СИГНАЛ S-TERM-10 / КОВ-10 СКС Котел газовый
Котлы S-ТЕRМ и S-ТЕRМ В, предназначены для отопления домов, коттеджей и квартир, оборудованных системой водяного отопления c рабочим давлением воды до 0,13 МПа, а котел S-ТЕRМ В и для снабжения горячей водой для санитарных нужд.
Преимущества
- Пониженный уровень шума работы котла (применения аналога итальянской горелки Polidoro).
- КПД не менее 90%.
- Новая пилотная горелка (использование импортной запальной горелки).
- В качестве источника термоЭДС используется термопара.
- Использование современных комплектующих, доступных в любом регионе России.
- Легкий доступ к элементам газогорелочного устройства для обслуживания и ремонта.
- Современная технология окрашивания.
Россия, Казахстан, Белоруссия, Узбекистан, Армения, Киргизия, Таджикистан — доставка в любой город и другие страны ЕАЭС и мира.
Имя должно быть не менее :error символов.
Не правильный E-mail.
Название должно быть не менее :error символов.
Обязательное поле
Защита от спама reCAPTCHA Конфиденциальность и Условия использования
Сообщение отправлено
Пожалуйста, заполните форму правильно.
Отправка…
Капча недействительна.
Повторите попытку позже.
ПРОИЗВОДСТВО
Система организации производства ЭЗОТ СИГНАЛ основывается на принципах бережливого производства.
 Ежедневно устраняются потери в производстве. Гибкость и мобильность производственных процессов предприятия помогает своевременно удовлетворять любые потребности рынка.
Ежедневно устраняются потери в производстве. Гибкость и мобильность производственных процессов предприятия помогает своевременно удовлетворять любые потребности рынка.КАЧЕСТВО
Продукция предприятия является образцом качества. Высокий уровень качества продукции достигается с помощью безусловного и осознанного выполнения каждым сотрудником предприятия всех требований системы менеджмента качества.
ОРИЕНТАЦИЯ НА КЛИЕНТА
Приоритетом деятельности ЭЗОТ СИГНАЛ является полное удовлетворение требований и ожиданий клиентов посредством непрерывного совершенствования системы менеджмента, повышения качества и обновления номенклатуры выпускаемой продукции.
Газовые котлы серии Эконом
КОВ-6,5СТ, КОВ-10CT, АОГВ-11,6-М
Котлы водогрейные серии Комфорт
КОВ-СТ1пс и др.

Газовые котлы СТ 7-16 кВт
КОВ-7СТс, КОВ-12.5СТс и пр.
Газовые котлы СТ 20-40 кВт
КОВ-20СТс, КОВ-31.5СТс и пр.
Газовые котлы СТ 50-100 кВт
КОВ-50СТ, КОВ-63СТ, КОВ-80СТ
-
Газовые котлы СТПВ 10-16 кВт
КОВ-12.5СТПВ, КОВ-16СТПВ и пр.
Газовые котлы СТПВ 20-40 кВт
КОВ-20СТПВ, КОВ-40СТПВ и пр.
Газовые котлы СК(В)С 7-25 КВт
КОВ-16СКC и др.
Газовые котлы S-TERM
S-TERM 10, S-TERM 12.
 5, S-TERM 16В
5, S-TERM 16ВТермогенераторы
ТГ-2, ТГ-4 и пр.
Имя должно быть не менее :error символов.
Не правильный E-mail.
Название должно быть не менее :error символов.
Обязательное поле
Защита от спама reCAPTCHA Конфиденциальность и Условия использования
Сообщение отправлено
Пожалуйста, заполните форму правильно.
Отправка…
Капча недействительна.
Повторите попытку позже.
адрес для заявок: [email protected]- (7273)495-231
- (3955)60-70-56
- (8182)63-90-72
- (8512)99-46-04
- (3852)73-04-60
- (4722)40-23-64
- (4162)22-76-07
- (4832)59-03-52
- (423)249-28-31
- (4922)49-43-18
- (844)278-03-48
- (8172)26-41-59
- (473)204-51-73
- (343)384-55-89
- (4932)77-34-06
- (3412)26-03-58
- (395)279-98-46
- (843)206-01-48
- (4012)72-03-81
- (4842)92-23-67
- (3842)65-04-62
- (8332)68-02-04
- (4966)23-41-49
- (4942)77-07-48
- (861)203-40-90
- (391)204-63-61
- (4712)77-13-04
- (3522)50-90-47
- (4742)52-20-81
- (3519)55-03-13
- (495)268-04-70
- (8152)59-64-93
- (8552)20-53-41
- (831)429-08-12
- (3843)20-46-81
- (3496)41-32-12
- (383)227-86-73
- (3812)21-46-40
- (4862)44-53-42
- (3532)37-68-04
- (8412)22-31-16
- (8142)55-98-37
- (8112)59-10-37
- (342)205-81-47
- (863)308-18-15
- (4912)46-61-64
- (846)206-03-16
- (8342)22-96-24
- (812)309-46-40
- (845)249-38-78
- (8692)22-31-93
- (3652)67-13-56
- (4812)29-41-54
- (862)225-72-31
- (8652)20-65-13
- (3462)77-98-35
- (8212)25-95-17
- (4752)50-40-97
- (4822)63-31-35
- (8482)63-91-07
- (3822)98-41-53
- (4872)33-79-87
- (3452)66-21-18
- (8422)24-23-59
- (3012)59-97-51
- (347)229-48-12
- (4212)92-98-04
- (8352)28-53-07
- (351)202-03-61
- (8202)49-02-64
- (3022)38-34-83
- (4112)23-90-97
- (4852)69-52-93
Задайте вопрос прямо сейчас:
Извините, сервис временно недоступен.
Некорректный номер.
Ожидайте звонка на введенный номер.
front/header.call_free_error
Заказать обратный звонок
мы перезвоним Вам в рабочее время
Настоящее соглашение является официальным документом OOO «Новые Технологии», ОГРН 1131690023178, ИНН 1656069657 (далее – Администратор) и определяют порядок использования посетителями (далее — Посетитель) сайта Администратора и обработки информации, получаемой Администратором от Посетителя.
- Соглашение может быть изменено Администратором в одностороннем порядке в любой момент, без какого-либо специального уведомления Посетителя Сайта.
- В случае, если при использовании Посетителями Сайта Администратору будет сообщена какая-либо информация, относящаяся прямо или косвенно к определенному или определяемому физическому лицу (далее – Персональные данные), ее последующая обработка будет осуществляться в соответствии с законодательством Российской Федерации.
 В отношении всех сообщаемых Персональных данных Посетитель дает Администратору согласие на их обработку. Администратор обрабатывает персональные данные Посетителя исключительно в целях предоставления Посетителю функций Сайта, размещенного на нем контента, маркетинговой, рекламной, иной информации, в целях получения Посетителем персонализированной (таргетированной) рекламы, исследования и анализа данных Посетителя, а также в целях предложения Посетителю своих товаров и услуг. В отношении всех сообщенных Администратору Посетителем своих персональных данных Администратор вправе осуществлять сбор, систематизацию, накопление, хранение, уточнение (обновление, изменение), использование, распространение (в том числе передача любым третьим лицам, включая передачу персональных данных третьим лицам на хранение или в случае поручения обработки персональных данных третьим лицам), обезличивание, блокирование, уничтожение, трансграничную передачу, обработку с применением основных способов такой обработки (хранение, запись на электронных носителях и их хранение, составление перечней, маркировка) и иные действия в соответствии со статьей 3 Федерального закона от 27.
В отношении всех сообщаемых Персональных данных Посетитель дает Администратору согласие на их обработку. Администратор обрабатывает персональные данные Посетителя исключительно в целях предоставления Посетителю функций Сайта, размещенного на нем контента, маркетинговой, рекламной, иной информации, в целях получения Посетителем персонализированной (таргетированной) рекламы, исследования и анализа данных Посетителя, а также в целях предложения Посетителю своих товаров и услуг. В отношении всех сообщенных Администратору Посетителем своих персональных данных Администратор вправе осуществлять сбор, систематизацию, накопление, хранение, уточнение (обновление, изменение), использование, распространение (в том числе передача любым третьим лицам, включая передачу персональных данных третьим лицам на хранение или в случае поручения обработки персональных данных третьим лицам), обезличивание, блокирование, уничтожение, трансграничную передачу, обработку с применением основных способов такой обработки (хранение, запись на электронных носителях и их хранение, составление перечней, маркировка) и иные действия в соответствии со статьей 3 Федерального закона от 27. 07.2006 № 152-ФЗ «О персональных данных».
07.2006 № 152-ФЗ «О персональных данных». - Посетитель понимает и соглашается с тем, что предоставление Администратору какой-либо информации о себе, не являющейся контактной и не относящейся к целям, обозначенным Администратором Сайта (не относящейся к деятельности Администратора, к продвигаемым им товарам и/или услугам, к условиям сотрудничества Администратора и Посетителя Сайта), а равно предоставление информации, относящейся к государственной, банковской и/или коммерческой тайне, информации о расовой и/или национальной принадлежности, политических взглядах, религиозных или философских убеждениях, состоянии здоровья, интимной жизни Посетителя Сайта или иного третьего лица запрещено.
- В случае принятия Посетителем решения о предоставлении Администратору какой-либо информации (каких-либо данных), Посетитель обязуется предоставлять исключительно достоверную и актуальную информацию. Посетитель Сайта не вправе вводить Администратора в заблуждение в отношении своей личности, сообщать ложную или недостоверную информацию о себе.

- Администратор принимает меры для защиты Персональных данных Посетителя Сайта в соответствии с законодательством Российской Федерации.
- Администратор не проверяет достоверность персональной информации, предоставляемой Посетителем Сайта, и не имеет возможности оценивать его дееспособность. Однако Администратор исходит из того, что Посетитель предоставляет достоверную персональную информацию и поддерживает эту информацию в актуальном состоянии.
- Администратор вправе запрещать Посетителю доступ к Сайту или к отдельным частям Сайта.
- Посетитель в соответствии с ч. 1 ст. 18 Федерального закона «О рекламе» дает Администратору свое согласие на получение сообщений рекламного характера.
- Согласие может быть отозвано субъектом персональных данных или его представителем путем направления письменного заявления ООО «Новые Технологии» или его представителю по адресу: 420030 Казань, Адмиралтейская д.
 3 к.4 п.1026.
3 к.4 п.1026. - В случае отзыва субъектом персональных данных или его представителем Согласия на обработку персональных данных, ООО «Новые Технологии» вправе продолжить обработку без разрешения субъекта персональных данных при наличии оснований, указанных в пунктах 2 — 11 части 1 статьи 6, части 2 статьи 10 и части 2 статьи 11 Федерального закона №152-ФЗ «О персональных данных» от 26.06.2006 г.
- Настоящее Согласие действует все время до момента прекращения обработки персональных данных по причинам, указанным в п. 9 данного документа.
Клапан электромагнитный для котла КОВ «Сигнал»
Клапан электромагнитный для котла КОВ «Сигнал»
Для котлов отопительных КОВ-7, КОВ-10, КОВ-12,5, КОВ-16, КОВ-20, КОВ-25, КОВ-31,5, КОВ-40, КОВ-50, КОВ-63, КОВ-80, КОВ-100
Моделей СТ, СТс, СТПВ, СТн, СТПВ1пс, СТ1пс, СТПВс
При заказе просим уточнять год выпуска котла и его мощность.
Например ― КОВ-31.5 СТ 2001 г.
Самые распространенные клапаны в наличии всегда !
Возможна срочная отправка клапана в день оплаты ТК «СДЭК» (при договоренности)
Внешний вид клапанов для котлов разных лет выпуска отличается
Просьба !!! при заказе у нас запчастей уточняйте год выпуска и производителя на бирке прибора.
Это делается для более точного подбора необходимых запчастей именно к Вашему прибору. Например один и тот же прибор с названием РДБК1-50 выпускается уже более 60 лет. Первоначально его выпускали 2 завода, в 2000-х годах производителей было уже 4-5, а в последние годы число производителей стало свыше 10. Плюс некоторые заводы раз в несколько лет вносили изменения в конструкцию. Для пользователей данного оборудования это могло остаться незаметным, но оно отразилось на запчастях прибора. Могли поменяться размер и материал мембран, изменились штоки, пружины, материалы седел, пилотов. Поменялось как правило и само литье прибора – ранее оно было чугунным, а в последние годы его вытеснил алюминиевый сплав. Запчасти из одного металла заменяли на другой более дешевый или более распространенный. Плюс часть запчастей особенно в последние годы изменялась в сторону удешевления для получения ценового конкурентного преимущества. Или например рабочие мембраны раньше вырезались из специального мембранного полотна, а позднее могли замениться на литые из специальной резины с армирующей нитью. Данные изменения относятся ко всем известным типам газового оборудования, таким как регуляторы РДГ, РДБК, РДУК, РДСК, РДГД, клапанам КПЗ, ПСК, ПКН, ПКВ, ПКК, КПЭГ. Также сообщаем Вам что большинство вышеуказанных приборов за последние 65 лет производились именно в Саратовской области т.к. именно отсюда протянулся первый газопровод в России в 1945г. и одновременно здесь заработал первый завод газового оборудования и позднее образовался головной научно-исследовательский институт газа ГипроНИИгаз.
Поменялось как правило и само литье прибора – ранее оно было чугунным, а в последние годы его вытеснил алюминиевый сплав. Запчасти из одного металла заменяли на другой более дешевый или более распространенный. Плюс часть запчастей особенно в последние годы изменялась в сторону удешевления для получения ценового конкурентного преимущества. Или например рабочие мембраны раньше вырезались из специального мембранного полотна, а позднее могли замениться на литые из специальной резины с армирующей нитью. Данные изменения относятся ко всем известным типам газового оборудования, таким как регуляторы РДГ, РДБК, РДУК, РДСК, РДГД, клапанам КПЗ, ПСК, ПКН, ПКВ, ПКК, КПЭГ. Также сообщаем Вам что большинство вышеуказанных приборов за последние 65 лет производились именно в Саратовской области т.к. именно отсюда протянулся первый газопровод в России в 1945г. и одновременно здесь заработал первый завод газового оборудования и позднее образовался головной научно-исследовательский институт газа ГипроНИИгаз. Поэтому запчасти к вышеуказанным приборам наиболее вероятно Вы найдете именно в Саратове или городе-спутнике Энгельсе. Просьба присылать нам на нашу электронную почту фото бирки прибора. Там как правило указан производитель, год выпуска и марка прибора. Причем указанный производитель на бирке не всегда является реальным заводом изготовившим этот прибор. Прибор мог быть просто приобретен у другого производителя и на него впоследствии было установлена бирка от другого производителя имеющего тоже разрешительные документы на его выпуск или не имеющий таковых (совсем редкий случай). В случае если бирка прибора не читаема, то на ней возможно разглядеть логотип производителя. Если бирка на приборе отсутствует, то желательно прислать нам скан паспорта прибора. Там тоже указан производитель и год выпуска. В некоторых случаях паспорт тоже бывает от другого производителя т.к. старый паспорт был утерян и на замену был приложен аналогичный. В таком случае для определения принадлежности прибора нам понадобится фото его с разных сторон.
Поэтому запчасти к вышеуказанным приборам наиболее вероятно Вы найдете именно в Саратове или городе-спутнике Энгельсе. Просьба присылать нам на нашу электронную почту фото бирки прибора. Там как правило указан производитель, год выпуска и марка прибора. Причем указанный производитель на бирке не всегда является реальным заводом изготовившим этот прибор. Прибор мог быть просто приобретен у другого производителя и на него впоследствии было установлена бирка от другого производителя имеющего тоже разрешительные документы на его выпуск или не имеющий таковых (совсем редкий случай). В случае если бирка прибора не читаема, то на ней возможно разглядеть логотип производителя. Если бирка на приборе отсутствует, то желательно прислать нам скан паспорта прибора. Там тоже указан производитель и год выпуска. В некоторых случаях паспорт тоже бывает от другого производителя т.к. старый паспорт был утерян и на замену был приложен аналогичный. В таком случае для определения принадлежности прибора нам понадобится фото его с разных сторон. За счет многолетнего опыта нашей работы даже если прибор без бирки и с чужим паспортом мы в 90% случаев сможем определить чей он. Разобраться в этих многолетних изменениях конструкций и соответствий стороннему человеку крайне сложно. Для этого как минимум необходимы специалисты газовой отрасли с опытом работы с данным оборудованием разных производителей от 10-15 лет. На нашем предприятии в настоящее время есть сотрудники с опытом от 16 лет. Резюмируя все вышесказанное, для более быстрой обработки Вашей заявки ждем от Вас :
За счет многолетнего опыта нашей работы даже если прибор без бирки и с чужим паспортом мы в 90% случаев сможем определить чей он. Разобраться в этих многолетних изменениях конструкций и соответствий стороннему человеку крайне сложно. Для этого как минимум необходимы специалисты газовой отрасли с опытом работы с данным оборудованием разных производителей от 10-15 лет. На нашем предприятии в настоящее время есть сотрудники с опытом от 16 лет. Резюмируя все вышесказанное, для более быстрой обработки Вашей заявки ждем от Вас :
Год выпуска, производитель прибора, точная марка. Если эта информация неизвестна то ждем фото прибора с разных сторон и скан паспорта (первая и последние 2 страницы).
Клапаны электромагнитные для котлов КОВ с мелкой и крупной резьбой.
