном воздействии на целлюлозу. Эти требования ограничивают выбор делигнифицирующих реагентов и условий проведения де-лигнификации.[ …]
В последнее время были затрачены немалые усилия для установления строения лигнина. Биосинтетическое воспроизведение лигнификации in vitro в лабораторных условиях составило определенный прогресс в химии лигнина, и полученный таким образом искусственный лигнин дал значительную информацию и для исследования структуры нативного лигнина.[ …]
В последнее время были затрачены немалые усилия для установления строения лигнина. Биосинтетическое воспроизведение лигнификации in vitro в лабораторных условиях составило определенный прогресс в химии лигнина, и полученный таким образом искусственный лигнин дал значительную информацию и для исследования структуры нативного лигнина.[ …]
Биосинтетическое воспроизведение лигнификации in vitro в лабораторных условиях составило определенный прогресс в химии лигнина, и полученный таким образом искусственный лигнин дал значительную информацию и для исследования структуры нативного лигнина.[ …]
Модельные опыты полимеризации смеси кониферилового и сина-пилового спиртов при действии этого же фермента привели к образованию аморфного сополимера, который имел сходство с лигнином покрытосеменных растений. Эти работы привели к заключению, что действие ферментной системы грибов на полимеризацию фенилпропаноидных соединений аналогично процессу лигнификации в высших растениях.[ …]
Молодые клетки каллуса могут дифференцироваться в клетки трахеид и элементы флоэмы. И в этих случаях большое значение имеет отношение ауксин/цитокинин и концентрация сахарозы. Ауксин в сочетании с сахарозой индуцирует формирование проводящих пучков, причем низкий уровень сахарозы благоприятствует образованию ксилемы, а высокий — флоэмы. Значение гормонального фактора (ауксина) для дифференцировки проводящих пучков иллюстрируется одним экспериментом Камю (Camus). Если в каллус вводятся почки, то ниже почек образуются тяжи проводящих пучков из клеток каллуса. Очевидно, что образование проводящих пучков индуцируется почкой—это можно доказать, поместив между почкой и каллусом пластинку целлофана: легко проницаемый целлофан не препятствует индукции (рис. 16.1).[ …]
Если в каллус вводятся почки, то ниже почек образуются тяжи проводящих пучков из клеток каллуса. Очевидно, что образование проводящих пучков индуцируется почкой—это можно доказать, поместив между почкой и каллусом пластинку целлофана: легко проницаемый целлофан не препятствует индукции (рис. 16.1).[ …]
В последующих разделах (ем. часть II) будет исчерпывающе рассмотрена химия клеточных стенок, относительные количества лигнина в них и другие родственные темы. Однако заканчивая рассмотрение четвертой и конечной фазы онтогенеза древесной клетки, следует упомянуть о некоторых явлениях, которые тем или иным путем связаны с лигннфикацией, как ее пошшают ботаники. Подобно образованию и разрастанию клеток, а также утолщению клеточных стенок, лигнификация может происходить лишь при жизни клеточного протопласта, так как отмершие клетки не могут лигнифи-цировать свои стенки. Процесс лигнификации может быть закончен в слое межклеточного вещества и в первичной стенке, но может продолжаться во вторичной стенке, даже если этот названный последним слой еще центростремительно увеличивается в толщину. В древесине деревьев лигнификация часто очень скоро заканчивается в слое, примыкающем к внутренней стороне камбия, обычно почти одновременно с тем, когда новые клетки достигли своего наибольшего размера, а вторичные стенки — своей конечной толщины. Это объясняет, почему заболонь при одинаковом содержании влаги так же или почти так же крепка, как ядровая древесина.[ …]
В древесине деревьев лигнификация часто очень скоро заканчивается в слое, примыкающем к внутренней стороне камбия, обычно почти одновременно с тем, когда новые клетки достигли своего наибольшего размера, а вторичные стенки — своей конечной толщины. Это объясняет, почему заболонь при одинаковом содержании влаги так же или почти так же крепка, как ядровая древесина.[ …]