Список запасных частей для котла КОВ
Клапан электромагнитный для котла КОВ «Сигнал» c итальянской автоматикой MiniSit, EuroSit, NovaSit
Список основных запасных частей для котлов отопительных КОВ «Сигнал»:
— датчик безопасности по тяге ( датчик тяги )
— датчик безопасности по предельной температуре ( датчик температуры )
— термометр
— горелка запальная
— теплообменник с топкой
— провод свечи
— свеча пьезорозжига
— горелка ГГУ
— провод с контактами
Отправка клапанов для котла КОВ транспортными компаниями во все города России
Отправка клапана электромагнитного для котла КОВ компанией СДЭК
Доставка клапана электромагнитного для котлов отопительных КОВ-10, КОВ-12,5, КОВ-16, КОВ-20, КОВ-25, КОВ-31,5, КОВ-40, КОВ-50, КОВ-63, КОВ-80, КОВ-100 осуществляется транспортными компаниями по таким городам как :
Абакан, Анадырь, Архангельск, Астрахань, Барнаул, Белгород, Биробиджан, Благовещенск, Брянск, Владивосток, Владикавказ, Владимир, Волгоград, Вологда, Воронеж, Горно-Алтайск, Грозный, Дудинка, Екатеринбург, Иваново, Ижевск, Йошкар-Ола, Иркутск, Казань, Калининград, Калуга, Кемерово, Киров, Кострома, Краснодар, Красноярск, Кудымкар, Курган, Курск, Кызыл, Липецк, Магадан, Майкоп, Махачкала, Москва, Мурманск, Назрань, Нальчик, Нарьян-Мар, Нижний Новгород, Новгород, Новосибирск, Омск, Орел, Оренбург, Агинское, Палана, Тура, Усть-Ордынский, Пенза, Пермь, Петрозаводск, Петропавловск-Камчатский, Псков, Ростов-на-Дону, Рязань, Салехард, Самара, Санкт-Петербург, Саранск, Саратов, Севастополь, Симферополь, Смоленск, Ставрополь, Сыктывкар, Тамбов, Томск, Тверь, Тула, Тюмень, Улан-Удэ, Ульяновск, Уфа, Хабаровск, Ханты-Мансийск, Чебоксары, Челябинск, Черкесск, Чита, Элиста, Южно-Сахалинск, Якутск, Ялта, Ярославль, Астана, Абай, Акколь, Аксай, Аксу, Актау, Актобе, Алга, Алматы, Арал, Аркалык, Арыс, Атбасар, Атырау, Аягоз, Байконыр, Балхаш, Булаево, Державинск, Ерейментау, Есик, Есиль, Жанаозен, Жанатас, Жаркент, Жезказган, Жем, Жетысай, Житикара, Зайсан, Зыряновск, Казалинск, Кандыагаш, Капшагай, Караганды, Каражал, Каратау, Каркаралинск, Каскелен, Кентау, Кокшетау, Костанай, Кулсары, Курчатов, Кызылорда, Ленгер, Лисаковск, Макинск, Мамлютка, Павлодар, Петропавловск, Приозёрск, Риддер, Рудный, Сарань, Сарканд, Сарыагаш, Сатлаев, Семей, Сергеевка, Серебрянск, Степногорск, Степняк, Тайынша, Талгар, Талдыкорган, Тараз, Текели, Темир, Темиртау, Туркестан, Уральск, Усть-Каменогорск, Ушарал, Уштобе, Форт-Шевченко, Хромтау, Шардара, Шалкар, Шар, Шахтинск, Шемонаиха, Шу, Шымкент, Щучинск, Экибастуз, Эмба, Керчь, Евпатория, Феодосия, Джанкой, Алушта, Бахчисарай, Саки, Красноперекопск, Армянск, Судак, Белогорск, Инкерман, Щёлкино, Старый Крым, Алупка, Форос, Гаспра, Гурзуф, Массандра, Ливадия, Симеиз, Кореиз, Морское, Курортное, Новый Свет, Коктебель, Орджоникидзе, Киев, Минск, Рига, Вильнюс, Кишинев, Таллин, Баку, Ереван, Тбилиси, Бишкек, Душанбе, Ашхабад, Ташкент, Крым .
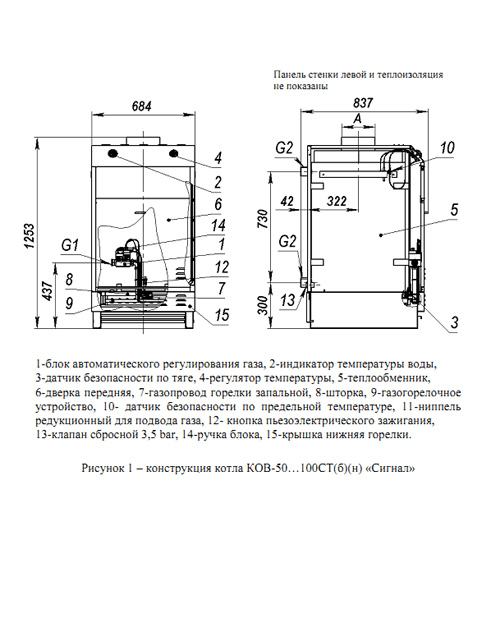
Сигнал-Теплотехника КОВ-10 СКс цена, характеристики, видео обзор, отзывы
- Тип отопительного котла газовый
- Число контуров одноконтурный
- Максимальная мощность обогрева 10 кВт
- Отапливаемая площадь 120 кв. м.
- КПД 90 %
- Способ установки напольный
- Тип горелки газовая
- Камера сгорания открытая
- Энергонезависимый Есть
- Автоподжиг Нет
- Показать все
Видео обзоры Сигнал-Теплотехника КОВ-10 СКс
Характеристики Сигнал-Теплотехника КОВ-10 СКс
Общие характеристики *
| Тип отопительного котла | газовый |
| Число контуров | одноконтурный |
| Максимальная мощность обогрева | 10 кВт |
| Отапливаемая площадь | 120 кв. м. м. |
| КПД | 90 % |
| Способ установки | напольный |
| Тип горелки | газовая |
| Камера сгорания | открытая |
| Энергонезависимый | Есть |
| Автоподжиг | Нет |
| Бункер в комплекте | Нет |
| Подключение теплого пола | Нет |
| Вытяжной вентилятор | Нет |
Технические характеристики *
| Циркуляционный насос | Нет |
| Тип газового котла | конвекционный |
| Материал первичного теплообменника | сталь |
| Максимальная температура горячей воды | 90 °С |
| Максимальное давление в контуре отопления | 1. 3 бар |
| Минимальная температура горячей воды | 30 °С |
| Расширительный бак | Нет |
| Номинальное давление газа на входе | 13 мБар |
Характеристики подключения *
| Диаметр дымохода | 80 мм |
| Патрубок подключения контура отопления | 1» |
| Патрубок подключения газа | 1/2» |
Используемое топливо *
| Природный газ | Есть |
| Расход природного газа | 1. 2 куб. м/час |
Управление и диагностика *
| Управление | механическое |
| Индикация включения | Нет |
| Разъем для подключения внешнего управления | Нет |
| Термометр | Есть |
| Манометр | Нет |
Системы защиты *
| Защита от перегрева | Есть |
| Газ-контроль | Есть |
Габариты и вес *
| Высота | 655 мм |
| Вес | 38 кг |
| Ширина | 260 мм |
| Глубина | 470 мм |
* Точные характеристики уточняйте у продавца.
Другие модели
Сигнал-Теплотехника КОВ 16 СТПВпс 16 кВт двухконтурный
Сигнал-Теплотехника КОВ 12,5 СТПВ1пс 12.5 кВт двухконтурный
Сигнал-Теплотехника КОВ 20 СТпс 20 кВт одноконтурный
Сигнал-Теплотехника КОВ 20 СТПВпс 20 кВт двухконтурный
Сигнал-Теплотехника S-TERM 25 (КОВ-25 СКс) 25 кВт одноконтурный
30757 ₽
Сигнал-Теплотехника КОВ 20 СКВс 20 кВт двухконтурный
24858 ₽
Сигнал-Теплотехника КОВ 25 СКВс 25 кВт двухконтурный
33775 ₽
Сигнал-Теплотехника S-TERM 7 (КОВ-7 СКс) 7 кВт одноконтурный
19990 ₽
Сигнал 10 , 10 кВт, одноконтурный
24900 ₽
сигнал
26500 ₽
Домены и функции шиповидного белка при SARS-Cov-2 в контексте дизайна вакцины
1. Чжоу П., Ян С.-Л., Ван С.-Г., Ху Б., Чжан Л., Чжан W., Si H.-R., Zhu Y., Li B., Huang C.-L., et al. Вспышка пневмонии, связанная с новым коронавирусом вероятного происхождения от летучих мышей. Природа. 2020; 579: 270–273. doi: 10.1038/s41586-020-2012-7. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Чжоу П., Ян С.-Л., Ван С.-Г., Ху Б., Чжан Л., Чжан W., Si H.-R., Zhu Y., Li B., Huang C.-L., et al. Вспышка пневмонии, связанная с новым коронавирусом вероятного происхождения от летучих мышей. Природа. 2020; 579: 270–273. doi: 10.1038/s41586-020-2012-7. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2. Хоффманн М., Кляйне-Вебер Х., Пельманн С. Сайт многоосновного расщепления в шиповидном белке SARS-CoV-2 необходим для инфекции клеток легких человека. Мол. Клетка. 2020;78:779–784.e775. doi: 10.1016/j.molcel.2020.04.022. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
3. Хоффманн М., Кляйне-Вебер Х., Шредер С., Крюгер Н., Херрлер Т., Эриксен С., Ширгенс Т.С., Херрлер Г. ., Wu N.H., Nitsche A., et al. Проникновение в клетку SARS-CoV-2 зависит от ACE2 и TMPRSS2 и блокируется клинически проверенным ингибитором протеазы. Клетка. 2020;181:271–280.e278. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Ou X., Liu Y., Lei X., Li P., Mi D. , Ren L., Guo L., Guo R., Chen T ., Ху Дж. и др. Характеристика спайкового гликопротеина SARS-CoV-2 при проникновении вируса и его перекрестная иммунная реактивность с SARS-CoV. Нац. коммун. 2020;11:1620. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Ren L., Guo L., Guo R., Chen T ., Ху Дж. и др. Характеристика спайкового гликопротеина SARS-CoV-2 при проникновении вируса и его перекрестная иммунная реактивность с SARS-CoV. Нац. коммун. 2020;11:1620. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Ли Ф., Ли В., Фарзан М., Харрисон С.С. Структура домена, связывающего рецептор спайка коронавируса SARS, в комплексе с рецептором. Наука. 2005; 309: 1864–1868. doi: 10.1126/science.1116480. [PubMed] [CrossRef] [Google Scholar]
6. Lu G., Wang Q., Gao G.F. От летучей мыши к человеку: признаки шипа определяют «переход хозяина» коронавирусов SARS-CoV, MERS-CoV и других. Тенденции микробиол. 2015; 23: 468–478. doi: 10.1016/j.tim.2015.06.003. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Халсвит Р.Дж.Г., де Хаан К.А.М., Бош Б.Дж. Коронавирусный спайковый белок и изменения тропизма. В: Ziebuhr J., редактор. Успехи в исследованиях вирусов. Том 96. Академическая пресса; Кембридж, Массачусетс, США: 2016.![]() стр. 29–57. [Бесплатная статья PMC] [PubMed] [Google Scholar]
стр. 29–57. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Coutard B., Valle C., de Lamballerie X., Canard B., Seidah NG, Decroly E. Спайковый гликопротеин нового коронавируса 2019-nCoV содержит фуриноподобный сайт расщепления, отсутствующий в CoV той же клады. Антивир. Рез. 2020;176:104742. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Walls A.C., Tortorici M.A., Bosch B.-J., Frenz B., Rottier PJM, DiMaio F., Rey F.A., Veesler D. Криоэлектронная микроскопия структуры тримера гликопротеина шипа коронавируса. Природа. 2016; 531:114–117. doi: 10.1038/nature16988. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Предварительно слитая структура шиповидного белка человеческого коронавируса. Природа. 2016; 531:118–121. doi: 10.1038/nature17200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Уоллс А.С., Парк Ю.Дж., Торторичи М.А., Уолл А., Макгуайр А.Т., Вислер Д. Структура, функция и антигенность гликопротеина шипа SARS-CoV-2. Клетка. 2020;181:281–292.e286. doi: 10.1016/j.cell.2020.02.058. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Клетка. 2020;181:281–292.e286. doi: 10.1016/j.cell.2020.02.058. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Wrapp D., Wang N., Corbett K.S., Goldsmith J.A., Hsieh C.-L., Abiona O., Graham B.S., McLellan J.S. Крио-ЭМ структура шипа 2019-nCoV в конформации префузии. Наука. 2020;367:1260. doi: 10.1126/science.abb2507. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Lan J., Ge J., Yu J., Shan S., Zhou H., Fan S., Zhang Q., Shi X., Wang Q., Zhang L., et al. Структура домена, связывающего шиповидный рецептор SARS-CoV-2, связанного с рецептором ACE2. Природа. 2020; 581: 215–220. doi: 10.1038/s41586-020-2180-5. [PubMed] [CrossRef] [Google Scholar]
14. Шан Дж., Е Г., Ши К., Ван Ю., Луо С., Айхара Х., Гэн К., Ауэрбах А., Ли Ф. Structural основе распознавания рецепторов SARS-CoV-2. Природа. 2020; 581: 221–224. doi: 10.1038/s41586-020-2179-y. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
15. Tian X., Li C., Huang A. , Xia S., Lu S., Shi Z., Lu L., Jiang S., Yang Z., Wu Y., et al. Мощное связывание шиповидного белка нового коронавируса 2019 года с человеческим моноклональным антителом, специфичным к коронавирусу SARS. Эмердж. микроб. Заразить. 2020; 9: 382–385. doi: 10.1080/22221751.2020.1729069. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Xia S., Lu S., Shi Z., Lu L., Jiang S., Yang Z., Wu Y., et al. Мощное связывание шиповидного белка нового коронавируса 2019 года с человеческим моноклональным антителом, специфичным к коронавирусу SARS. Эмердж. микроб. Заразить. 2020; 9: 382–385. doi: 10.1080/22221751.2020.1729069. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
16. Xia X. DAMBE5: комплексный программный пакет для анализа данных в области молекулярной биологии и эволюции. Мол. биол. Эвол. 2013; 30:1720–1728. дои: 10.1093/молбев/mst064. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
17. Лай М.М., Кавана Д. Молекулярная биология коронавирусов. Доп. Вирус рез. 1997; 48:1–100. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Чакраборти С., Прабакаран П., Сяо Х., Димитров Д.С. Домен, связывающий рецептор гликопротеина S коронавируса SARS: точное картирование и функциональная характеристика. Вирол. Дж. 2005; 2:73. doi: 10.1186/1743-422X-2-73. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
19. Xiao X., Feng Y., Chakraborti S., Dimitrov D.S. Олигомеризация гликопротеина S SARS-CoV: димеризация N-конца и тримеризация эктодомена. Биохим. Биофиз. Рез. коммун. 2004; 322:93–99. doi: 10.1016/j.bbrc.2004.07.084. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Xiao X., Feng Y., Chakraborti S., Dimitrov D.S. Олигомеризация гликопротеина S SARS-CoV: димеризация N-конца и тримеризация эктодомена. Биохим. Биофиз. Рез. коммун. 2004; 322:93–99. doi: 10.1016/j.bbrc.2004.07.084. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
20. Beniac D.R., deVarennes S.L., Andonov A., He R., Booth T.F. Конформационная реорганизация спайка коронавируса SARS после связывания с рецептором: последствия для слияния мембран. ПЛОС ОДИН. 2007;2:e1082. doi: 10.1371/journal.pone.0001082. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Маду И.Г., Белузар С., Уиттакер Г.Р. Домен S2 шипа SARS-коронавируса, фланкированный остатками цистеина C822 и C833, важен для активации слияния мембран. Вирусология. 2009; 393: 265–271. doi: 10.1016/j.virol.2009.07.038. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22. Мицуки Ю.Ю., Охниси К., Такаги Х., Осима М., Ямамото Т., Мизукоши Ф., Терахара К., Кобаяси К., Ямамото Н. , Ямаока С. и др. Единственная аминокислотная замена в доменах шиповидных белков S1 и S2 определяет фенотип SARS-CoV, избегающий нейтрализации. микробы заражают. 2008;10:908–915. doi: 10.1016/j.micinf.2008.05.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Ямаока С. и др. Единственная аминокислотная замена в доменах шиповидных белков S1 и S2 определяет фенотип SARS-CoV, избегающий нейтрализации. микробы заражают. 2008;10:908–915. doi: 10.1016/j.micinf.2008.05.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Ng OW, Keng CT, Leung C.S., Peiris JS, Poon L.L., Tan YJ. нейтрализующее моноклональное антитело, нацеленное на домен S2. ПЛОС ОДИН. 2014;9:e102415. doi: 10.1371/journal.pone.0102415. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Song H.C., Seo M.Y., Stadler K., Yoo B.J., Choo Q.L., Coates S.R., Uematsu Y., Harada T., Greer C.E., Polo Дж. М. и др. Синтез и характеристика нативной олигомерной формы рекомбинантного гликопротеина спайка коронавируса тяжелого острого респираторного синдрома. Дж. Вирол. 2004;78:10328–10335. дои: 10.1128/ОВИ.78.19.10328-10335.2004. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
25. Jin D.Y., Zheng B.J. Роль шиповидного белка в патогенезе коронавируса SARS. Гонконг Мед. Дж. 2009; 15:37–40. [PubMed] [Google Scholar]
Гонконг Мед. Дж. 2009; 15:37–40. [PubMed] [Google Scholar]
26. Xia X. Контроль трансляции HAC1 путем регуляции сплайсинга в Saccharomyces cerevisiae . Междунар. Дж. Мол. науч. 2019;20:2860. doi: 10.3390/ijms20122860. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
27. Chow K.Y., Yeung Y.S., Hon C.C., Zeng F., Law K.M., Leung F.C. Опосредованной аденовирусом экспрессии С-концевого домена шиповидного белка SARS-CoV достаточно, чтобы вызвать апоптоз в клетках Vero E6. ФЭБС лат. 2005;579: 6699–6704. doi: 10.1016/j.febslet.2005.10.065. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Шаджахан А., Супекар Н.Т., Глейнич А.С., Азади П. Вывод профиля N- и О-гликозилирования шиповидного белка нового коронавируса SARS- КоВ-2. Гликобиология. 2020; 30: 981–988. doi: 10.1093/гликоб/cwaa042. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Андерсен К.Г., Рамбо А., Липкин В.И., Холмс Э.К., Гарри Р.Ф. Проксимальное происхождение SARS-CoV-2. Нац. Мед. 2020; 26: 450–452. дои: 10.1038/s41591-020-0820-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Нац. Мед. 2020; 26: 450–452. дои: 10.1038/s41591-020-0820-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Fagerberg L., Hallström B.M., Oksvold P., Kampf C., Djureinovic D., Odeberg J., Habuka M., Tahmasebpoor S., Даниэльссон А., Эдлунд К. и др. Анализ тканеспецифичной экспрессии человека путем полногеномной интеграции транскриптомики и протеомики на основе антител. Мол. Клетка. протеом. 2014;13:397–406. doi: 10.1074/mcp.M113.035600. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Sun X., Tse L.V., Ferguson A.D., Whittaker G.R. Модификации сайта расщепления гемагглютинина контролируют вирулентность нейротропного вируса гриппа h2N1. Дж. Вирол. 2010;84:8683. дои: 10.1128/ОВИ.00797-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Кидо Х., Окумура Ю., Такахаши Э., Пан Х.-Ю., Ван С., Яо Д., Яо М., Чида Дж., Яно М. Роль клеточных протеаз хозяина в патогенезе гриппа и полиорганной недостаточности, вызванной гриппом. Биохим. Биофиз. Acta Proteins Proteom. 2012; 1824: 186–194. doi: 10.1016/j.bbapap.2011.07.001. [PubMed] [CrossRef] [Google Scholar]
Биохим. Биофиз. Acta Proteins Proteom. 2012; 1824: 186–194. doi: 10.1016/j.bbapap.2011.07.001. [PubMed] [CrossRef] [Google Scholar]
33. Wei Y., Silke J.R., Aris P., Xia X. Геномы коронавирусов несут в себе признаки среды их обитания. ПЛОС ОДИН. 2020;15:e0244025. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Белузар С., Чу В.К., Уиттакер Г.Р. Активация спайкового белка коронавируса SARS посредством последовательного протеолитического расщепления в двух разных местах. проц. Натл. акад. науч. США. 2009; 106: 5871–5876. doi: 10.1073/pnas.0809524106. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Hsieh C.-L., Goldsmith J.A., Schaub J.M., DiVenere A.M., Kuo H.-C., Javanmardi K., Le K.C., Wrapp Д., Ли А.Г., Лю Ю. и др. Основанный на структуре дизайн шипов SARS-CoV-2, стабилизированных до слияния. Наука. 2020;369:1501. doi: 10.1126/science.abd0826. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
36. Симмонс Г., Госалия Д.Н., Реннекамп А. Дж., Ривз Дж.Д., Даймонд С.Л., Бейтс П. Ингибиторы катепсина L предотвращают проникновение коронавируса тяжелого острого респираторного синдрома. проц. Натл. акад. науч. США. 2005; 102:11876. doi: 10.1073/pnas.0505577102. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж., Ривз Дж.Д., Даймонд С.Л., Бейтс П. Ингибиторы катепсина L предотвращают проникновение коронавируса тяжелого острого респираторного синдрома. проц. Натл. акад. науч. США. 2005; 102:11876. doi: 10.1073/pnas.0505577102. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Bosch B.J., Bartelink W., Rottier P.J.M. Катепсин L функционально расщепляет гибридный белок коронавируса класса I тяжелого острого респираторного синдрома выше, а не рядом с гибридным пептидом. Дж. Вирол. 2008;82:8887. doi: 10.1128/ОВИ.00415-08. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
38. Burkard C., Verheije M.H., Wicht O., van Kasteren S.I., van Kuppeveld F.J., Haagmans B.L., Pelkmans L., Rottier P.J., Bosch B.J., de Haan C.A. Проникновение клеток коронавируса происходит через эндо-/лизосомальный путь зависимым от протеолиза образом. PLoS Патог. 2014;10:e1004502. doi: 10.1371/journal.ppat.1004502. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Kirschke H. Chapter 410—Cathepsin L. In: Rawlings ND, Salvesen G., editors. Справочник по протеолитическим ферментам. 3-е изд. Академическая пресса; Кембридж, Массачусетс, США: 2013. стр. 1808–1817. [Академия Google]
Kirschke H. Chapter 410—Cathepsin L. In: Rawlings ND, Salvesen G., editors. Справочник по протеолитическим ферментам. 3-е изд. Академическая пресса; Кембридж, Массачусетс, США: 2013. стр. 1808–1817. [Академия Google]
40. Джеймс Дж.А., Андре Н.М., Чаппи Дж.С., Милле Дж.К., Уиттакер Г.Р. Филогенетический анализ и структурное моделирование шиповидного белка SARS-CoV-2 выявили особую эволюционную и протеолитически чувствительную петлю активации. Дж. Мол. биол. 2020; 432:3309–3325. doi: 10.1016/j.jmb.2020.04.009. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
41. Мацуяма С., Нагата Н., Ширато К., Кавасэ М., Такеда М., Тагучи Ф. Эффективная активация тяжелого острого респираторного синдрома Спайковый белок коронавируса с помощью трансмембранной протеазы TMPRSS2. Дж. Вирол. 2010;84:12658. doi: 10.1128/ОВИ.01542-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Хоффманн М., Хофманн-Винклер Х., Пёльманн С. Время прайминга: как клеточные протеазы усиливают шиповидные белки коронавируса. В: Бёттхер-Фрибертсхойзер Э., Гартен В., Кленк Х.Д., редакторы. Активация вирусов протеазами-хозяевами. Международное издательство Спрингер; Чам, Швейцария: 2018. стр. 71–98. [Google Scholar]
43. Glowacka I., Bertram S., Müller M.A., Allen P., Soilleux E., Pfefferle S., Steffen I., Tsegaye T.S., He Y., Gnirss K., et al. Доказательства того, что TMPRSS2 активирует шиповидный белок коронавируса тяжелого острого респираторного синдрома для слияния мембран и снижает вирусный контроль за счет гуморального иммунного ответа. Дж. Вирол. 2011;85:4122–4134. doi: 10.1128/ОВИ.02232-10. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
44. Kleine-Weber H., Elzayat M.T., Hoffmann M., Pöhlmann S. Функциональный анализ потенциальных сайтов расщепления шиповидного белка MERS-коронавируса. науч. 2018; 8:16597. doi: 10.1038/s41598-018-34859-w. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Симмонс Г., Ривз Дж.Д., Реннекамп А.Дж., Амберг С.М., Пифер А. Дж., Бейтс П. CoV) опосредованное гликопротеином проникновение вируса. проц. Натл. акад. науч. США. 2004; 101:4240–4245. doi: 10.1073/pnas.0306446101. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж., Бейтс П. CoV) опосредованное гликопротеином проникновение вируса. проц. Натл. акад. науч. США. 2004; 101:4240–4245. doi: 10.1073/pnas.0306446101. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
46. Мацуяма С., Удзике М., Морикава С., Таширо М., Тагучи Ф. Опосредованное протеазами усиление тяжелого острого респираторного синдрома при коронавирусной инфекции. проц. Натл. акад. науч. США. 2005;102:12543. doi: 10.1073/pnas.0503203102. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
47. Peng G., Sun D., Rajashankar K.R., Qian Z., Holmes K.V., Li F. Кристаллическая структура комплексного домена, связывающего рецептор мышиного коронавируса со своим мышиным рецептором. проц. Натл. акад. науч. США. 2011;108:10696. doi: 10.1073/pnas.1104306108. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
48. Уильямс Р.К., Цзян Г.С., Холмс К.В. Рецептор вируса гепатита мыши является членом семейства гликопротеинов карциноэмбрионального антигена. проц. Натл. акад. науч. США. 1991;88:5533. doi: 10.1073/pnas.88.13.5533. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
проц. Натл. акад. науч. США. 1991;88:5533. doi: 10.1073/pnas.88.13.5533. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Xia X. DAMBE7: Новые и улучшенные инструменты для анализа данных в молекулярной биологии и эволюции. Мол. биол. Эвол. 2018;35:1550–1552. дои: 10.1093/молбев/msy073. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Gui M., Song W., Zhou H., Xu J., Chen S., Xiang Y., Wang X. Криоэлектронная микроскопия структуры шиповидного гликопротеина SARS-CoV обнаруживают необходимое конформационное состояние для связывания с рецептором. Сотовый рез. 2017;27:119–129. doi: 10.1038/cr.2016.152. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
51. Захарчук А.Н., Шарон С., Саткунараджа М., Ауперин Т., Вишванатан С., Мутвири Г., Петрик М., See R.H., Brunham R.C., Finlay B.B. и соавт. Иммуногенность рецептор-связывающего домена шиповидного белка коронавируса SARS у мышей: последствия для субъединичной вакцины. вакцина. 2007; 25: 136–143. doi: 10.1016/j.vaccine.2006.06.084. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
2007; 25: 136–143. doi: 10.1016/j.vaccine.2006.06.084. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. He Y., Zhou Y., Liu S., Kou Z., Li W., Farzan M., Jiang S. Рецептор-связывающий домен спайкового белка SARS-CoV индуцирует сильнодействующие нейтрализующие антитела: значение для развития субъединичная вакцина. Биохим. Биофиз. Рез. коммун. 2004; 324: 773–781. doi: 10.1016/j.bbrc.2004.09.106. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
53. He Y., Zhou Y., Siddiqui P., Jiang S. Инактивированная вакцина против SARS-CoV вызывает высокие титры специфичных к шиповидному белку антител, которые блокируют связывание с рецептором и проникновение вируса. Биохим. Биофиз. Рез. коммун. 2004; 325:445–452. doi: 10.1016/j.bbrc.2004.10.052. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
54. Du L., Zhao G., Chan C.C., Sun S., Chen M., Liu Z., Guo H., He Y., Zhou Y., Zheng B.J., et al. Рекомбинантный рецептор-связывающий домен шиповидного белка SARS-CoV, экспрессируемый в клетках млекопитающих, насекомых и кишечной палочки, вызывает сильнодействующие нейтрализующие антитела и защитный иммунитет. Вирусология. 2009; 393: 144–150. doi: 10.1016/j.virol.2009.07.018. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Вирусология. 2009; 393: 144–150. doi: 10.1016/j.virol.2009.07.018. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
55. Du L., Zhao G., He Y., Guo Y., Zheng B.J., Jiang S., Zhou Y. Рецепторно-связывающий домен Спайковый белок SARS-CoV индуцирует долгосрочный защитный иммунитет у животных моделей. вакцина. 2007; 25: 2832–2838. doi: 10.1016/j.vaccine.2006.10.031. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
56. Du L., Zhao G., Lin Y., Sui H., Chan C., Ma S., He Y., Jiang S., Wu C., Yuen K.Y., et al. Интраназальная вакцинация рекомбинантным аденоассоциированным вирусом, кодирующим шиповидный белок рецептор-связывающего домена коронавируса тяжелого острого респираторного синдрома (SARS-CoV), вызывает сильные иммунные реакции слизистых оболочек и обеспечивает долгосрочную защиту от инфекции SARS-CoV. Дж. Иммунол. 2008; 180:948–956. [Бесплатная статья PMC] [PubMed] [Google Scholar]
57. Zhang X., Wang J., Wen K., Mou Z., Zou L., Che X., Ni B. , Wu Y. Сайт связывания антител картирование домена, связывающего рецептор спайкового белка SARS-CoV, с помощью комбинации поверхностного дисплея дрожжей и скрининга библиотеки фаговых пептидов. Вирус Иммунол. 2009 г.;22:407–415. [PubMed] [Google Scholar]
, Wu Y. Сайт связывания антител картирование домена, связывающего рецептор спайкового белка SARS-CoV, с помощью комбинации поверхностного дисплея дрожжей и скрининга библиотеки фаговых пептидов. Вирус Иммунол. 2009 г.;22:407–415. [PubMed] [Google Scholar]
58. Cao Z., Liu L., Du L., Zhang C., Jiang S., Li T., He Y. Мощный и стойкий ответ антител против рецептор-связывающего домена Спайковый белок SARS-CoV у выздоровевших пациентов. Вирол. Дж. 2010; 7:299. [PMC free article] [PubMed] [Google Scholar]
59. Prabakaran P., Gan J., Feng Y., Zhu Z., Choudhry V., Xiao X., Ji X., Dimitrov D.S. Структура тяжелого острого Домен связывания рецептора коронавируса респираторного синдрома в комплексе с нейтрализующим антителом. Дж. Биол. хим. 2006;281:15829–15836. doi: 10.1074/jbc.M600697200. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
60. Chen WH, Du L., Chag S.M., Ma C., Tricoche N., Tao X., Seid C.A., Hudspeth EM, Lustigman S. , Tseng C.T., et al. Экспрессируемый дрожжами рекомбинантный белок рецептор-связывающего домена шиповидного белка SARS-CoV с дегликозилированными формами в качестве кандидата на вакцину против SARS. Гум. Вакцины Иммунотер. 2014; 10: 648–658. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Гум. Вакцины Иммунотер. 2014; 10: 648–658. [Статья бесплатно PMC] [PubMed] [Google Scholar]
61. Du L., Ma C., Jiang S. Антитела, индуцированные рецептор-связывающим доменом в спайковом белке SARS-CoV, не перекрестно нейтрализуют новый коронавирус человека. ВГС-ЭМС. Дж. Заразить. 2013;67:348–350. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Zhu Z., Chakraborti S., He Y., Roberts A., Sheahan T., Xiao X., Hensley L.E., Prabakaran P., Rockx B., Sidorov I.A., et al. Мощная перекрестно-реактивная нейтрализация изолятов коронавируса SARS человеческими моноклональными антителами. проц. Натл. акад. науч. США. 2007;104:12123–12128. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Elshabrawy H.A., Coughlin M.M., Baker S.C., Prabhakar B.S. Моноклональные антитела человека против высококонсервативных доменов HR1 и HR2 шиповидного белка SARS-CoV нейтрализуют в более широком смысле. ПЛОС ОДИН. 2012;7:e50366. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. He Y., Li J., Jiang S. Замена одной аминокислоты (R441A) в рецептор-связывающем домене шиповидного белка коронавируса SARS нарушает антигенную структуру и связывающую активность. Биохим. Биофиз. Рез. коммун. 2006; 344:106–113. doi: 10.1016/j.bbrc.2006.03.139. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
He Y., Li J., Jiang S. Замена одной аминокислоты (R441A) в рецептор-связывающем домене шиповидного белка коронавируса SARS нарушает антигенную структуру и связывающую активность. Биохим. Биофиз. Рез. коммун. 2006; 344:106–113. doi: 10.1016/j.bbrc.2006.03.139. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
65. Poh W.P., Narasaraju T., Pereira N.A., Zhong F., Phoon M.C., Macary P.A., Wong S.H., Lu J., Koh D.R., Chow В.Т. Характеристика эпитопов цитотоксических Т-лимфоцитов и иммунных ответов на ДНК-вакцину с шипами коронавируса SARS, экспрессирующую RGD-интегрин-связывающий мотив. Дж. Мед. Вирол. 2009 г.;81:1131–1139. doi: 10.1002/jmv.21571. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
66. Lin Y.S., Lin C.F., Fang Y.T., Kuo Y.M., Liao PC, Yeh TM, Hwa K.Y., Shieh C.C., Yen J.H., Wang H.J., et др. Антитело к домену 2 шиповидного белка коронавируса, связанного с тяжелым острым респираторным синдромом (SARS), перекрестно реагирует с эпителиальными клетками легких и вызывает цитотоксичность. клин. Эксп. Иммунол. 2005; 141: 500–508. doi: 10.1111/j.1365-2249.2005.02864.x. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
клин. Эксп. Иммунол. 2005; 141: 500–508. doi: 10.1111/j.1365-2249.2005.02864.x. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
67. Уайт Дж.М., Делос С.Е., Брехер М., Шорнберг К. Структуры и механизмы белков слияния вирусных мембран: многочисленные вариации на общую тему. крит. Преподобный Биохим. Мол. биол. 2008;43:189–219. doi: 10.1080/10409230802058320. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
68. Bosch B.J., Martina B.E., Van Der Zee R., Lepault J., Haijema B.J., Versluis C., Heck A.J., De Groot R., Остерхаус А. Д., Ротье П. Дж. Ингибирование инфекции коронавируса тяжелого острого респираторного синдрома (SARS-CoV) с использованием пептидов, полученных из гептадных повторов шиповидного белка. проц. Натл. акад. науч. США. 2004; 101:8455–8460. doi: 10.1073/pnas.0400576101. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
69. Броер Р., Босон Б., Спаан В., Коссет Ф.Л., Корвер Дж. Важная роль трансмембранного домена шиповидного белка коронавируса тяжелого острого респираторного синдрома во время проникновения. Дж. Вирол. 2006; 80: 1302–1310. doi: 10.1128/ОВИ.80.3.1302-1310.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Вирол. 2006; 80: 1302–1310. doi: 10.1128/ОВИ.80.3.1302-1310.2006. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
70. Modis Y. Слитые белки класса II. Доп. Эксп. Мед. биол. 2013; 790:150–166. [Бесплатная статья PMC] [PubMed] [Google Scholar]
71. Zeng X., Herndon A.M., Hu JC. Скрытые аспарагины определяют специфичность димеризации мутантов лейциновой молнии. проц. Натл. акад. науч. США. 1997;94:3673. doi: 10.1073/pnas.94.8.3673. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
72. Yoshida H., Oku M., Suzuki M., Mori K. pXBP1(U), закодированная в пре-мРНК XBP1, негативно регулирует развернутый активатор ответа белка pXBP1(S) в стрессовой реакции ER млекопитающих. Дж. Селл. биол. 2006; 172: 565–575. doi: 10.1083/jcb.200508145. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
73. Ся X. За пределами деревьев: регулоны и характеристика регуляторных мотивов. Гены. 2020;11:995. doi: 10.3390/genes110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
74. Chambers P., Pringle C.R., Easton A.J. Последовательности гептадных повторов расположены рядом с гидрофобными областями в нескольких типах гликопротеинов слияния вирусов. Дж. Генерал Вирол. 1990;71:3075–3080. doi: 10.1099/0022-1317-71-12-3075. [PubMed] [CrossRef] [Google Scholar]
Chambers P., Pringle C.R., Easton A.J. Последовательности гептадных повторов расположены рядом с гидрофобными областями в нескольких типах гликопротеинов слияния вирусов. Дж. Генерал Вирол. 1990;71:3075–3080. doi: 10.1099/0022-1317-71-12-3075. [PubMed] [CrossRef] [Google Scholar]
75. Basak S., Hao X., Chen A., Chrétien M., Basak A. Структурное и биохимическое исследование пептидов, полученных из гептадных повторов, вируса короны SARS человека (hSARS- CoV) шиповидный белок. Белковый пепт. лат. 2008; 15: 874–886. дои: 10.2174/092986608785849173. [PubMed] [CrossRef] [Google Scholar]
76. Xia S., Liu M., Wang C., Xu W., Lan Q., Feng S., Qi F., Bao L., Du L. ., Лю С. и др. Ингибирование инфекции SARS-CoV-2 (ранее 2019-nCoV) с помощью сильнодействующего ингибитора слияния панкоронавирусов, нацеленного на его шиповидный белок, который обладает высокой способностью опосредовать слияние мембран. Сотовый рез. 2020; 30: 343–355. doi: 10.1038/s41422-020-0305-x. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
77. Юань Ю., Цао Д., Чжан Ю., Ма Дж., Ци Дж., Ван Ц., Лу Г., Ву Ю. , Ян Дж., Ши Ю. и др. Крио-ЭМ-структуры шиповидных гликопротеинов MERS-CoV и SARS-CoV выявляют динамические домены связывания рецепторов. Нац. коммун. 2017;8:15092. doi: 10.1038/ncomms15092. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Юань Ю., Цао Д., Чжан Ю., Ма Дж., Ци Дж., Ван Ц., Лу Г., Ву Ю. , Ян Дж., Ши Ю. и др. Крио-ЭМ-структуры шиповидных гликопротеинов MERS-CoV и SARS-CoV выявляют динамические домены связывания рецепторов. Нац. коммун. 2017;8:15092. doi: 10.1038/ncomms15092. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
78. Ni L., Zhu J., Zhang J., Yan M., Gao G.F., Tien P. Дизайн SARS-CoV на основе рекомбинантного белка ингибиторы проникновения, нацеленные на области гептадных повторов S2-домена шиповидного белка. Биохим. Биофиз. Рез. коммун. 2005; 330:39–45. doi: 10.1016/j.bbrc.2005.02.117. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
79. Pallesen J., Wang N., Corbett K.S., Wrapp D., Kirchdoerfer R.N., Turner H.L., Cottrell C.A., Becker M.M., Wang L., Ши В. и др. Иммуногенность и структуры рационально сконструированного префузии спайкового антигена БВРС-КоВ. проц. Натл. акад. науч. США. 2017;114:E7348. doi: 10.1073/pnas.1707304114. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