Положительная роль регулируемой газовой среды в замедлении процессов послеуборочного дозревания объясняется также и влиянием газового состава на образование и биологическую активность растительного гормона этилена в плодах, который, как известно, даже в минимальных количествах (1:2000) заметно ускоряет процесс созревания, а затем и старения. Этилен воздействует и на синтез фенольных соединений. Повышая активность фенилаланин аммоний лиазы — ключевого фермента фенольного метаболизма, этилен способствует накоплению промежуточных продуктов фенольного обмена, которые могут вызвать появление неприятного привкуса или стать основой синтеза лигнина. Лигнификация ухудшает структуру-растительной ткани. С этим процессом связано, например, затвердение сердцевины батата. В присутствии этилена становится жесткой также спаржа, а у моркови вследствие накопления фенольных соединений появляется горький привкус. В среде с низким содержанием кислорода сильно подавляется биосинтез этилена: вначале он до определенного количества накапливается, а затем его концентрация остается неизменной. В данном случае сам этилен выполняет роль ингибитора ферментативных реакций его обра-• зования. Повышенные концентрации С02, являющегося, конкурентным ингибитором этилена, снижают биологическую активность последнего. Таким образом, в атмосфере с повышенным содержанием С02 и пониженной концентрацией 02 действие этилена как стимулятора созревания плодов практически устранено или значительно ослаблено.[ …]
Лигнификация ухудшает структуру-растительной ткани. С этим процессом связано, например, затвердение сердцевины батата. В присутствии этилена становится жесткой также спаржа, а у моркови вследствие накопления фенольных соединений появляется горький привкус. В среде с низким содержанием кислорода сильно подавляется биосинтез этилена: вначале он до определенного количества накапливается, а затем его концентрация остается неизменной. В данном случае сам этилен выполняет роль ингибитора ферментативных реакций его обра-• зования. Повышенные концентрации С02, являющегося, конкурентным ингибитором этилена, снижают биологическую активность последнего. Таким образом, в атмосфере с повышенным содержанием С02 и пониженной концентрацией 02 действие этилена как стимулятора созревания плодов практически устранено или значительно ослаблено.[ …]
Ранее нами было выявлено, что предобработка проростков пшеницы салициловой кислотой (СК) оказывает предадаптирующий эффект на растения к последующему воздействию засоления. В спектре защитного действия СК важное место занимает ее эффект на транзитное усиление продукции активных форм кислорода (АФК) и активацию супероксиддисмута-зы и пероксидазы в корнях. В условиях же засоления СК способствовала снижению резкой стресс-индуцированной продукции АФК и, соответственно, активности антиоксидантных ферментов (Сахабутдинова, Фархутдино-ва, Шакирова, 2004). Одним из механизмов защиты растений от действия стрессовых факторов является укрепление барьерных свойств клеток. Важную роль в лигнификации клеточных стенок играют фенилаланиниамми-ак-лиазы (ФАП), анионная пероксидаза и АФК. Показано, что предобработка проростков пшеницы СК способствовала ускорению образования лигнина в корнях при воздействии 2% NaCI по сравнению с СК-необрабо-танными растениями. Интересно, что сама обработка СК также ускоряла лигнификацию клеток пучков ксилемы, вероятно, за счет повышения активности ФАП и содержания анионной пероксидазы. Образование лигнина в клетках корней в предобработайных СК растениях является важным механизмом защиты к засолению, о чем свидетельствуют данные по снижению уровня пероксидации липидов и выхода электролитов из клеток обработанных СК проростков, что отражалось в существенном уменьшении степени ингибирующего действия 2% NaCI на рост растений пшеницы.