80. Корвер Дж., Броер Р., ван Кастерен П., Спаан В. Мутагенез трансмембранного домена гликопротеина спайка коронавируса SARS: уточнение требований для проникновения в клетку коронавируса SARS. Вирол. Дж. 2009; 6:230. doi: 10.1186/1743-422X-6-230. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
Корвер Дж., Броер Р., ван Кастерен П., Спаан В. Мутагенез трансмембранного домена гликопротеина спайка коронавируса SARS: уточнение требований для проникновения в клетку коронавируса SARS. Вирол. Дж. 2009; 6:230. doi: 10.1186/1743-422X-6-230. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
81. Ляо Ю., Чжан С.М., Нео Т.Л., Там Дж.П. Триптофан-зависимое мембранное взаимодействие и гетеромеризация с внутренним слитым пептидом проксимальной внешней областью мембраны Спайковый белок SARS-CoV. Биохимия. 2015;54:1819–1830. дои: 10.1021/bi501352u. [PubMed] [CrossRef] [Google Scholar]
82. Пети С.М., Чульенко В.Н., Айер А., Колгроув Р., Фарзан М., Книпе Д.М., Кусулас К.Г. Пальмитоилирование богатого цистеином эндодомена гликопротеина шипа SARS-коронавируса важно для спайко-опосредованного слияния клеток. Вирусология. 2007; 360: 264–274. doi: 10.1016/j.virol.2006.10.034. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
83. Petit C.M., Melancon J.M., Chouljenko V. N., Colgrove R., Farzan M., Knipe D.M., Kousoulas K.G. Генетический анализ функциональных доменов гликопротеиновых шипов SARS-коронавируса, участвующих в экспрессии на клеточной поверхности и слиянии клеток с клетками. Вирусология. 2005; 341: 215–230. doi: 10.1016/j.virol.2005.06.046. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
N., Colgrove R., Farzan M., Knipe D.M., Kousoulas K.G. Генетический анализ функциональных доменов гликопротеиновых шипов SARS-коронавируса, участвующих в экспрессии на клеточной поверхности и слиянии клеток с клетками. Вирусология. 2005; 341: 215–230. doi: 10.1016/j.virol.2005.06.046. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
84. Милле Дж.К., Киен Ф., Чунг С.Ю., Сиу Ю.Л., Чан В.Л., Ли Х., Леунг Х.Л., Жауме М., Бруццоне Р., Пейрис Дж.С. и др. Эзрин взаимодействует с шиповидным белком коронавируса SARS и сдерживает инфекцию на начальной стадии. ПЛОС ОДИН. 2012;7:e49566. [Бесплатная статья PMC] [PubMed] [Google Scholar]
85. Anderson E.J., Rouphael N.G., Widge AT, Jackson L.A., Roberts PC, Makhene M., Chappell J.D., Denison M.R., Stevens L.J., Pruijssers A.J., et al. Безопасность и иммуногенность мРНК-вакцины SARS-CoV-2-1273 у пожилых людей. Н. англ. Дж. Мед. 2020; 383: 2427–2438. doi: 10.1056/NEJMoa2028436. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
86. Джексон Л.А., Андерсон Э.Дж., Руфаэль Н.Г., Робертс П.С., Махен М., Колер Р.Н., Маккалоу М.П., Чаппелл Дж.Д., Денисон М.Р., Стивенс Л.Дж. и др. мРНК-вакцина против SARS-CoV-2 — предварительный отчет. Н. англ. Дж. Мед. 2020; 383:1920–1931. doi: 10.1056/NEJMoa2022483. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Джексон Л.А., Андерсон Э.Дж., Руфаэль Н.Г., Робертс П.С., Махен М., Колер Р.Н., Маккалоу М.П., Чаппелл Дж.Д., Денисон М.Р., Стивенс Л.Дж. и др. мРНК-вакцина против SARS-CoV-2 — предварительный отчет. Н. англ. Дж. Мед. 2020; 383:1920–1931. doi: 10.1056/NEJMoa2022483. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
87. Denison M.R., Graham R.L., Donaldson E.F., Eckerle L.D., Baric R.S. Коронавирусы: машина для считывания РНК регулирует точность репликации и разнообразие. РНК биол. 2011; 8: 270–279.. doi: 10.4161/rna.8.2.15013. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
88. Ferron F., Subissi L., Silveira De Morais A.T., Le N.T.T., Sevajol M., Gluais L., Decroly E., Vonrhein C. , Bricogne G., Canard B., et al. Структурно-молекулярные основы коррекции несоответствия и удаления рибавирина из РНК коронавируса. проц. Натл. акад. науч. США. 2018;115:E162–E171. doi: 10.1073/pnas.1718806115. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
89. Робсон Ф., Хан К.С., Ле Т.К., Пэрис К., Демирбаг С., Барфусс П., Рокки П., Нг В.-Л. . Вычитка РНК коронавируса: молекулярная основа и терапевтическое нацеливание. Мол. Клетка. 2020;79: 710–727. doi: 10.1016/j.molcel.2020.07.027. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Робсон Ф., Хан К.С., Ле Т.К., Пэрис К., Демирбаг С., Барфусс П., Рокки П., Нг В.-Л. . Вычитка РНК коронавируса: молекулярная основа и терапевтическое нацеливание. Мол. Клетка. 2020;79: 710–727. doi: 10.1016/j.molcel.2020.07.027. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
90. Корбер Б., Фишер В.М., Гнанакаран С., Юн Х., Тайлер Дж., Абфальтерер В., Хенгартнер Н., Гиорги Э.Э., Бхаттачарья Т., Фоли Б. и др. Отслеживание изменений в пике SARS-CoV-2: доказательства того, что D614G повышает инфекционность вируса COVID-19. Клетка. 2020;182:812–827.e819. doi: 10.1016/j.cell.2020.06.043. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
91. Юрковецкий Л., Ван С., Паскаль К.Е., Томкинс-Тинч С., Ньялиле Т.П., Ван Ю., Баум А., Диль В.Е., Дофин А., Карбон К. и др. Структурно-функциональный анализ варианта спайкового белка D614G SARS-CoV-2. Клетка. 2020;183:739–751.e738. doi: 10.1016/j.cell.2020.09.032. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
92. Zhou H., Chen X., Hu T., Li J., Song H., Liu Y., Wang P., Liu D. , Yang J., Holmes E.C., et al. Новый коронавирус летучих мышей, тесно связанный с SARS-CoV-2, содержит естественные вставки в сайте расщепления S1/S2 шиповидного белка. Курс. биол. 2020;30:2196–2203.e3. doi: 10.1016/j.cub.2020.05.023. [Статья PMC бесплатно] [PubMed] [CrossRef] [Google Scholar]
Zhou H., Chen X., Hu T., Li J., Song H., Liu Y., Wang P., Liu D. , Yang J., Holmes E.C., et al. Новый коронавирус летучих мышей, тесно связанный с SARS-CoV-2, содержит естественные вставки в сайте расщепления S1/S2 шиповидного белка. Курс. биол. 2020;30:2196–2203.e3. doi: 10.1016/j.cub.2020.05.023. [Статья PMC бесплатно] [PubMed] [CrossRef] [Google Scholar]
93. Рейнке Л.М., Шпигель М., Плегге Т., Хартлейб А., Нельмайер И., Гиерер С., Хоффманн М., Хофманн-Винклер Х. ., Winkler M., Pöhlmann S. Различные остатки в шиповидном белке SARS-CoV определяют расщепление и активацию протеазой клетки-хозяина TMPRSS2. ПЛОС ОДИН. 2017;12:e0179177. [Бесплатная статья PMC] [PubMed] [Google Scholar]
94. Yao Y.X., Ren J., Heinen P., Zambon M., Jones I.M. Расщепление и сывороточная реактивность шиповидного белка тяжелого острого респираторного синдрома при коронавирусе. Дж. Заразить. Дис. 2004; 190:91–98. дои: 10.1086/421280. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
95. Xia X. Экстремальный дефицит геномного CpG при SARS-CoV-2 и уклонение от противовирусной защиты хозяина. Мол. биол. Эвол. 2020; 37: 2699–2705. doi: 10.1093/molbev/msaa094. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Xia X. Экстремальный дефицит геномного CpG при SARS-CoV-2 и уклонение от противовирусной защиты хозяина. Мол. биол. Эвол. 2020; 37: 2699–2705. doi: 10.1093/molbev/msaa094. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
96. Таката М.А., Гонсалвеш-Карнейро Д., Занг Т.М., Солл С.Дж., Йорк А., Бланко-Мело Д., Бениас П.Д. Подавление динуклеотидов CG позволяет противовирусной защите воздействовать на чужеродную РНК. Природа. 2017; 550:124–127. doi: 10.1038/nature24039. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
97. Лобри Дж. Р. Происхождение репликации Mycoplasma genitalium . Наука. 1996; 272: 745–746. doi: 10.1126/science.272.5262.745. [PubMed] [CrossRef] [Академия Google]
98. Kleine-Weber H., Pöhlmann S., Hoffmann M. Спайковые белки новых изолятов MERS-коронавируса от североафриканских и западноафриканских одногорбых верблюдов опосредуют надежное проникновение вируса в клетки-мишени человека. Вирусология. 2019; 535: 261–265. doi: 10.1016/j.virol.2019.07.016. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
doi: 10.1016/j.virol.2019.07.016. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
ACE2-независимая инфекция Т-лимфоцитов SARS-CoV-2
Введение
С момента своего появления в декабре 2019 года SARS-CoV-2, этиология коронавирусной болезни 2019(COVID-19), быстро распространились в большинстве стран мира и представляли большую угрозу для здоровья населения. Вирус имеет 79,5% геномную идентичность с SARS-CoV-1, а также использует ангиотензинпревращающий фермент 2 (ACE2) в качестве рецептора входа в клетку. 1,2,3,4,5 Типичные клинические симптомы пациентов с COVID-19 включают лихорадку, усталость, сухой кашель и пневмонию, тогда как около 20% тяжелых случаев могут умереть от полиорганной недостаточности. 6,7,8,9
Помимо дыхательной системы, многие органы, включая иммунную систему COVID-19пациенты также были заражены инфекцией SARS-CoV-2. Примечательно, что лимфопения наблюдалась у 83,2% пациентов при поступлении, а фатальные инфекции со временем ассоциировались с более тяжелой лимфопенией. 6,7,8 Лимфоциты (особенно Т-клетки) играют центральную роль в иммунной системе человека, снижение которой может привести к подавлению иммунитета и серьезным осложнениям. 10 Было высказано предположение, что вирусная лимфопения может быть связана с прямой инфекцией, опосредованной цитокинами гибелью клеток, тканевой секвестрацией лимфоцитов или подавлением образования Т-клеток костным мозгом или тимусом. 11 В случае БВРС-КоВ in vitro наблюдался апоптоз, вызванный прямой вирусной инфекцией Т-клеток, что, возможно, объясняет лимфопению у пациентов с БВРС. 11 Вирусные частицы SARS-CoV-1 также наблюдались в множественных лейкоцитах при вскрытии, что позволяет предположить, что прямое инфицирование может быть причиной снижения лимфоцитов. 12 Точно так же частицы или белки SARS-CoV-2 также были обнаружены в селезенке и лимфатических узлах при исследовании 91 умершего от COVID-19.случаи, предполагающие инфекцию лимфоцитов. 13 Кроме того, в иммунном ландшафте COVID-19, показанном в исследованиях секвенирования одноклеточной РНК, вирусная РНК SARS-CoV-2 была обнаружена во многих иммунных клетках, включая миелоидные клетки с фагоцитарной активностью (нейтрофилы и макрофаги) и лимфоциты без фагоцитарная активность (Т-, В- и NK-клетки).
6,7,8 Лимфоциты (особенно Т-клетки) играют центральную роль в иммунной системе человека, снижение которой может привести к подавлению иммунитета и серьезным осложнениям. 10 Было высказано предположение, что вирусная лимфопения может быть связана с прямой инфекцией, опосредованной цитокинами гибелью клеток, тканевой секвестрацией лимфоцитов или подавлением образования Т-клеток костным мозгом или тимусом. 11 В случае БВРС-КоВ in vitro наблюдался апоптоз, вызванный прямой вирусной инфекцией Т-клеток, что, возможно, объясняет лимфопению у пациентов с БВРС. 11 Вирусные частицы SARS-CoV-1 также наблюдались в множественных лейкоцитах при вскрытии, что позволяет предположить, что прямое инфицирование может быть причиной снижения лимфоцитов. 12 Точно так же частицы или белки SARS-CoV-2 также были обнаружены в селезенке и лимфатических узлах при исследовании 91 умершего от COVID-19.случаи, предполагающие инфекцию лимфоцитов. 13 Кроме того, в иммунном ландшафте COVID-19, показанном в исследованиях секвенирования одноклеточной РНК, вирусная РНК SARS-CoV-2 была обнаружена во многих иммунных клетках, включая миелоидные клетки с фагоцитарной активностью (нейтрофилы и макрофаги) и лимфоциты без фагоцитарная активность (Т-, В- и NK-клетки). 14,15 Примечательно, что РНК-положительные иммунные клетки SARS-CoV-2 не экспрессировали одновременно факторы входа ACE2 и TMPRSS2 или другие предполагаемые кофакторы входа. 14,15 Предполагается, что ассоциированная с клетками положительная реакция на вирус SARS-CoV-2 может представлять собой смесь репликации вируса, поглощения иммунными клетками и вирионов или инфицированных вирусом клеток, прикрепленных к клеточной поверхности. 14,15
14,15 Примечательно, что РНК-положительные иммунные клетки SARS-CoV-2 не экспрессировали одновременно факторы входа ACE2 и TMPRSS2 или другие предполагаемые кофакторы входа. 14,15 Предполагается, что ассоциированная с клетками положительная реакция на вирус SARS-CoV-2 может представлять собой смесь репликации вируса, поглощения иммунными клетками и вирионов или инфицированных вирусом клеток, прикрепленных к клеточной поверхности. 14,15
Было показано, что SARS-CoV-2 инфицирует человеческие моноциты, моноцитарные макрофаги и дендритные клетки in vitro, что потенциально играет важную роль в патогенезе COVID-19. 16,17 Однако до сих пор неизвестно, заражает ли SARS-CoV-2 лимфоциты, которые не экспрессируют ACE2, приводя к лимфопении. Этот пробел в знаниях также затрудняет наше понимание того, как лимфоциты утратили способность контролировать вирусную инфекцию. Здесь мы представили доказательства того, что активированные Т-лимфоциты могут быть инфицированы SARS-CoV-2 независимым от ACE2 образом. Инфекция приводит к выраженному апоптозу Т-клеток in vitro или у пациентов с COVID-19.. Наши результаты проливают свет на понимание лимфопении, вызванной инфекцией SARS-CoV-2.
Инфекция приводит к выраженному апоптозу Т-клеток in vitro или у пациентов с COVID-19.. Наши результаты проливают свет на понимание лимфопении, вызванной инфекцией SARS-CoV-2.
Результаты
Присутствие SARS-CoV-2 в лимфоцитах пациентов с COVID-19
Было показано, что несколько типов иммунных клеток, включая лимфоциты, обогащены вирусной РНК SARS-CoV-2 в нескольких одноклеточных РНК-последовательностях исследования. 14,15 Чтобы определить, инфицирует ли SARS-CoV-2 лимфоциты, мы проанализировали клетки периферической крови (ПБК), взятые у пациентов с COVID-19. ПБЦ были получены от 22 пациентов, которые во время исследования находились в тяжелом состоянии, а также от 15 здоровых доноров. Сначала мы проанализировали основные типы лимфоцитов, включая Т (CD4 + хелперные Т и CD8 + цитотоксические Т), В и естественные клетки-киллеры (NK) на предмет изменений их популяции или присутствия вирусного антигена при инфицировании. У всех протестированных пациентов соотношение Т-лимфоцитов крови значительно снизилось по сравнению с таковым у здоровых доноров, тогда как В- и NK-клетки не изменились (рис. 1а). Примечательно, что у некоторых пациентов количество CD4 + и CD8 + Т-лимфоцитов почти снизилось до нуля (рис. 1б). Результаты показали, что лимфопения у этих пациентов, вероятно, связана со снижением Т-лимфоцитов.
1а). Примечательно, что у некоторых пациентов количество CD4 + и CD8 + Т-лимфоцитов почти снизилось до нуля (рис. 1б). Результаты показали, что лимфопения у этих пациентов, вероятно, связана со снижением Т-лимфоцитов.
Лимфоциты периферической крови инфицированы SARS-CoV-2 у пациентов с COVID-19. a Процент различных типов лимфоцитов у здоровых доноров ( n = 15) или у пациентов с COVID-19 ( n = 22). ( b ) Процент CD4 + и CD8 + Т-лимфоцитов у здорового донора ( n = 8) или у пациентов с COVID-19 ( n = 9). c , d Иммунофлюоресцентный тест на наличие вирусного антигена SARS-CoV-2 в Т-клетках. КПБ ( c ) или посмертный срез легкого ( d ) пациентов с COVID-19 окрашивали Т-лимфоцитами (CD3, зеленый), SARS-CoV-2 (NP, красный) и ядрами (DAPI, синий). Использовали pAb собственного производства против NP SARS-CoV-2. Белые стрелки указывают области Т-лимфоцитов, которые были инфицированы SARS-CoV-2. Снимки сделаны под конфокальной микроскопией с полосой = 50 мкм ( c ) или 200 мкм ( d ). Сравнение средних значений ( a , b ) между двумя группами было проанализировано с помощью критерия Стьюдента.0249 т тест. * P < 0,05; ** P < 0,01; **** P < 0,0001; NS не имеет значения
Снимки сделаны под конфокальной микроскопией с полосой = 50 мкм ( c ) или 200 мкм ( d ). Сравнение средних значений ( a , b ) между двумя группами было проанализировано с помощью критерия Стьюдента.0249 т тест. * P < 0,05; ** P < 0,01; **** P < 0,0001; NS не имеет значения
Изображение в полный размер
Затем мы проанализировали наличие вирусных антигенов SARS-CoV-2 в ПБЦ с помощью проточной цитометрии или иммунофлуоресцентного анализа (ИФА). Результаты показали, что Т-лимфоциты были инфицированы, и у некоторых пациентов CD4 + Т-клетки показали высокий уровень инфицирования (дополнительная рис. S1a). Мы также подтвердили наличие вирусного антигена в Т-лимфоцитах крови пациентов с помощью иммунофлуоресцентного анализа (ИФА) (рис. 1в). Кроме того, мы подготовили посмертные срезы легких пациентов со смертельной инфекцией и проанализировали инфильтрацию Т-лимфоцитов и вирусную инфекцию. Мы обнаружили инфильтрацию Т-лимфоцитов в срезе легкого, и многие Т-лимфоциты также были положительными при окрашивании NP SARS-CoV-2, что указывает на вирусную инфекцию (рис. 1d). Сообщалось также об аналогичной находке. 13 В совокупности мы показали наличие вирусного антигена SARS-CoV-2 в Т-лимфоцитах либо в крови, либо в срезе легких у пациентов с COVID-19.