В спектре защитного действия СК важное место занимает ее эффект на транзитное усиление продукции активных форм кислорода (АФК) и активацию супероксиддисмута-зы и пероксидазы в корнях. В условиях же засоления СК способствовала снижению резкой стресс-индуцированной продукции АФК и, соответственно, активности антиоксидантных ферментов (Сахабутдинова, Фархутдино-ва, Шакирова, 2004). Одним из механизмов защиты растений от действия стрессовых факторов является укрепление барьерных свойств клеток. Важную роль в лигнификации клеточных стенок играют фенилаланиниамми-ак-лиазы (ФАП), анионная пероксидаза и АФК. Показано, что предобработка проростков пшеницы СК способствовала ускорению образования лигнина в корнях при воздействии 2% NaCI по сравнению с СК-необрабо-танными растениями. Интересно, что сама обработка СК также ускоряла лигнификацию клеток пучков ксилемы, вероятно, за счет повышения активности ФАП и содержания анионной пероксидазы. Образование лигнина в клетках корней в предобработайных СК растениях является важным механизмом защиты к засолению, о чем свидетельствуют данные по снижению уровня пероксидации липидов и выхода электролитов из клеток обработанных СК проростков, что отражалось в существенном уменьшении степени ингибирующего действия 2% NaCI на рост растений пшеницы. Полученные данные свидетельствуют о вовлечении ФАП и анионной пероксидазы, участвующей с привлечением АФК в образовании лигнина, в спектр защитного действия СК на растения пшеницы.[ …]
Полученные данные свидетельствуют о вовлечении ФАП и анионной пероксидазы, участвующей с привлечением АФК в образовании лигнина, в спектр защитного действия СК на растения пшеницы.[ …]
Что касается сезонной динамики названных показателей, кратко охарактеризуем ее в соответствии с данными, полученными нами для листьев березы повислой (1997-2001). Так, содержание фотосинтетических пигментов (хлорофиллов А, В, каротиноидов) в листьях березы достигало максимума в июне — июле, к августу снижалось примерно в два раза, к сентябрю, в зависимости от пункта наблюдений, находилось на уровне 1/4-1/6 от сезонного максимума. Наибольшее количество, как и максимально выраженные его изменения, было обнаружено для хлорофилла А, тогда как количества хлорофилла В и каротиноидов были близки и характеризовались сходной динамикой. Содержание свободных аминокислот в листьях березы, высокое в начале вегетационного периода — во время формирования листового аппарата, в июне несколько снижалось, далее наблюдался июльский подъем, очевидно, вызванный воздействием высокой температуры. После августовского снижения содержания аминокислот к сентябрю их количество вновь возрастало в результате связанного со старением листьев протеолиза. Водорастворимые фенольные соединения в листьях березы обнаруживались в наибольших количествах в начале вегетационного периода и его конце. Это соответствовало использованию низкомолекулярных фенолов для лигнификации формирующихся проводящих и механических тканей побега (май), накоплению фенолов в стареющих листьях, подготовке побега в целом к будущему периоду покоя.[ …]
После августовского снижения содержания аминокислот к сентябрю их количество вновь возрастало в результате связанного со старением листьев протеолиза. Водорастворимые фенольные соединения в листьях березы обнаруживались в наибольших количествах в начале вегетационного периода и его конце. Это соответствовало использованию низкомолекулярных фенолов для лигнификации формирующихся проводящих и механических тканей побега (май), накоплению фенолов в стареющих листьях, подготовке побега в целом к будущему периоду покоя.[ …]
Древесные клетки при повторном делении материнской клетки в боковом камбии расположены радиальными рядами (рис. 8). Клетки данного ряда вне зависимости от того, являются ли они лубяными или древесными, происходят от одной и той же камбиальной материнской клетки. Радиальное размещение элементов, возникающих таким образом, хорошо сохраняется в хвойных породах; в лиственных породах, наоборот, это размещение выражено нечетко или совсем незаметно, что объясняется послекамбиаль-ным развитием пор (сосуды в поперечном сечении), которые часто занимают в несколько раз большую площадь, чем клетки, от которых они происходят. На рис. 8 можно видеть с какой быстротой древесные клетки созревают позади камбия. Этот вопрос имеет большое значение для химиков. Камбий (несколько потерявший первоначальную форму из-за нежной природы ткани) расположен у ряда 2. Ксилярные клетки в процессе созревания показаны у ряда 3. Примерно на расстоянии 10—12 клеток позади камбия эти элементы, по-видимому, уже созрели, т. е. они уже прешли в живом состоянии через все фазы вышеописанного онтогенеза древесных клеток. Трахеиды, расположенные в самой наружной части прироста, являются теперь мертвыми клетками, хотя они составляют часть заболони. Древесные клетки позади камбия созревают в течение того года, в который они образуются. Если они по природе прозенхшиатичны, то теряют свой протопласт, как только завершается лигнификация; с другой стороны, древесная паренхима продолжает жить, пока находится в заболони, даже если она прошла через четыре фазы онтогенеза.[ …]
На рис. 8 можно видеть с какой быстротой древесные клетки созревают позади камбия. Этот вопрос имеет большое значение для химиков. Камбий (несколько потерявший первоначальную форму из-за нежной природы ткани) расположен у ряда 2. Ксилярные клетки в процессе созревания показаны у ряда 3. Примерно на расстоянии 10—12 клеток позади камбия эти элементы, по-видимому, уже созрели, т. е. они уже прешли в живом состоянии через все фазы вышеописанного онтогенеза древесных клеток. Трахеиды, расположенные в самой наружной части прироста, являются теперь мертвыми клетками, хотя они составляют часть заболони. Древесные клетки позади камбия созревают в течение того года, в который они образуются. Если они по природе прозенхшиатичны, то теряют свой протопласт, как только завершается лигнификация; с другой стороны, древесная паренхима продолжает жить, пока находится в заболони, даже если она прошла через четыре фазы онтогенеза.[ …]
Многочисленные анатомические исследования различных видов древесины в процессе ее развития показали, что молодые клетки вблизи камбия не содержат лигнина [1]. В дальнейшем, по мере утолщения клеточных стенок, относительное количество лигнина в них постепенно возрастает. Однако наибольшее количество лигнина откладывается в последней стадии развития клеток, перед их отмиранием. В этот период содержание лигнина в древесине достигает предельной величины, характерной для созревшей, мертвой ткани. Содержание полисахаридов, состоящих из пектиновых веществ, гемицеллюлоз и целлюлозы, в противоположность лигнину по мере старения клеток постепенно уменьшается (рис. 31). Необходимо, однако, учитывать, что на рис. 31 содержание отдельных компонентов в клеточных стенках трахеид приведено в относительных процентах. В действительности по мере увеличения толщины клеточных стенок в них откладываются слои неодинакового состава. Кроме того, отсутствовавший в межклетном, веществе и первичной оболочке лигнин к концу развития клетки откладывается там в наибольших количествах. Это наблюдение, сделанное с помощью цветных реакций на лигнин и углеводы, было подтверждено прямым определением содержания лигнина в срединной пластинке древесины дугласовой пихты, выделенной с помощью микроманипулятора [2].
В дальнейшем, по мере утолщения клеточных стенок, относительное количество лигнина в них постепенно возрастает. Однако наибольшее количество лигнина откладывается в последней стадии развития клеток, перед их отмиранием. В этот период содержание лигнина в древесине достигает предельной величины, характерной для созревшей, мертвой ткани. Содержание полисахаридов, состоящих из пектиновых веществ, гемицеллюлоз и целлюлозы, в противоположность лигнину по мере старения клеток постепенно уменьшается (рис. 31). Необходимо, однако, учитывать, что на рис. 31 содержание отдельных компонентов в клеточных стенках трахеид приведено в относительных процентах. В действительности по мере увеличения толщины клеточных стенок в них откладываются слои неодинакового состава. Кроме того, отсутствовавший в межклетном, веществе и первичной оболочке лигнин к концу развития клетки откладывается там в наибольших количествах. Это наблюдение, сделанное с помощью цветных реакций на лигнин и углеводы, было подтверждено прямым определением содержания лигнина в срединной пластинке древесины дугласовой пихты, выделенной с помощью микроманипулятора [2]. В последней было найдено около 71% лигнина при среднем содержании его в древесине 28%. Предсуществование части гемицеллюлоз в клетках молодой древесины до их лигнификации, а также возникновение из камбия лубяной ткани, содержащей пектиновые вещества, целлюлозу и гемицеллюлозы, которые в живой ткани не лигнифицируются, дает основание предполагать, что основная масса лигнина и гемицеллюлоз откладывается в клеточных стенках на разных стадиях их развития.[ …]
В последней было найдено около 71% лигнина при среднем содержании его в древесине 28%. Предсуществование части гемицеллюлоз в клетках молодой древесины до их лигнификации, а также возникновение из камбия лубяной ткани, содержащей пектиновые вещества, целлюлозу и гемицеллюлозы, которые в живой ткани не лигнифицируются, дает основание предполагать, что основная масса лигнина и гемицеллюлоз откладывается в клеточных стенках на разных стадиях их развития.[ …]
путь вперед в жизнь — BODY CENTERED LIVING
Мы часто думаем о мягкой части растения как о наполненной жизнью и прекрасной. Прорастающее семя, бутонирующий стебель, цветки гладкие и яркие.
Мы ценим молодежь и те части системы, которые потребляют энергию и привлекают все внимание.
Дети и молодые люди думают, что они занимают центральное место. И правильно! Разоблаченные, как они есть, незащищенные от извлеченных уроков. У руля самотворения.
О, но быть одревесневшим! Быть ожесточенным в самых своих клетках. Быть силой, которая поддается новому росту и более глубоким корням. Иметь способность быть старшей сестрой, родителем, великим и скромным дедушкой.
Быть силой, которая поддается новому росту и более глубоким корням. Иметь способность быть старшей сестрой, родителем, великим и скромным дедушкой.