1d). Сообщалось также об аналогичной находке. 13 В совокупности мы показали наличие вирусного антигена SARS-CoV-2 в Т-лимфоцитах либо в крови, либо в срезе легких у пациентов с COVID-19.
Для дальнейшего подтверждения этих результатов была предпринята попытка выделения вируса из вирусных NP-позитивных ПБЦ. PBC пациентов собирали, определяли на вирусный антиген с помощью проточной цитометрии, а затем совместно культивировали с клетками Caco2 после трех промывок. Положительное обнаружение вирусной РНК в супернатанте или вирусного белка в клетках Caco2 после совместного культивирования свидетельствует об успешном выделении и амплификации SARS-CoV-2 из PBC некоторых COVID-19.пациентов (3 из 5), но не из здорового контроля (дополнительная рис. S1b – e). Примечательно, что в трех образцах, положительных на выделение вируса, два также показали положительный результат на вирус в анализе проточной цитометрии (P2 и P4), в то время как третий (P5), вероятно, содержал инфекционный вирус на уровне, который был ниже предела обнаружения проточного цитометрического анализа. . Прежде всего, мы наблюдали вирусную РНК и вирусный белок SAR-CoV-2, а также, вероятно, инфекционный вирус в Т-лимфоцитах пациентов с COVID-19.
. Прежде всего, мы наблюдали вирусную РНК и вирусный белок SAR-CoV-2, а также, вероятно, инфекционный вирус в Т-лимфоцитах пациентов с COVID-19.
Заражение SARS-CoV-2 Т-клеток in vitro
Поскольку популяция Т-лимфоцитов уменьшилась у пациентов с COVID-19, а CD4 + Т-лимфоциты демонстрировали высокий уровень положительной реакции на вирусный антиген, мы затем исследовали, инфицирует ли SARS-CoV-2 CD4 + Т-лимфоциты. С этой целью мы провели серию экспериментов, чтобы проверить, заражает ли SARS-CoV-2 Т-клетки. После инфицирования тестировались как обнаружение вирусной РНК, нацеленной на рецептор-связывающий домен (RBD), так и вирусная субгеномная мРНК (sgRNA), направленная на ген М. Вирусная sgRNA транскрибируется только в инфицированных клетках во время репликации вируса и не упаковывается в вирионы, что свидетельствует о наличии в образцах активно инфицированных клеток. Вирусный нуклеокапсидный белок (НП) и вирусные частицы также выявляли с помощью вестерн-блоттинга (ВБ), проточной или электронной микроскопии (ЭМ). Клетки Jurkat или MT4, две обычно используемые линии CD4 + Т-клеток и первичные Т-клетки, выделенные от здоровых доноров, были инфицированы SARS-CoV-2 (рис. 2а). В некоторых экспериментах Т-клетки также активировали форболмиристатацетатом (PMA) в течение 2 часов для клеток Jurkat или комбинацией IL2++CD3++CD28 в течение 3 дней для первичных Т-клеток до инфицирования, учитывая, что большая часть Т-клеток активирован у человека (дополнительный рис. S2).
Клетки Jurkat или MT4, две обычно используемые линии CD4 + Т-клеток и первичные Т-клетки, выделенные от здоровых доноров, были инфицированы SARS-CoV-2 (рис. 2а). В некоторых экспериментах Т-клетки также активировали форболмиристатацетатом (PMA) в течение 2 часов для клеток Jurkat или комбинацией IL2++CD3++CD28 в течение 3 дней для первичных Т-клеток до инфицирования, учитывая, что большая часть Т-клеток активирован у человека (дополнительный рис. S2).
Заражение SARS-CoV-2 in vitro линий Т-клеток или первичных Т-клеток. a Расписание экспериментов. b Неактивированные или активированные клетки Jurkat были инфицированы SARS-CoV-2 (MOI = 0,1), и образцы были собраны через 0, 24, 48 и 72 часа после заражения. Вирусную нагрузку в клетках или клеточном супернатанте затем количественно определяли с помощью количественной ПЦР, обнаруживая общую вирусную РНК (RBD спайкового гена в качестве мишени) или субгеномную РНК (sgRNA, ген М в качестве мишени). c Сравнение глубины и покрытия для активированных клеток Jurkat, инфицированных SARS-CoV-2 через 0 или 24 часа. Для каждого образца количество вирусов нормализовали по общему количеству прочтений секвенирования. Два повторения показаны для каждой временной точки. d Вирусные НЧ в инфицированных активированных клетках Jurkat и клеточном супернатанте анализировали с помощью вестерн-блоттинга через 0, 24, 48 и 72 ч после инфицирования. e Клетки, инфицированные вирусом NP 72 h из ( d ), были проанализированы с помощью проточной цитометрии, и количество повторов, представленных на гистограмме, равно трем. f Вирусные частицы в инфицированных активированных клетках Jurkat или MT4 наблюдали с помощью просвечивающего электронного микроскопа. bar = 1 мкм или 500 нм, увеличение: 3500-кратное и 9600-кратный для ячейки Jurkat, 5000-кратный или 11500-кратный для ячейки MT4. г Неактивированные или активированные первичные Т-клетки были инфицированы SARS-CoV-2 (MOI = 0,01), и образцы были собраны через 0, 4, 8 и 12 часов после заражения.
c Сравнение глубины и покрытия для активированных клеток Jurkat, инфицированных SARS-CoV-2 через 0 или 24 часа. Для каждого образца количество вирусов нормализовали по общему количеству прочтений секвенирования. Два повторения показаны для каждой временной точки. d Вирусные НЧ в инфицированных активированных клетках Jurkat и клеточном супернатанте анализировали с помощью вестерн-блоттинга через 0, 24, 48 и 72 ч после инфицирования. e Клетки, инфицированные вирусом NP 72 h из ( d ), были проанализированы с помощью проточной цитометрии, и количество повторов, представленных на гистограмме, равно трем. f Вирусные частицы в инфицированных активированных клетках Jurkat или MT4 наблюдали с помощью просвечивающего электронного микроскопа. bar = 1 мкм или 500 нм, увеличение: 3500-кратное и 9600-кратный для ячейки Jurkat, 5000-кратный или 11500-кратный для ячейки MT4. г Неактивированные или активированные первичные Т-клетки были инфицированы SARS-CoV-2 (MOI = 0,01), и образцы были собраны через 0, 4, 8 и 12 часов после заражения. Затем вирусную нагрузку в клетках определяли количественно с помощью количественной ПЦР. ч Органоиды толстой кишки были инфицированы SARS-CoV-2 (MOI = 0,01). Образцы нулевого часа и 24 часов были собраны и количественно оценены с помощью количественной ПЦР. Данные были проанализированы с помощью теста Стьюдента t , и статистическая значимость указана звездочками (* P < 0,05; ** P < 0,01; **** P < 0,0001; NS не имеет значения)
Затем вирусную нагрузку в клетках определяли количественно с помощью количественной ПЦР. ч Органоиды толстой кишки были инфицированы SARS-CoV-2 (MOI = 0,01). Образцы нулевого часа и 24 часов были собраны и количественно оценены с помощью количественной ПЦР. Данные были проанализированы с помощью теста Стьюдента t , и статистическая значимость указана звездочками (* P < 0,05; ** P < 0,01; **** P < 0,0001; NS не имеет значения)
Полноразмерное изображение
Через 0, 24, 48 и 72 часа после заражения было замечено, что SARS-CoV-2-инфицированная Т-клеточная линия Jurkat в зависимости от времени, и инфекция был более устойчивым в активированных Т-клетках. Наблюдали накопление вирусной РНК и sgRNA в клетках или вирусной РНК в культуральном супернатанте (рис. 2б). Затем мы попытались определить, представляет ли анализ обнаружения qPCR только частичную репликацию вирусного генома. Мы провели анализ секвенирования РНК активированных Т-клеток Jurkat, инфицированных SARS-CoV-2, через 0 или 24 часа после заражения. и проанализировали глубину вирусных прочтений и охват вирусного генома. По сравнению с зараженными через 0 часов, в клетках, инфицированных через 24 часа, можно наблюдать гораздо большую глубину вирусных геномов (до 5000 прочтений), что демонстрирует эффективную репликацию (рис. 2с). Затем мы определили вирусные антигены с помощью WB и проточного анализа. Наши результаты показали зависящее от времени увеличение уровня вирусных NP в клетках или в супернатанте, аналогично результатам обнаружения вирусной РНК (рис. 2d, e). Далее мы использовали электронную микроскопию для анализа инфекции Т-клеточных линий SARS-CoV-2. Активированные клетки Jurkat или MT4 были инфицированы SARS-CoV-2 в течение 72 часов, и в цитоплазме инфицированных клеток наблюдались вирусные частицы с типичной для коронавируса морфологией (рис. 2f). Наконец, чтобы подтвердить выводы из линий Т-клеток, мы проверили инфекционность первичных Т-клеток, выделенных от здоровых доноров. У трех доноров SARS-CoV-2 продемонстрировал зависимое от времени инфицирование Т-клеток с пиком через 8 часов, вероятно, из-за обширной гибели клеток, вызванной вирусом в этот момент времени (обсуждается ниже).
и проанализировали глубину вирусных прочтений и охват вирусного генома. По сравнению с зараженными через 0 часов, в клетках, инфицированных через 24 часа, можно наблюдать гораздо большую глубину вирусных геномов (до 5000 прочтений), что демонстрирует эффективную репликацию (рис. 2с). Затем мы определили вирусные антигены с помощью WB и проточного анализа. Наши результаты показали зависящее от времени увеличение уровня вирусных NP в клетках или в супернатанте, аналогично результатам обнаружения вирусной РНК (рис. 2d, e). Далее мы использовали электронную микроскопию для анализа инфекции Т-клеточных линий SARS-CoV-2. Активированные клетки Jurkat или MT4 были инфицированы SARS-CoV-2 в течение 72 часов, и в цитоплазме инфицированных клеток наблюдались вирусные частицы с типичной для коронавируса морфологией (рис. 2f). Наконец, чтобы подтвердить выводы из линий Т-клеток, мы проверили инфекционность первичных Т-клеток, выделенных от здоровых доноров. У трех доноров SARS-CoV-2 продемонстрировал зависимое от времени инфицирование Т-клеток с пиком через 8 часов, вероятно, из-за обширной гибели клеток, вызванной вирусом в этот момент времени (обсуждается ниже). Активация сделала клетки чувствительными к инфекции SARS-CoV-2 у двух из трех доноров. Для сравнения, был также инфицирован первичный органоид толстой кишки, который показал гораздо более высокую эффективность инфекции по сравнению с Т-клетками (рис. 2ж, з). В совокупности наши данные ясно показывают, что SARS-CoV-2 может инфицировать Т-клетки in vitro, хотя и с меньшей эффективностью по сравнению с тканевыми клетками.
Активация сделала клетки чувствительными к инфекции SARS-CoV-2 у двух из трех доноров. Для сравнения, был также инфицирован первичный органоид толстой кишки, который показал гораздо более высокую эффективность инфекции по сравнению с Т-клетками (рис. 2ж, з). В совокупности наши данные ясно показывают, что SARS-CoV-2 может инфицировать Т-клетки in vitro, хотя и с меньшей эффективностью по сравнению с тканевыми клетками.
Инфекция Т-клеток SARS-CoV-2 не зависит от ACE2 и TMPRSS2
Обычно считается, что ACE2 является входным рецептором для SARS-CoV-2. Однако основные клеточные популяции в ПБЦ экспрессируют крайне низкие уровни ACE2, что ставит вопрос о том, опосредует ли ACE2 также проникновение вируса SARS-CoV-2 в Т-клетки. Сначала мы проверили, может ли нокдаун ACE2 ослабить инфекцию SARS-CoV-2 Т-клеток. Данные показали, что ACE2 был успешно сбит с помощью ACE2-shRNAs в клетках Caco2. Т-клетки Jurkat не экспрессируют обнаруживаемый ACE2 ни в фиктивных, ни в нокдаун-условиях (рис. 3а). Соответственно, нокдаун ACE2 привел к резкому снижению инфекции SARS-CoV-2 в клетках Caco2, но не в T-клетках Jurkat (рис. 3b). Чтобы дополнительно подтвердить это открытие, мы нокаутировали ACE2 в клетках Caco2 и Jurkat (рис. 3c). Подобно клеткам с нокдауном ACE2, вирусная нагрузка снижалась в клетках Caco2-ACE2-KO, но не в клетках Jurkat-ACE2-KO (рис. 3d). Эти результаты свидетельствуют о том, что Т-клетки, инфицированные SARS-CoV-2, независимы от АПФ2.
3а). Соответственно, нокдаун ACE2 привел к резкому снижению инфекции SARS-CoV-2 в клетках Caco2, но не в T-клетках Jurkat (рис. 3b). Чтобы дополнительно подтвердить это открытие, мы нокаутировали ACE2 в клетках Caco2 и Jurkat (рис. 3c). Подобно клеткам с нокдауном ACE2, вирусная нагрузка снижалась в клетках Caco2-ACE2-KO, но не в клетках Jurkat-ACE2-KO (рис. 3d). Эти результаты свидетельствуют о том, что Т-клетки, инфицированные SARS-CoV-2, независимы от АПФ2.
Заражение Т-клеток SARS-CoV-2 не зависит от спайк-ACE2/TMPRSS2. a Уровень экспрессии ACE2 в клетках Scramble или клетках Caco2 или Jurkat с нокдауном ACE2 анализировали с помощью количественной ПЦР или WB. b ACE2 со стабильным нокдауном Caco2 или активированные клетки Jurkat были инфицированы SARS-CoV-2 (MOI = 0,01). Вирусную РНК или вирусный NP в клетках анализировали с помощью количественной ПЦР или WB через 24 часа после инфицирования. c Уровень экспрессии ACE2 в контрольных или нокаутных по ACE клетках Caco2 или Jurkat определяли количественно с помощью количественной ПЦР или определяли с помощью WB. d клетки Caco2 со стабильным нокаутом ACE2 или активированные клетки Jurkat были инфицированы SARS-CoV-2 (MOI = 0,01). Вирусную РНК или вирусный NP выявляли с помощью qPCR или WB. e Для блокирования ACE2 клетки Caco2 или активированные клетки Jurkat предварительно инкубировали с антителами против ACE2 (конечная концентрация 3,33 нг/мкл) перед инфицированием SARS-CoV-2. Для блокирования вируса белок ACE2-Fc (10 мкг/мкл, конечный результат) или RD#4-анти-Spike Ab (160 нг/мкл, конечный) инкубировали с SARS-CoV-2 в объеме 1:1 при 37 °C. на 30 мин. Клетки инфицировали при 0,01 MOI в течение 24 часов, прежде чем они были количественно определены на вирусную РНК SARS-CoV-2 или белок NP с помощью количественной ПЦР или WB. f Уровень экспрессии TMPRSS2 в клетках Caco2, Jurkat и активированных клетках Jurkat анализировали с использованием количественной ПЦР. г клеток Caco2 или активированных клеток Jurkat предварительно инкубировали с камостатом (2 мкМ или 20 мкМ) в течение 1 часа, а затем инфицировали SARS-CoV-2 (MOI = 0,01).
d клетки Caco2 со стабильным нокаутом ACE2 или активированные клетки Jurkat были инфицированы SARS-CoV-2 (MOI = 0,01). Вирусную РНК или вирусный NP выявляли с помощью qPCR или WB. e Для блокирования ACE2 клетки Caco2 или активированные клетки Jurkat предварительно инкубировали с антителами против ACE2 (конечная концентрация 3,33 нг/мкл) перед инфицированием SARS-CoV-2. Для блокирования вируса белок ACE2-Fc (10 мкг/мкл, конечный результат) или RD#4-анти-Spike Ab (160 нг/мкл, конечный) инкубировали с SARS-CoV-2 в объеме 1:1 при 37 °C. на 30 мин. Клетки инфицировали при 0,01 MOI в течение 24 часов, прежде чем они были количественно определены на вирусную РНК SARS-CoV-2 или белок NP с помощью количественной ПЦР или WB. f Уровень экспрессии TMPRSS2 в клетках Caco2, Jurkat и активированных клетках Jurkat анализировали с использованием количественной ПЦР. г клеток Caco2 или активированных клеток Jurkat предварительно инкубировали с камостатом (2 мкМ или 20 мкМ) в течение 1 часа, а затем инфицировали SARS-CoV-2 (MOI = 0,01). Вирусную РНК или белки NP определяли количественно. Результаты были получены из трех независимых экспериментов. Статистический анализ проводили с использованием критерия Стьюдента t (* P < 0,05; ** P < 0,01; **** P < 0,0001; NS не имеет значения)
Вирусную РНК или белки NP определяли количественно. Результаты были получены из трех независимых экспериментов. Статистический анализ проводили с использованием критерия Стьюдента t (* P < 0,05; ** P < 0,01; **** P < 0,0001; NS не имеет значения)
Изображение в полный размер
Сообщалось, что растворимый человеческий белок ACE2 может блокировать инфекцию SARS-CoV-2 за счет конкурирующего связывания вируса с клеточным рецептором. 3 Таким образом, предварительно инкубированные клетки с антителом ACE2 или предварительно инкубированные с шипом антитела к SARS-CoV-2 также должны блокировать вирусную инфекцию, если инфекция зависит от связывания шипа с ACE2. Чтобы проанализировать, влияют ли эти молекулы на инфицирование Т-клеток SARS-CoV-2, мы инкубировали вирус с растворимым человеческим белком ACE2 или коммерческим mAb, нацеленным на связывание RBD-ACE2, или инкубировали клетки с блокирующим ACE2 антителом до заражения Caco2 или активированного Jurkat. Т-клетки. Внутриклеточную вирусную РНК анализировали после инфицирования. В Caco2 три блокатора сильно блокировали инфекцию SARS-CoV-2, а белок ACE2 оказался более эффективным, чем два других препарата. Напротив, ни один из трех видов лечения не повлиял на инфекцию SARS-CoV-2 T-клеток Jurkat (рис. 3e).
Т-клетки. Внутриклеточную вирусную РНК анализировали после инфицирования. В Caco2 три блокатора сильно блокировали инфекцию SARS-CoV-2, а белок ACE2 оказался более эффективным, чем два других препарата. Напротив, ни один из трех видов лечения не повлиял на инфекцию SARS-CoV-2 T-клеток Jurkat (рис. 3e).