Я наткнулся на термин одревесневший как на желательное, так и на необходимое качество для размножения инжира. При обрезке оставьте одеревеневшие ветки, воткните их — на два узла вглубь — в землю, и вуаля, у вас новая инжир!
«Лигнификация клеточной стенки — сложный процесс, происходящий исключительно у высших растений; его основная функция заключается в укреплении сосудистого тела растения».
Эта закалка является желаемой чертой, и я хочу рассказать вам о ней подробнее.
Люди… понимаете.
С людьми бывает трудно ориентироваться. Особенно, когда мы любим и заботимся о них и хотим, чтобы они были нам другом, братом, сестрой, матерью, ребенком. Когда мы хотим, чтобы они были рядом и доверчивы и веселились, разделяя с нами дни этой жизни.
Что ж, радость от того, что у тебя есть одревесневший друг, — это настоящая радость.
У меня есть воспоминание из моего 20-летнего возраста. На травянистой вершине холма с видом на великолепие национального парка Терра Булга. Мой друг и я сидели рядом друг с другом, глядя на луну в дневном небе. Мы точно были друзьями. Наши дети ходили в одну школу, и мы тусовались, когда могли, и до сих пор узнавали друг друга. В эти выходные мы покинули город, чтобы разбить лагерь на природе. Сама эта прогулка была «одревеснением» дружбы.
Но в этот момент, когда мы оба сидели, прижав колени к груди, я почувствовал потребность отпустить и положил голову ей на плечо. Просто протянул руку и отдал все свои владения, не зная, как она отреагирует.
Я мог бы годами жить с этим скрытым чувством «хождения по яичной скорлупе» вокруг нее. Такой, который остается незамеченным и возникает из-за того, что он не знает, на каком месте стоит тот или иной аспект жизни. Но когда моя голова коснулась ее плеча, я почувствовал, что она крепка, как скала. Смелее глубоко зарыто в землю. Неподвижный. Ее взгляд устремлен вдаль.
Неподвижный. Ее взгляд устремлен вдаль.
Вся моя неуверенность, сомнения в себе, перепроверка себя, все напрасно. Она была невозмутима мной. Неподвижный. Она была целостной и завершенной. Я был относительно незначительным. Как бабочка, приземлившаяся на гору. Ее деревянистые клетки представляли для меня ее устойчивое чувство собственного достоинства. Другими словами, она сидела в своем собственном центре.
Обнаружение этого качества в другом действительно освободило меня.
Итак, да, мы можем быть мягкими, чувствительными, нежными и ходить легко, но стабилизировать себя — это тоже божественно и возвышенно.
Фиговому дереву нужны подросшие, более сильные и твердые части, чтобы каждый год тянуться и плодоносить.
Наша эмоциональная жизнь тоже нуждается в твердых, ограниченных областях. Слишком озабоченный «нежным и милым» не всегда будет лучшим путем. Хорошее слишком часто заставляет нас пропускать или скользить по песчаным, значимым частям жизни.
Я не говорю ужесточаться. Не по-старому, послевоенному. Я говорю: стань сильным, когда будешь готов. Слегка надавите на края. Мягко идите к дискомфорту. Оставайтесь в проблемах личности друга, или гнилых коллег, или чего-то еще, чего вы быстро избегаете. Поиграйте немного в песке. Копните немного глубже, чтобы прикоснуться к песку и сделать его своим. Создайте жизнь, которая приведет вас в соответствие с вашим глубочайшим, истинным Я. Найдите свой более глубокий, спокойный центр.
Не по-старому, послевоенному. Я говорю: стань сильным, когда будешь готов. Слегка надавите на края. Мягко идите к дискомфорту. Оставайтесь в проблемах личности друга, или гнилых коллег, или чего-то еще, чего вы быстро избегаете. Поиграйте немного в песке. Копните немного глубже, чтобы прикоснуться к песку и сделать его своим. Создайте жизнь, которая приведет вас в соответствие с вашим глубочайшим, истинным Я. Найдите свой более глубокий, спокойный центр.
Многие приходят на йогу, чтобы стать более гибкими. Я пришел, чтобы найти радость в стабильности. И это требует выработки небольшого количества лигнина, чтобы поддерживать прочность клеточных стенок.
Еще одно определение, которое мне нравится:
«Лигнификация — это динамичный, гибкий процесс укрепления клеточных стенок в соответствии с различными потребностями (проводимость воды, механическая поддержка, защита) растения в течение его жизни».