Наконец, известно, что SARS-CoV-2 использует сериновую протеазу TMPRSS2 для праймирования S-белка перед связыванием с рецептором ACE2, а ингибитор TMPRSS2 был одобрен для клинического применения (Camostat) для блокирования проникновения SARS-CoV-2. 1 Экспрессию РНК TMPRSS2 в клетках Caco2, Jurkat и активированных клетках Jurkat определяли с помощью количественной ПЦР. Результат показал, что ни неактивированные, ни активированные клетки Jurkat не экспрессируют TMPRSS2 (рис. 3f). Мы наблюдали, что камостат дозозависимым образом ингибировал заражение SARS-CoV-2 клеток Caco2. Камостат в дозе 20 мкм практически полностью блокировал вирусную инфекцию клеток Caco2. Напротив, камостат не проявлял ингибирующего действия на инфекцию SARS-CoV-2 T-клеток Jurkat даже в высокой дозе (рис. 3g). В совокупности эти результаты свидетельствуют о том, что заражение Т-клеток SARS-CoV-2 не зависит от взаимодействия шип-ACE2/TMPRSS2.
3g). В совокупности эти результаты свидетельствуют о том, что заражение Т-клеток SARS-CoV-2 не зависит от взаимодействия шип-ACE2/TMPRSS2.
Инфекция SARS-CoV-2 вызвала гибель Т-клеток
Известно, что у тяжелых пациентов с COVID-19 наблюдается заметное снижение популяций лимфоцитов. Чтобы определить, способствует ли инфекция SARS-CoV-2 гибели Т-клеток, мы протестировали апоптоз Т-лимфоцитов ПБЦ, собранных у пациентов с COVID-19. Т-лимфоциты пациентов или здоровых доноров были дважды помечены антителом к CD3 и антителом к вирусному NP, а апоптоз был проанализирован с помощью анализа TUNEL. Т-лимфоциты от COVID-19пациенты подверглись выраженному апоптозу по сравнению со здоровыми донорами, показывая более чем десятикратное увеличение апоптотических клеток. У некоторых пациентов большинство апоптотических клеток также были положительными по вирусному антигену (например, 65% у пациента 1), что позволяет предположить, что вирусная инфекция играла роль в гибели Т-лимфоцитов периферической крови у этих пациентов (рис. 4а).
4а).
Апоптоз Т-клеток, вызванный инфекцией SARS-CoV-2. a Обнаружение апоптотических Т-лимфоцитов в ПБЦ человека. КПБ были приготовлены из COVID-19больных или от здоровых доноров. Апоптоз в инфицированных вирусом Т-лимфоцитах определяли с использованием CD3, антител к NP SARS-CoV-2 и анализа TUNEL. Для здоровых доноров (H, n = 3) или пациентов (P, n = 3) или пациентов (P, n = 5) были показаны результаты детекции для двух пациентов и одного здорового донора, либо статистика апоптотических клеток или TUNEL/NP-двойных положительных клеток. Сравнение средних значений между двумя группами анализировали с помощью теста Стьюдента t . * P < 0,05. б Неактивированные или активированные первичные Т-клетки инфицировали SARS-CoV-2 (MOI = 0,01) в течение 8 часов и анализировали клеточный апоптоз с помощью анализа TUNEL. ** P < 0,01. c Активированные клетки Jurkat были инфицированы SARS-CoV-2 (MOI = 0,01) в течение 0, 24, 48 и 72 часов. Было показано соотношение апоптотических клеток в 72-часовых имитационных или инфицированных SARS-CoV-2 клетках (блок-схема). Соотношение апоптотических клеток в разные моменты времени также сравнивали (график). d Образцы из C были подвергнуты анализу секвенирования РНК. Показаны десять основных путей активации ГО через 48 часов по сравнению с группой 24 часов. Тепловая карта показывает нормализованную экспрессию генов, которые были обогащены путем PID HIF1. Данные были проанализированы Стьюдентом t тест (* P < 0,05; ** P < 0,01; NS не имеет значения). e По сравнению со здоровыми донорами, дифференциально экспрессированные e генов пациентов с тяжелым течением COVID-19 были показаны на графике вулкана, а первые десять дифференциально экспрессированных путей были показаны на правой панели
Было показано соотношение апоптотических клеток в 72-часовых имитационных или инфицированных SARS-CoV-2 клетках (блок-схема). Соотношение апоптотических клеток в разные моменты времени также сравнивали (график). d Образцы из C были подвергнуты анализу секвенирования РНК. Показаны десять основных путей активации ГО через 48 часов по сравнению с группой 24 часов. Тепловая карта показывает нормализованную экспрессию генов, которые были обогащены путем PID HIF1. Данные были проанализированы Стьюдентом t тест (* P < 0,05; ** P < 0,01; NS не имеет значения). e По сравнению со здоровыми донорами, дифференциально экспрессированные e генов пациентов с тяжелым течением COVID-19 были показаны на графике вулкана, а первые десять дифференциально экспрессированных путей были показаны на правой панели
Полноразмерное изображение
Для подтверждения Роль вирусной инфекции в гибели Т-клеток мы экспериментально инфицировали первичными Т-клетками, выделенными от здоровых доноров. С активацией или без нее клетки экспериментально инфицировали SARS-CoV-2 в течение 8 часов и анализировали апоптоз с помощью анализа TUNEL. Можно заметить, что инфекция SARS-CoV-2 индуцировала выраженный апоптоз в инфицированных Т-клетках по сравнению с ложнообработанными клетками. Активация сенсибилизирует Т-клетки к вирусной инфекции, о чем свидетельствуют более высокие апоптотические клетки в активированной группе (фиг. 4b).
С активацией или без нее клетки экспериментально инфицировали SARS-CoV-2 в течение 8 часов и анализировали апоптоз с помощью анализа TUNEL. Можно заметить, что инфекция SARS-CoV-2 индуцировала выраженный апоптоз в инфицированных Т-клетках по сравнению с ложнообработанными клетками. Активация сенсибилизирует Т-клетки к вирусной инфекции, о чем свидетельствуют более высокие апоптотические клетки в активированной группе (фиг. 4b).
Наконец, мы определили клеточные ответы в Т-клетках при заражении SARS-CoV-2 с помощью анализа секвенирования РНК. Активированные Т-клетки Jurkat были инфицированы SARS-CoV-2 в течение 0, 24, 48 и 72 ч, прежде чем они были собраны для анализа TUNEL. Можно заметить, что вирус индуцировал значительный апоптоз через 72 часа после заражения по сравнению с ложно инфицированными клетками или клетками в другие моменты времени (рис. 4с). Затем мы определили динамические клеточные ответы в клетках, которые были инфицированы в течение 24 или 48 часов, поскольку клетки в 72-часовых группах содержали слишком много мертвых клеток и не подходили для анализа секвенирования РНК. По сравнению с группой 24 ч, связанные с гипоксией пути GO значительно активизировались в группе 48 ч, включая «путь PID HIF1 TF», «реакцию на гипоксию», «позитивную регуляцию клеточной гибели» и «внутренний апоптотический сигнальный путь». . Было показано, что инфекция SARS-CoV-2 запускает митохондриальную продукцию АФК, что индуцирует стабилизацию индуцируемого гипоксией фактора-1а (HIF-1a) в моноцитах. 16 Аналогичным образом в Т-клетках активировались несколько генов, участвующих в этом ответе на окислительный стресс: BNIP3, PFKFB3, FOS, JUN, BHLHE40, GADD45B, PDK1 и DDIT4 (рис. 4d). Чтобы подтвердить выводы о линиях Т-клеток, мы провели анализ секвенирования РНК первичных мононуклеарных клеток периферической крови (РВМС), взятых у трех здоровых доноров и трех пациентов с тяжелым течением COVID-19. Наши данные показали усиление клеточных ответов на стимулы, гибель клеток или реакцию на пути гипоксии, а также снижение активации лейкоцитов и сигнальных путей, аналогично результатам в линии Т-клеток (рис.
По сравнению с группой 24 ч, связанные с гипоксией пути GO значительно активизировались в группе 48 ч, включая «путь PID HIF1 TF», «реакцию на гипоксию», «позитивную регуляцию клеточной гибели» и «внутренний апоптотический сигнальный путь». . Было показано, что инфекция SARS-CoV-2 запускает митохондриальную продукцию АФК, что индуцирует стабилизацию индуцируемого гипоксией фактора-1а (HIF-1a) в моноцитах. 16 Аналогичным образом в Т-клетках активировались несколько генов, участвующих в этом ответе на окислительный стресс: BNIP3, PFKFB3, FOS, JUN, BHLHE40, GADD45B, PDK1 и DDIT4 (рис. 4d). Чтобы подтвердить выводы о линиях Т-клеток, мы провели анализ секвенирования РНК первичных мононуклеарных клеток периферической крови (РВМС), взятых у трех здоровых доноров и трех пациентов с тяжелым течением COVID-19. Наши данные показали усиление клеточных ответов на стимулы, гибель клеток или реакцию на пути гипоксии, а также снижение активации лейкоцитов и сигнальных путей, аналогично результатам в линии Т-клеток (рис. 4e). Таким образом, инфекция SARS-CoV-2 индуцировала выраженную гибель Т-клеток, которая, вероятно, зависит от путей митохондриальной АФК-гипоксии.
4e). Таким образом, инфекция SARS-CoV-2 индуцировала выраженную гибель Т-клеток, которая, вероятно, зависит от путей митохондриальной АФК-гипоксии.
Исследование потенциальных рецепторов в Т-клетках
Поскольку наши результаты показали, что заражение SARS-CoV-2 Т-клетками Jurkat не зависит от ACE2, мы попытались идентифицировать потенциальные рецепторы. Сначала мы исследовали экспрессию известных рецепторов или кофакторов SARS-CoV-2, которые были идентифицированы в первичных Т-клетках из общедоступных данных одноклеточного NGS 14 , а также в Т-клетках Jurkat в анализе секвенирования РНК с активацией или без нее. , включая ACE2/TMPRSS2, AXL, NRP1, KIM-1/TIM-1, ASGR1 и KREMEN1. 18,19,20 Кроме того, предполагалось, что ITGB2 (лейкоцит-ассоциированная молекула-1, LFA-1), молекула адгезии лейкоцитов связывается с ORF7a SARS-CoV-1. 21 Поскольку SARS-CoV-2 имеет ту же ORF7a, что и SARS-CoV-1, было бы интересно оценить, опосредует ли LFA-1 также инфицирование T-клеток SARS-CoV-2.
Наши данные показали минимальную экспрессию следующих молекул в SARS-CoV-2-положительных Т-клетках пациентов: ACE2, TMPRSS2, ASGR1, KREMEN1 и NRP1 (рис. 5a и дополнительная рис. S3a). Напротив, в этих клетках экспрессировались AXL и LFA-1. В клетках Jurkat LFA-1 также демонстрировал очень высокую экспрессию, хотя она не повышалась после 2-часовой активации (дополнительная рис. S3b). Взятые вместе, AXL и LFA-1 оказались многообещающими мишенями в качестве входных молекул.
Рис. 5 Исследование потенциальных рецепторов в Т-клетках. a Экспрессия ACE2, TMPRSS2 и ITGB2 (LFA-1) в Т-клетках крови здоровых доноров и пациентов с COVID-19. Анализ зависит от общедоступных данных NGS для одной ячейки. 14 Экспрессия трех генов показана в РНК-позитивных Т-клетках вируса SARS-CoV-2 для пациентов в уголовном порядке. b BEAS-2B и активированные клетки Jurkat инкубировали с различными концентрациями белков AXL (25, 50 или 100 мкг/мл) при 37 °C в течение 30 минут до заражения SARS-CoV-2 (MOI 0,01). Внутриклеточную вирусную РНК (RBD) через 24 часа после инфицирования определяли количественно с помощью количественной ПЦР. c Нокдаун или сверхэкспрессию AXL проводили на клетках Jurkat, а уровень экспрессии AXL определяли с помощью количественной ПЦР. Линии клеток инфицировали SARS-CoV-2 при множественности заражения 0,01 в течение 24 часов, после чего определяли количество вирусной РНК. d LFA-1 стабильно сверхэкспрессировался в клетках Caco2 с нокдауном ACE2 (Caco2-ACE2-shRNA) или клетках Jurkat, и уровень РНК определяли количественно с помощью количественной ПЦР. e Caco2, активированный Jurkat и их соответствующие клетки со сверхэкспрессией LFA-1 были инфицированы SARS-CoV-2 при MOI 0,01 и собраны через 24 часа после заражения. Внутриклеточную вирусную РНК выявляли с помощью количественной ПЦР. f , г Caco2, Caco2-ACE2-shRNA и его клеточная линия со сверхэкспрессией LFA-1 были инфицированы SARS-CoV-2 при MOI = 5 в течение 8 часов. Микроскоп высокого содержания использовали для наблюдения ( f ) и количественного определения ( g ) вирусных NP-положительных клеток.
Внутриклеточную вирусную РНК (RBD) через 24 часа после инфицирования определяли количественно с помощью количественной ПЦР. c Нокдаун или сверхэкспрессию AXL проводили на клетках Jurkat, а уровень экспрессии AXL определяли с помощью количественной ПЦР. Линии клеток инфицировали SARS-CoV-2 при множественности заражения 0,01 в течение 24 часов, после чего определяли количество вирусной РНК. d LFA-1 стабильно сверхэкспрессировался в клетках Caco2 с нокдауном ACE2 (Caco2-ACE2-shRNA) или клетках Jurkat, и уровень РНК определяли количественно с помощью количественной ПЦР. e Caco2, активированный Jurkat и их соответствующие клетки со сверхэкспрессией LFA-1 были инфицированы SARS-CoV-2 при MOI 0,01 и собраны через 24 часа после заражения. Внутриклеточную вирусную РНК выявляли с помощью количественной ПЦР. f , г Caco2, Caco2-ACE2-shRNA и его клеточная линия со сверхэкспрессией LFA-1 были инфицированы SARS-CoV-2 при MOI = 5 в течение 8 часов. Микроскоп высокого содержания использовали для наблюдения ( f ) и количественного определения ( g ) вирусных NP-положительных клеток. ч Нокдаун LFA-1 Клетки Jurkat инфицировали SARS-CoV-2 (MOI = 0,01) в течение 24 ч, а экспрессию LFA-1 и внутриклеточной вирусной РНК анализировали с помощью количественной ПЦР. i Активированные клетки Jurkat предварительно обрабатывали Lifitegrast, ингибитором LFA-1, в различных концентрациях (50, 100 или 200 нМ) при 37 °C в течение 30 мин до инфицирования (MOI = 0,01). Внутриклеточную вирусную РНК количественно определяли через 24 ч после инфицирования. Статистические данные были выполнены с использованием Стьюдента T Тест (* P <0,05; ** P <0,01; *** P <0,001; **** P <0,0001; NS NO) В предыдущем исследовании предполагалось, что AXL может быть рецептором-кандидатом для SARS-CoV-2, и его функция в опосредовании инфекции SARS-CoV-2 не зависит от ACE2. 19 BEAS-2B, использовавшийся в качестве положительного контроля в исследованиях AXL-SARS-CoV-2, был предварительно обработан белками AXL различных концентраций (25, 50, 100 мкг/мл) в течение 30 мин, а затем инфицирован SARS-CoV -2.
ч Нокдаун LFA-1 Клетки Jurkat инфицировали SARS-CoV-2 (MOI = 0,01) в течение 24 ч, а экспрессию LFA-1 и внутриклеточной вирусной РНК анализировали с помощью количественной ПЦР. i Активированные клетки Jurkat предварительно обрабатывали Lifitegrast, ингибитором LFA-1, в различных концентрациях (50, 100 или 200 нМ) при 37 °C в течение 30 мин до инфицирования (MOI = 0,01). Внутриклеточную вирусную РНК количественно определяли через 24 ч после инфицирования. Статистические данные были выполнены с использованием Стьюдента T Тест (* P <0,05; ** P <0,01; *** P <0,001; **** P <0,0001; NS NO) В предыдущем исследовании предполагалось, что AXL может быть рецептором-кандидатом для SARS-CoV-2, и его функция в опосредовании инфекции SARS-CoV-2 не зависит от ACE2. 19 BEAS-2B, использовавшийся в качестве положительного контроля в исследованиях AXL-SARS-CoV-2, был предварительно обработан белками AXL различных концентраций (25, 50, 100 мкг/мл) в течение 30 мин, а затем инфицирован SARS-CoV -2. Заражение SARS-CoV-2 может быть значительно ингибировано белком AXL в концентрации 25 мкг/мл. Напротив, заражение клеток Jurkat SARS-CoV-2 не удалось подавить даже при 100 мкг/мл (рис. 5b). Затем мы сконструировали клеточные линии с нокдауном или сверхэкспрессией AXL на клетках Jurkat, а затем проверили влияние на вирусную инфекцию. Наши данные показали, что нокдаун AXL не может блокировать инфекцию SARS-CoV-2, но сверхэкспрессия AXL может немного усиливать инфекцию (в 1,5 раза) (рис. 5c). В совокупности AXL не должен быть основным рецептором для SARS-CoV-2 в клетках Jurkat, но может способствовать инфицированию.
Заражение SARS-CoV-2 может быть значительно ингибировано белком AXL в концентрации 25 мкг/мл. Напротив, заражение клеток Jurkat SARS-CoV-2 не удалось подавить даже при 100 мкг/мл (рис. 5b). Затем мы сконструировали клеточные линии с нокдауном или сверхэкспрессией AXL на клетках Jurkat, а затем проверили влияние на вирусную инфекцию. Наши данные показали, что нокдаун AXL не может блокировать инфекцию SARS-CoV-2, но сверхэкспрессия AXL может немного усиливать инфекцию (в 1,5 раза) (рис. 5c). В совокупности AXL не должен быть основным рецептором для SARS-CoV-2 в клетках Jurkat, но может способствовать инфицированию.
LFA-1 широко экспрессируется на поверхности многих лейкоцитов, и активация Т-клеток изменила структуру LFA-1 на высокоаффинный режим, но не уровень экспрессии. 22 Затем мы сверхэкспрессировали высокоаффинную альфа-субъединицу белка LFA-1 в клетках Caco2 с нокдауном ACE2 (Caco2-ACE2-shRNA) и клетках Jurkat. Наши данные количественной ПЦР показали, что сверхэкспрессия LFA-1 успешно восстановила подавленную инфекцию в клетках Caco2 с нокдауном ACE2, а также усилила вирусную инфекцию в клетках Jurkat (увеличение в три раза), как показано на уровнях клеточной вирусной РНК (рис. 5d, e). Чтобы подтвердить вывод, мы также провели IFA для обнаружения экспрессии вирусных NP. После 8-часовой инфекции сравнивали вирусные NP-положительные клетки. Наши данные показали ослабленную инфекцию SARS-CoV-2 в клетках с нокдауном ACE2 и гораздо более высокий NP в клетках с избыточной экспрессией LFA-1 ACE2-нокдаун (рис. 5f, g).
5d, e). Чтобы подтвердить вывод, мы также провели IFA для обнаружения экспрессии вирусных NP. После 8-часовой инфекции сравнивали вирусные NP-положительные клетки. Наши данные показали ослабленную инфекцию SARS-CoV-2 в клетках с нокдауном ACE2 и гораздо более высокий NP в клетках с избыточной экспрессией LFA-1 ACE2-нокдаун (рис. 5f, g).