Обратите внимание на то, как я использую слово «жесткий»: это слово нехорошее, но вся эта статья посвящена тому, чтобы немного больше заинтересоваться твердостью. Оно не должно автоматически означать плохое, мертвое, препятствующее, старое, безжизненное, подлое, невосприимчивое.
Оно не должно автоматически означать плохое, мертвое, препятствующее, старое, безжизненное, подлое, невосприимчивое.
Обратите внимание: в растениях этот процесс необратим. Как люди, мы могли бы проявлять некоторую осторожность: пока мы крепко держимся за то, что связывает и стабилизирует нас, мы также можем поддерживать текучесть тех мест, где могут процветать любовь, сострадание и радость.
Клеточная биология лигнификации высших растений
Искать:Статьи
- AJ Cann
Обзор современного понимания биосинтеза и полимеризации лигнина.
Клеточная биология лигнификации высших растений Лигнин (лат. lignum «древесина») представляет собой полифенольный полимер, откладывающийся непосредственно в клеточной стенке специализированных клеток. Он не только ограничен древесными тканями растений, но и представляет собой неотъемлемую функцию, обеспечивающую правильную клеточную функцию многих других типов клеток в различных тканях/органах растения. Появление лигнина в ходе эволюции растений совпало с появлением в девоне сосудистых наземных растений. Механически более слабый, чем целлюлоза, лигнин, тем не менее, значительно упрочняет любую клеточную стенку, обеспечивая дополнительную прочность на растяжение.
Он не только ограничен древесными тканями растений, но и представляет собой неотъемлемую функцию, обеспечивающую правильную клеточную функцию многих других типов клеток в различных тканях/органах растения. Появление лигнина в ходе эволюции растений совпало с появлением в девоне сосудистых наземных растений. Механически более слабый, чем целлюлоза, лигнин, тем не менее, значительно упрочняет любую клеточную стенку, обеспечивая дополнительную прочность на растяжение.
Несмотря на то, что лигнин является вторым наиболее распространенным наземным биополимером после целлюлозы, наше понимание образования лигнина остается фрагментарным. В отличие от целлюлозы, которая имеет определенную биохимическую структуру независимо от типа растительной клетки, образование лигнина является клеточно-специфическим и демонстрирует как четкую внутриклеточную локализацию, так и мономерный состав: таким образом, общий механизм лигнификации не может быть описан для всех типов лигнифицированных клеток и может объяснить, почему одревеснение до сих пор изучено лишь частично. Наше текущее биохимическое понимание состоит в том, что лигнин образуется в промежутках между микрофибриллами целлюлозы в результате окислительного связывания свободных мономеров лигнина, секретируемых непосредственно в клеточную стенку растения.
Наше текущее биохимическое понимание состоит в том, что лигнин образуется в промежутках между микрофибриллами целлюлозы в результате окислительного связывания свободных мономеров лигнина, секретируемых непосредственно в клеточную стенку растения.
В недавней статье в журнале Annals of Botany рассматривается современное понимание биосинтеза и полимеризации лигнина и отмечается, что в зависимости от типа клетки процесс лигнификации варьируется от полной автономии до полного сотрудничества с окружающими клетками-партнерами. Различные роли лигнина для функции каждого конкретного типа растительных клеток ясно иллюстрируются множественными фенотипическими дефектами, проявляемыми нокаутными мутантами в синтезе лигнина, что может объяснить, почему до сих пор не определен общий механизм лигнификации.
Баррос, Дж., Серк, Х., Гранлунд, И., и Песке, Э. (2015) Клеточная биология лигнификации высших растений. Анналы ботаники 115 (7): 1053–1074. doi: 10.1093/aob/mcv046
doi: 10.1093/aob/mcv046
Лигнин представляет собой полифенольный полимер, укрепляющий и защищающий от влаги клеточную стенку специализированных типов растительных клеток. Одревеснение является частью нормальной программы дифференцировки и функционирования определенных типов клеток, но также может быть вызвано в ответ на различные биотические и абиотические стрессы в клетках, которые иначе не стали бы одревесневать. Лигнификация клеточной стенки имеет специфические характеристики в зависимости от рассматриваемого типа клеток. Эти характеристики включают время лигнификации во время дифференцировки клеток, палитру связанных ферментов и субстратов, места субклеточного отложения, мономерный состав и клеточную автономию для производства мономера лигнина. В этом обзоре представлен обзор современного понимания биосинтеза и полимеризации лигнина на уровне клеточной биологии. Процесс лигнификации варьируется от полной автономии до полного сотрудничества в зависимости от типа клеток.