Наконец, клеточная линия Jurkat с нокдауном LFA-1 была сконструирована и инфицирована SARS-CoV-2 (MOI = 0,01). Через 24 часа после инфицирования вирусная нагрузка в нокдаунной клеточной линии была значительно снижена по сравнению с контрольной клеточной линией (рис. 5h). Лифитеграст, ингибитор, блокирующий связывание LFA-1 с его внеклеточным лигандом, также использовали для предварительной обработки активированных клеток Jurkat перед инфекцией. Результаты количественной ПЦР показали, что при концентрации 200 нМ лифитеграст также может снижать вирусную нагрузку в клетках Jurkat (рис. 5i). В совокупности наши результаты показали, что LFA-1 должен быть фактором прикрепления или потенциальным молекулярным входом для SARS-CoV-2 во время его заражения в клетках Jurkat.
Обсуждение
Здесь мы показали, что SARS-CoV-2 инфицировал Т-лимфоциты, в основном CD4 + Т-клетки, независимым от АПФ2 образом. Инфекция SARS-CoV-2 вызывала выраженную гибель Т-клеток, что потенциально способствовало лимфопении у пациентов с COVID-19. Инфекция Т-клеток также может оказывать сильное влияние на пациентов. Инфицированные Т-лимфоциты не только утратили способность контролировать вирусную инфекцию, но также могут переносить вирусы в другие части тела посредством кровообращения. Кроме того, этот независимый от ACE2 способ заражения может поставить под угрозу терапевтический эффект нейтрализующих антител, нацеленных на связывание шипа с ACE2. Это может синергетически привести к более тяжелым исходам инфекции у пациентов с COVID-19..
Обсуждается, нарушил ли SARS-CoV-2 функциональность популяций иммунных клеток в результате прямой инфекции. Наши результаты предоставили доказательства того, что инфицированные SARS-CoV-2 Т-клетки, такие как вирусная РНК, вирусная sgРНК, вирусный белок и инфекционный вирус, могут быть обнаружены в Т-клетках при инфицировании или в ПБЦ пациентов, хотя продукция инфекционного вируса частицы могут оставаться на низком уровне. Несколько недавних исследований также показали, что множественные иммунные клетки несут вирусный антиген или вирусную РНК, включая нейтрофилы, макрофаги, воспалительные моноциты, В-клетки плазмы, Т-клетки и NK-клетки, посредством посмертного гистологического анализа и секвенирования одноклеточной/одноядерной РНК. легкие или БАЛ. 13,14,15 Это говорит о том, что SARS-CoV-2 должен иметь широкий тропизм клеток-мишеней, включая основные популяции иммунных клеток.
Несколько недавних исследований также показали, что множественные иммунные клетки несут вирусный антиген или вирусную РНК, включая нейтрофилы, макрофаги, воспалительные моноциты, В-клетки плазмы, Т-клетки и NK-клетки, посредством посмертного гистологического анализа и секвенирования одноклеточной/одноядерной РНК. легкие или БАЛ. 13,14,15 Это говорит о том, что SARS-CoV-2 должен иметь широкий тропизм клеток-мишеней, включая основные популяции иммунных клеток.
Белки ACE2 и TMPRSS2 человека были признаны основными белками, опосредующими проникновение в клетки SARS-CoV-2. 14 Недавно обнаруженные связывающие молекулы AXL и NRP1 все еще зависят от ACE2 как основного рецептора. 18,19 Наше открытие ACE2-независимой инфекции Т-клеток неожиданно, но оно также подтверждается предыдущими открытиями о том, что существует РНК SARS-CoV-2 + клеток, которые не экспрессировали совместно ACE2 и TMPRSS2. 15 По нашим данным, SARS-CoV-2 показал значительную инфекцию активированных Т-клеток, что позволяет предположить наличие нового механизма проникновения в Т-клетки. Идентификация LFA-1 как входной молекулы, которая способствовала заражению Т-клеток SARS-CoV-2, будет иметь важное значение для разработки клинических терапевтических средств, хотя вопросы в будущем остаются. Например, что такое связывающий белок LFA-1 в вирионе SARS-CoV-2, если это не спайковый белок. Поскольку LFA-1 экспрессируется в ряде других лейкоцитов, можно ожидать, что другие иммунные клетки (включая макрофаги или моноциты) также могут быть инфицированы SARS-CoV-2 потенциально через связывание с LFA-1. Эти вопросы должны быть рассмотрены в будущих исследованиях.
Идентификация LFA-1 как входной молекулы, которая способствовала заражению Т-клеток SARS-CoV-2, будет иметь важное значение для разработки клинических терапевтических средств, хотя вопросы в будущем остаются. Например, что такое связывающий белок LFA-1 в вирионе SARS-CoV-2, если это не спайковый белок. Поскольку LFA-1 экспрессируется в ряде других лейкоцитов, можно ожидать, что другие иммунные клетки (включая макрофаги или моноциты) также могут быть инфицированы SARS-CoV-2 потенциально через связывание с LFA-1. Эти вопросы должны быть рассмотрены в будущих исследованиях.
Инфицирование CD4 + Т-лимфоцитов вирусом SARS-CoV-2 может быть основным фактором патогенеза, индуцированного вирусом. Вооруженные Т-клетки играют ключевую роль в борьбе с патогенной инфекцией. 10 Как показывают наши данные, эти Т-клетки, вероятно, являются мишенями инфекции SARS-CoV-2 и подвергаются апоптозу в HIF-1a-зависимом пути. Эти события могут привести к дисфункции Т-клеток, их истощению и, в конечном итоге, к лимфопении у пациентов. Кроме того, погибающие CD4 + Т-лимфоциты могут спровоцировать чрезмерное воспаление, которое приводит к тяжелому иммунопатогенезу у пациентов. Примечательно, что популяция CD8 + Т-лимфоцитов также значительно снижается при COVID-19.пациенты. В отличие от CD4 + Т-лимфоцитов, в проточной цитометрии не было определено, что эти клетки содержат вирусный антиген SARS-CoV-2. Механизм, лежащий в основе истощения CD8 + T-лимфоцитов, вызванного инфекцией SARS-CoV-2, в настоящее время неизвестен. Помимо вирусной инфекции, истощению лимфоцитов также могут способствовать несколько механизмов, включая присутствие эндогенных или экзогенных глюкокортикоидов, сверхактивированных ингибиторов высвобождения нейтрофилов активации Т-клеток (аргиназы 1 и CD274) и регулируемой цитокинами избирательной дифференцировки клеток костного мозга. 11,23 Необходимы дальнейшие углубленные исследования для выявления потенциально мультирежимных механизмов, которые приводят к лимфопении у пациентов с COVID-19.
Кроме того, погибающие CD4 + Т-лимфоциты могут спровоцировать чрезмерное воспаление, которое приводит к тяжелому иммунопатогенезу у пациентов. Примечательно, что популяция CD8 + Т-лимфоцитов также значительно снижается при COVID-19.пациенты. В отличие от CD4 + Т-лимфоцитов, в проточной цитометрии не было определено, что эти клетки содержат вирусный антиген SARS-CoV-2. Механизм, лежащий в основе истощения CD8 + T-лимфоцитов, вызванного инфекцией SARS-CoV-2, в настоящее время неизвестен. Помимо вирусной инфекции, истощению лимфоцитов также могут способствовать несколько механизмов, включая присутствие эндогенных или экзогенных глюкокортикоидов, сверхактивированных ингибиторов высвобождения нейтрофилов активации Т-клеток (аргиназы 1 и CD274) и регулируемой цитокинами избирательной дифференцировки клеток костного мозга. 11,23 Необходимы дальнейшие углубленные исследования для выявления потенциально мультирежимных механизмов, которые приводят к лимфопении у пациентов с COVID-19. Принимая во внимание очевидную корреляцию между лимфопенией и прогрессированием заболевания у пациентов с COVID-19, важно разработать стратегии профилактики вирус-индуцированной лимфопении.
Принимая во внимание очевидную корреляцию между лимфопенией и прогрессированием заболевания у пациентов с COVID-19, важно разработать стратегии профилактики вирус-индуцированной лимфопении.
Материалы и методы
Образцы и этика
Образцы крови и тканей человека от пациентов с COVID-19 или от здоровых доноров были взяты больницей Тунцзи с согласия всех лиц. Были приготовлены свежие срезы биопсии легкого от умершего пациента. Комитет по этике назначенных больниц для новых инфекционных заболеваний одобрил все образцы, взятые у людей.
Клеточные линии и вирусная культура
Vero E6, Caco2, 293T-sg, GP2-293 и BEAS-2B в DMEM + 10% FBS или Т-клетки MT4 и Jurkat в RPMI1640 + 10% FBS (Gibco, C22400500BT) , или клетки A549 в среде DMEM/F12 + 10% FBS, или первичные Т-клетки в среде X-vivo (Lonza, 04-418Q), содержащей IL-2 (Peprotech, 200-02), культивировали при 37°C во влажной атмосфере. 5% CO 2 . Все клеточные линии были протестированы на отсутствие контаминации микоплазмой и применены для идентификации видов и аутентификации с помощью микроскопической морфологической оценки. Ни одна из клеточных линий не была включена в список часто ошибочно идентифицируемых клеточных линий (согласно ICLAC). В этом исследовании использовали изолят SARS-CoV-2 WIV04 (инвентарный номер GISAID EPI_ISL_402124). WIV04 выделяли из клеток Huh7 исходного образца и пассировали в клетках Caco2. Титр вируса (TCID50/мл) определяли в клетках Vero E6.
Ни одна из клеточных линий не была включена в список часто ошибочно идентифицируемых клеточных линий (согласно ICLAC). В этом исследовании использовали изолят SARS-CoV-2 WIV04 (инвентарный номер GISAID EPI_ISL_402124). WIV04 выделяли из клеток Huh7 исходного образца и пассировали в клетках Caco2. Титр вируса (TCID50/мл) определяли в клетках Vero E6.
Белки и антитела к SARS-CoV-2
NP штамма SARS-CoV-2 WIV04 и предсказанный RBD были вставлены в вектор pCAGGS с N-концевой S-меткой. Сконструированные плазмиды временно трансфицировали в HEK293T-17. Супернатант, собранный для очистки белка, очищали с помощью смолы S-метки, чистоту и выход тестировали с использованием mAb против S-метки (собственного производства). Кроликов иммунизировали очищенными белками NP три раза в дозе 700 нг/каждый с интервалом в 2 недели. Сыворотку кроликов собирали через 10 дней после последней иммунизации. Титр антител определяли с помощью ELISA с использованием очищенного белка NP в качестве антигена для обнаружения.
Подготовка клеток периферической крови (ПБК) и инфекция SARS-CoV-2
Образцы крови пациентов с COVID-19 или здоровых доноров были обработаны в лаборатории BSL3 в WIV. Всего перед экспериментом из 10-кратного буфера для лизиса RBC eBioscience™ был приготовлен 1× буфер для лизиса RBC (Multi-species, Invitrogen). Образцы крови человека центрифугировали при 500 × g в течение 10 минут, а затем обрабатывали 2 мл 1× буфера для лизиса эритроцитов в течение не более 15 минут при комнатной температуре. Клетки центрифугировали при 500 × г в течение 10 минут с последующей обработкой с использованием 2 мл 1× буфера для лизиса эритроцитов в течение еще 10 минут при комнатной температуре для удаления остатков эритроцитов. Клетки были готовы к использованию после центрифугирования. Клетки отмывали центрифугированием (500 × г в течение 10 минут каждый раз) три раза с PBS, содержащим 2% BSA, перед окрашиванием антител к клеточным маркерам.
Для инфицирования PBC высевали в 24-луночные планшеты в культуральной среде Roswell Park Memorial Institute 1640 (RPMI1640, ThermoFisher, 22400500BT) с добавлением 10% эмбриональной бычьей сыворотки (FBS, Life Technologies, 1009). 9141) при плотности 1 × 10 6 клеток/мл. ПБЦ были инфицированы SARS-CoV-2 при 0,1 множественности множественности. Через час после инкубации клетки трижды отмывали центрифугированием с использованием RPMI1640. Затем PBC засевали RPMI1640 с добавлением 10% FBS в новые 24-луночные планшеты при 37°C, снабженные 5% CO 2 , в течение 12 или 24 часов перед сбором для дальнейшего анализа.
9141) при плотности 1 × 10 6 клеток/мл. ПБЦ были инфицированы SARS-CoV-2 при 0,1 множественности множественности. Через час после инкубации клетки трижды отмывали центрифугированием с использованием RPMI1640. Затем PBC засевали RPMI1640 с добавлением 10% FBS в новые 24-луночные планшеты при 37°C, снабженные 5% CO 2 , в течение 12 или 24 часов перед сбором для дальнейшего анализа.
Для ИФА на ПБЦ пациентов фиксированные в течение ночи клетки равномерно наносили на покровное стекло. Присутствие вирусных NP определяли с помощью кроличьего pAb против белка NP SARS-CoV-2 (собственно созданного, 1:1000) и козьего антикроличьего IgG, конъюгированного с Cy3 (1:200, Abcam, ab69).39). Т-лимфоциты выявляли с использованием кроличьего антитела против CD3 человека (1:100, Abcam, ab5690). Ядра окрашивали DAPI (Beyotime, C1002). Картины окрашивания изучали с помощью конфокальной микроскопии на микроскопе FV1200 (Olympus).
Для иммуногистохимического анализа легкого пациента биопсийные ткани умершего пациента фиксировали 4% параформальдегидом в течение 24 часов, заливали парафином и разрезали на срезы толщиной 5 мкм. Мультиплексное иммунофлуоресцентное окрашивание получали с использованием набора PANO 7-plex IHC (0004100100, Panovue, Пекин, Китай). Предметные стекла депарафинизировали и регидратировали с последующим 15-минутным выделением антигена, индуцированного нагреванием, с ЭДТА, рН 9..0. Предметные стекла промывали PBS/0,02% Triton X-100, затем блокировали 10% BSA при комнатной температуре в течение 30 мин, кроличьим pAb против белка NP SARS-CoV-2 (создано собственными силами, 1:1000) и кроличьим анти- Человеческое антитело к CD3 (1:100, Abcam, ab5690) затем использовали в инкубации при 37°C в течение 1 ч с последующей инкубацией вторичного антитела, конъюгированного с пероксидазой хрена, и усилением тирамидного сигнала. Предметные стекла подвергали микроволновой термообработке после каждой операции TSA. На заключительном этапе окрашивания ядра окрашивали 4’-6’-диамидино-2-фенилиндолом (DAPI, Beyotime, C1002). Для получения мультиспектральных изображений окрашенные предметные стекла сканировали с помощью системы Mantra System (PerkinElmer).
Мультиплексное иммунофлуоресцентное окрашивание получали с использованием набора PANO 7-plex IHC (0004100100, Panovue, Пекин, Китай). Предметные стекла депарафинизировали и регидратировали с последующим 15-минутным выделением антигена, индуцированного нагреванием, с ЭДТА, рН 9..0. Предметные стекла промывали PBS/0,02% Triton X-100, затем блокировали 10% BSA при комнатной температуре в течение 30 мин, кроличьим pAb против белка NP SARS-CoV-2 (создано собственными силами, 1:1000) и кроличьим анти- Человеческое антитело к CD3 (1:100, Abcam, ab5690) затем использовали в инкубации при 37°C в течение 1 ч с последующей инкубацией вторичного антитела, конъюгированного с пероксидазой хрена, и усилением тирамидного сигнала. Предметные стекла подвергали микроволновой термообработке после каждой операции TSA. На заключительном этапе окрашивания ядра окрашивали 4’-6’-диамидино-2-фенилиндолом (DAPI, Beyotime, C1002). Для получения мультиспектральных изображений окрашенные предметные стекла сканировали с помощью системы Mantra System (PerkinElmer). Сканы были объединены для создания единого изображения стека. Неокрашенные изображения и одноокрашенные срезы использовали для извлечения спектра автофлуоресценции тканей или каждого флуоресцеина соответственно. Извлеченные изображения далее использовались для создания спектральной библиотеки, необходимой для мультиспектрального смешения с помощью программного обеспечения для анализа изображений InForm (PerkinElmer). Используя эту спектральную библиотеку, мы получили реконструированные изображения срезов с удаленной автофлуоресценцией.
Сканы были объединены для создания единого изображения стека. Неокрашенные изображения и одноокрашенные срезы использовали для извлечения спектра автофлуоресценции тканей или каждого флуоресцеина соответственно. Извлеченные изображения далее использовались для создания спектральной библиотеки, необходимой для мультиспектрального смешения с помощью программного обеспечения для анализа изображений InForm (PerkinElmer). Используя эту спектральную библиотеку, мы получили реконструированные изображения срезов с удаленной автофлуоресценцией.
ПБЦ, совместно культивированные с клетками Caco2
ПБЦ пяти пациентов трижды промывали PBS перед совместным культивированием с клетками Caco2 в течение 4 дней и тестировали на вирусную РНК SARS-CoV-2 в супернатанте или антиген в клетках Caco2. Тем временем PBC анализировали на присутствие вирусного антигена с помощью проточной цитометрии.
Культура органоидов толстой кишки человека и инфекция SARS-CoV-2
Органоиды толстой кишки человека были получены и культивированы, как описано в предыдущем исследовании. 24 Вкратце, органоиды толстой кишки в матригеле расщепляли и дважды промывали средой перед заражением. SARS-CoV-2 был добавлен для заражения органоидов толстой кишки при MOI 0,01. Через 24 ч органоиды толстой кишки центрифугировали и дважды промывали средой. Вирусную РНК в органоидах толстой кишки определяли с помощью количественной ПЦР.
24 Вкратце, органоиды толстой кишки в матригеле расщепляли и дважды промывали средой перед заражением. SARS-CoV-2 был добавлен для заражения органоидов толстой кишки при MOI 0,01. Через 24 ч органоиды толстой кишки центрифугировали и дважды промывали средой. Вирусную РНК в органоидах толстой кишки определяли с помощью количественной ПЦР.
Активация Jurkat и первичных Т-клеток
Для активации клеток Jurkat 2,5E + 06 клеток высевали в лунку шестилуночного планшета, содержащую 2,5 мл среды RPMI1640, содержащей 10% FBS. Всего к клеткам добавляли 40 нг/мл ФМА (Invivogen, tlrl-pma) и инкубировали при 37°С в течение 2 ч. Клетки центрифугировали при 300 × г при комнатной температуре в течение 10 минут перед удалением супернатанта и культивированием со свежей средой RPMI1640, содержащей 10% FBS. Первичные CD3 T-лимфоциты человека выделяли из крови здоровых доноров с использованием CD3 Microbeads of Human (Miltenyi, 130-050-101). Для активации первичных Т-клеток замороженные Т-клетки размораживали и культивировали с X-vivo (Lonza, 04-418Q), содержащим 1 мкг/мл IL-2 (Peprotech, 200-02). Клетки культивировали в объеме 7,5 мкл T Cell TransAct (Miltenyi Biotec) в среде в течение 3 сут при 37 °С. Затем клетки центрифугировали и культивировали со свежей средой IL-2/X-vivo до заражения вирусом.
Клетки культивировали в объеме 7,5 мкл T Cell TransAct (Miltenyi Biotec) в среде в течение 3 сут при 37 °С. Затем клетки центрифугировали и культивировали со свежей средой IL-2/X-vivo до заражения вирусом.
Инфекция Т-клеток
Т-клетки Jurkat или первичные Т-лимфоциты CD3 человека были инфицированы SARS-CoV-2 при множественности заражения 0,01, 0,1 или 1 в зависимости от цели эксперимента. Супернатант или клетки собирали при 0, 24, 48 или 72 hpi после трехкратной промывки PBS для Т-клеток Jurkat или при 0, 4, 8 и 12 hpi для первичных Т-клеток. Экспрессию клеточной или супернатантной вирусной РНК или белка определяли с помощью количественной ПЦР, секвенирования РНК, WB или проточной цитометрии. GAPDH использовали в количественной ПЦР в качестве внутреннего контроля, а бета-тубулин использовали в WB (1:5000, 66240-1-Ig от Proteintech) в качестве внутреннего контроля.
Анализ образцов периферической крови человека методом проточной цитометрии
Для поверхностного окрашивания КПК перед фиксацией инкубировали со специфическими для человека антителами, мечеными флуорохромом: AF-700-анти-CD45 (2D1), percp-анти-CD19 (HIB19), APC/CY7-анти-CD3 (UCHT1), BV510-анти-CD4 (OKT4) и percp/Cy5. 5-анти-CD8a (HTT8a). Окрашенные антителами PBC фиксировали в течение ночи с помощью 4% PFA при 4 °C и вывозили из лаборатории BSL3 для последующего анализа. Клетки дополнительно окрашивали NP pAb SARS-CoV-1 собственного производства (1:500) при 4°C в течение 30 минут после пермеабилизации. Затем клетки окрашивали FITC-anti-Rabbit IgG (H + L) при комнатной температуре в течение 30 мин. Антитела AF-700-анти-CD45 (2D1), APC/CY7-анти-CD3 (UCHT1), BV510-анти-CD4 и percp/Cy5.5-анти-CD8a были приобретены у Biolegend, и все они были использованы в 1: 100. FITC-антикроличьи IgG (H + L) были получены от Proteintech (SA00003-2).
5-анти-CD8a (HTT8a). Окрашенные антителами PBC фиксировали в течение ночи с помощью 4% PFA при 4 °C и вывозили из лаборатории BSL3 для последующего анализа. Клетки дополнительно окрашивали NP pAb SARS-CoV-1 собственного производства (1:500) при 4°C в течение 30 минут после пермеабилизации. Затем клетки окрашивали FITC-anti-Rabbit IgG (H + L) при комнатной температуре в течение 30 мин. Антитела AF-700-анти-CD45 (2D1), APC/CY7-анти-CD3 (UCHT1), BV510-анти-CD4 и percp/Cy5.5-анти-CD8a были приобретены у Biolegend, и все они были использованы в 1: 100. FITC-антикроличьи IgG (H + L) были получены от Proteintech (SA00003-2).
Экстракция РНК и количественная ПЦР
Всякий раз, когда использовались коммерческие наборы, инструкции производителя следовали без изменений. РНК экстрагировали из 140 мкл образцов с помощью набора QIAamp ® Viral RNA Mini Kit (QIAGEN). РНК элюировали 50 мкл элюирующего буфера и использовали в качестве матрицы. Обнаружение SARS-CoV-2 методом кПЦР было выполнено с использованием набора HiScript ® II для одноэтапной кПЦР SYBR ® Green Kit плюс набора зондов для одноэтапной кПЦР, направленного либо на M для sgRNA (разработано собственными силами), либо на RBD гена шипа. (коммерческий) в соответствии с инструкциями производителя (Q222-CN, Vazyme Biotech Co., Ltd). QPCR запускали на машине Step-One Plus для ПЦР в реальном времени (ABI) с настройками по умолчанию.
(коммерческий) в соответствии с инструкциями производителя (Q222-CN, Vazyme Biotech Co., Ltd). QPCR запускали на машине Step-One Plus для ПЦР в реальном времени (ABI) с настройками по умолчанию.
Анализ глубины и покрытия генома SARS-CoV-2
РНК была выделена из 24-часовых активированных T-клеток Jurkat, инфицированных SARS-CoV-2, с помощью набора RNAprep Pure Cell/Bacteria Kit (TIANGEN, DP430). РНК элюировали 50 мкл элюирующего буфера и использовали в качестве матрицы для РНК-секвенирования. Чистые считывания были сопоставлены с геномом SARS-CoV-2 (WIV04) с использованием программного обеспечения HISAT2 v2.1.0. После сортировки и индексации с помощью samtools v1.10-24 покрытие было рассчитано с использованием функции genomeCoverageBed из bedtools v2.29.2.
Транскриптомный анализ
T-клетки Jurkat, инфицированные SARS-CoV-2 через 24 часа и 48 часов (по 3 повторения каждая), образцы крови трех здоровых доноров и 3 пациентов с тяжелой формой COVID-19 были подвергнуты анализу РНК-seq. После сопоставления чистых прочтений с GRCh48.p13 с помощью HISAT2 v2.1.0 и преобразования формата с помощью samtools v1.10-24 мы использовали stringtie v2.1.0 для сборки и количественного определения транскриптов. Таблица количества прочтений транскриптома, сгенерированная prepDE.py, инструментом в stringtie, использовалась для анализа дифференциальной экспрессии генов в R v4.1.0 с пакетом DESeq2 v1.32.0. Ген с log2-кратным изменением >2 и Значение P <0,05 было выбрано для выполнения анализа обогащения с использованием онлайн-инструментов Metascape.
После сопоставления чистых прочтений с GRCh48.p13 с помощью HISAT2 v2.1.0 и преобразования формата с помощью samtools v1.10-24 мы использовали stringtie v2.1.0 для сборки и количественного определения транскриптов. Таблица количества прочтений транскриптома, сгенерированная prepDE.py, инструментом в stringtie, использовалась для анализа дифференциальной экспрессии генов в R v4.1.0 с пакетом DESeq2 v1.32.0. Ген с log2-кратным изменением >2 и Значение P <0,05 было выбрано для выполнения анализа обогащения с использованием онлайн-инструментов Metascape.
Общедоступный анализ данных одноклеточного NGS
Публичные данные одноклеточного NGS были загружены, данные пациентов с COVID-19 были загружены из GSE158055 14 , а данные здоровых доноров были загружены из GSE134355 (ландшафт клеток человека). Согласно исходной информации каждой статьи, мы извлекли данные первичных Т-клеток из легких, тимуса и периферической крови здоровых доноров и вирус-положительных Т-клеток COVID-19. пациенты. Следуя стандартному рабочему процессу Seurat v4.0.4, мы нормализовали данные и масштабировали их с помощью информации UMI. Экспрессию рецепторов-кандидатов или кофакторов визуализировали с помощью функции Seurat FeaturePlot.
пациенты. Следуя стандартному рабочему процессу Seurat v4.0.4, мы нормализовали данные и масштабировали их с помощью информации UMI. Экспрессию рецепторов-кандидатов или кофакторов визуализировали с помощью функции Seurat FeaturePlot.
Вестерн-блоттинг (WB) анализ
Инфицированные или трансдуцированные клетки собирали в указанный момент времени и лизировали лизисным буфером RIPA (Beyotime, P0013C) для WB. Затем белки в клеточных лизатах разделяли с помощью электрофореза в 10–12% SDS-полиакриламидном геле (PAGE) и далее переносили на мембраны из поливинилидендифторида (PVDF) (Millipore, SLHVR33RB). Блоты инкубировали с кроличьим поликлональным анти-ACE2 (Servicebio, GB11267, разведение 1:1000), кроличьим поликлональным анти-2019-nCoV NP (разведение 1:1000), мышиные моноклональные антитела к бета-тубулину (Proteintech, 66240-1-Ig, разведение 1:5000), а затем соответствующие вторичные антитела кролика или мыши, конъюгированные с пероксидазой (Proteintech, разведение 1:5000). , SA00001-2 или SA00001-1). Для обнаружения белка использовали хемилюминесцентный субстрат HRP Immobilon western (Millipore, WBKLS0500).
, SA00001-2 или SA00001-1). Для обнаружения белка использовали хемилюминесцентный субстрат HRP Immobilon western (Millipore, WBKLS0500).
Анализ концевого мечения концевой дезоксинуклеотидилтрансферазы dUTP (TUNEL)
Набор для анализа TUNEL, приобретенный у Beyotime Biotechnology (C1088), использовался для обнаружения апоптоза в клетках, инфицированных SARS-CoV-2, в соответствии с инструкциями производителя. Вкратце, клетки, фиксированные в 4% параформальдегиде, пермеабилизировали 0,25% тритоном X-100 в течение 20 минут при 4°C. Затем к клеткам добавляли реакционную смесь TdT, содержащую фермент TdT и раствор флуоресцентной метки, для мечения фрагментированной ДНК. Клетки дополнительно окрашивали Rp3-CoV NP pAb (1:8000) или кроличьи анти-SARS-CoV-2 NP pAb (1:500) и CY3-антикроличьи IgG (H + L) (Proteintech, SA00009-2) после фиксации. Меченые клетки анализировали с помощью проточного цитометра (BD LSRFortessa).
Эксперименты по ингибированию конкуренции ACE2 и блокированию антител
Человеческий рекомбинантный полноразмерный белок ACE2-Fc (GenScript, Z03484), анти-ACE2 Ab (R&D, AF933) и RD#4-анти-Spike Ab (моноклональные антитела собственного производства ) были использованы. Белок ACE2-Fc разбавляли до 20 мкг/мкл в культуральной среде, а затем инкубировали с раствором вируса SARS-CoV-2 (MOI = 0,01) в объеме 1:1 при 37 °C в течение 30 мин. RD#4-анти-Spike Ab разбавляли до 320 нг/мкл в культуральной среде, а затем инкубировали с раствором вируса SARS-CoV-2 (MOI = 0,01) в объеме 1:1 при 37 °C в течение 30 мин. Затем к клеткам Jurkat или клеткам Caco2 добавляли смесь вирус-ACE2 или вирус-антитело. Клетки собирали для дальнейшего анализа через 24 ч после инфицирования. Для экспериментов по блокированию антител против ACE2 клетки Jurkat или клетки Caco2 предварительно обрабатывали 3,33 нг/мкл антитела против ACE2 (R&D Systems, козы, AF9).33) при 37 °С за 30 мин до заражения.
Белок ACE2-Fc разбавляли до 20 мкг/мкл в культуральной среде, а затем инкубировали с раствором вируса SARS-CoV-2 (MOI = 0,01) в объеме 1:1 при 37 °C в течение 30 мин. RD#4-анти-Spike Ab разбавляли до 320 нг/мкл в культуральной среде, а затем инкубировали с раствором вируса SARS-CoV-2 (MOI = 0,01) в объеме 1:1 при 37 °C в течение 30 мин. Затем к клеткам Jurkat или клеткам Caco2 добавляли смесь вирус-ACE2 или вирус-антитело. Клетки собирали для дальнейшего анализа через 24 ч после инфицирования. Для экспериментов по блокированию антител против ACE2 клетки Jurkat или клетки Caco2 предварительно обрабатывали 3,33 нг/мкл антитела против ACE2 (R&D Systems, козы, AF9).33) при 37 °С за 30 мин до заражения.
Генерация KO, KD, клеточных линий со сверхэкспрессией
KO, KD и плазмиды со сверхэкспрессией были сконструированы на разных векторах (pLenti-V2 для нокаута, вектор pLKO.1 для нокдауна и вектор pQCXIH для сверхэкспрессии). Нокаут ACE2 осуществляли путем трансдукции клеток Caco2 и Jurkat лентивирусами, экспрессирующими специфические sgRNA, нацеленные на ACE2 (F: CACCG GCCTCCATCGATATTAGCAA; R: AAAC TTGCTAATATCGATGGAGGCC).
Нокдаун ACE2, AXL, LFA-1 осуществляли путем трансдукции клеток Caco2 или Jurkat лентивирусами, экспрессирующими специфические siРНК (ACE2: 5′-GCCGAAGACCTGTTCTATCAA-3′; AXL: 5′-CCTGTGGTCATCTTACCTT-3′; LFA-1: 5′-GCCATCAATTATGTCGCGACA-3′ или киРНК скремблирования).
Затем трансдуцированные клетки культивировали с пуромицином (5 мкг/мл для Caco2 или 1,5 мкг/мл для Jurkat) в течение 7 дней.
Для сверхэкспрессии полную длину AXL или домен I альфа-субъединицы LFA-1 амплифицировали из клеток Hep G2 или клеток Jurkat соответственно. Клетки, трансдуцированные лентивирусом, культивировали с гигромицином (35 мкг/мл для клеток Caco2 и Jurkat) в течение 7 дней. Для заражения к клеткам добавляли вирус до конца эксперимента с 0,01 MOI. Зараженные клетки собирали при 24 HPI после двукратной промывки PBS. Экспрессию внутриклеточного вирусного белка определяли вестерн-блоттингом с антителом против вирусного белка NP, а вирусную РНК в цитоплазме определяли методом количественной ПЦР.
Анализ блокирования TMPRSS2
Мезилат камостата (MCE, HY-13512-10 мМ) разбавляли до конечной концентрации 20 мкМ или 2 мкМ. Всего к клеткам добавляли 100 мкл (для 48-луночного планшета) или 200 мкл (для 24-луночного планшета) растворов камостата. Через час активированные клетки Jurkat и Caco2 инфицировали SARS-CoV-2 при 0,01 MOI. Клеточный лизат собирали при 24 HPI и определяли вирусную РНК в цитоплазме с помощью количественной ПЦР. Вирусный NP анализировали вестерн-блоттингом.
Эксперименты по ингибированию конкуренции белков-кандидатов рецепторов
Рекомбинантный белок AXL человека (MedChemExpress, HY-P7622) разбавляли культуральной средой до различных концентраций, а затем инкубировали с вирусом SARS-CoV-2 (MOI = 0,01) в объеме 1:1 при 37 °C в течение 30 мин. . Затем добавляли смеси для заражения активированных клеток Jurkat и клеток BEAS-2B. Образцы собирали при 24 HPI и определяли клеточную вирусную РНК с помощью количественной ПЦР.
Эксперимент по ингибированию LFA-1
Лифитеграст (MedChemExpress, HY-19344) разводили до различных концентраций и предварительно обрабатывали активированные клетки Jurkat при 37 °C перед заражением. Тридцать минут спустя клетки были инфицированы SARS-CoV-2 (MOI = 0,01) и образцы были собраны при 24 HPI. Вирусную РНК в цитоплазме определяли методом количественной ПЦР.
Тридцать минут спустя клетки были инфицированы SARS-CoV-2 (MOI = 0,01) и образцы были собраны при 24 HPI. Вирусную РНК в цитоплазме определяли методом количественной ПЦР.
Электронная микроскопия
Активированные клетки Jurkat и MT4 были инфицированы SARS-CoV-2 (MOI = 1) в течение 72 часов. Клетки собирали и фиксировали 2,5% (масса/объем) глутарового альдегида и 1% четырехокиси осмия, обезвоживали с помощью ряда концентраций этанола (от 30 до 100%) и заливали эпоксидной смолой. Были приготовлены ультратонкие срезы (80 нм) встроенных клеток, нанесены на медные сетки с формваровым покрытием (200 меш), окрашены уранилацетатом и цитратом свинца и проанализированы с использованием электронного микроскопа Tecnai G2 200 кВ.
Статистический анализ
Анализ данных проводили с использованием программного обеспечения GraphPad Prism 7.0. Данные были представлены как среднее значение ± стандартное отклонение. Данные были проанализированы с помощью критерия нормальности Шапиро-Уилка и подтверждены распределением Гаусса. Статистический анализ проводили с использованием критерия Стьюдента t с двусторонним доверительным интервалом 95%. Значения P менее 0,05 считались статистически значимыми.
Статистический анализ проводили с использованием критерия Стьюдента t с двусторонним доверительным интервалом 95%. Значения P менее 0,05 считались статистически значимыми.
План реагирования на COVID-19 — Департамент общественного здравоохранения округа Лос-Анджелес
9Последнее обновление страницы 0002: 13.10.2022Текущий уровень сообщества CDC
округа Лос-Анджелес:
низкий
Посмотреть план реагирования округа Лос-Анджелес на COVID-19 (PDF)
Метрики округа Лос-Анджелес | ||
| Новые ящики (на 100 000 человек за последние 7 дней) | новых госпитализаций с COVID-19 на 100 000 населения (всего за 7 дней) | Доля укомплектованных стационарных коек, занятых пациентами с COVID-19 (в среднем за 7 дней) |
| 64,6/100 000 | 4,9 | 2,8% |
Уровень заболеваемости рассчитывается Департаментом общественного здравоохранения округа Лос-Анджелес, чтобы обеспечить более своевременные обновления. Два показателя госпитализации рассчитываются CDC и публикуются на веб-сайте CDC Community Levels. Данные обновлены 13 октября 2022 г.
Два показателя госпитализации рассчитываются CDC и публикуются на веб-сайте CDC Community Levels. Данные обновлены 13 октября 2022 г.
Округ Лос-Анджелес (LAC) будет продолжать использовать матрицу COVID-19 COVID-19 на уровне сообщества Центров по контролю и профилактике заболеваний (CDC) для руководства оценкой риска в округе и информирования о принятии профилактических стратегий (см. Меры по обеспечению безопасности и стратегии предотвращения ниже). Округ Лос-Анджелес внедрит стратегии для более высокого уровня сообщества, когда результаты по обоим больничным показателям в структуре будут на уровне или выше пороговых значений для более высокого уровня сообщества в течение как минимум четырнадцати (14) дней подряд. Стратегии вернутся к более низкому уровню сообщества, когда результат по крайней мере для одного больничного показателя упадет ниже порогового значения для более высокого уровня сообщества в течение как минимум четырнадцати (14) дней подряд.
| Индикаторы | Низкий | Средний | Высокий | |
| Менее 200 | новых госпитализаций с COVID-19 на 100 000 населения (всего за 7 дней) | 10,0-19,9 | ≥20,0 | |
| Доля укомплектованных стационарных коек, занятых пациентами с COVID-19 (в среднем за 7 дней) | 10,0-14,9% | ≥15,0% | ||
| 200 или более | новых госпитализаций с COVID-19 на 100 000 населения (всего за 7 дней) | нет данных | ≥10,0 | |
| Доля укомплектованных стационарных коек, занятых пациентами с COVID-19 (в среднем за 7 дней) | нет данных | ≥10,0% |
(Согласовано с CDC на уровне сообщества и передачей от сообщества)
Приведенные ниже меры безопасности сообщества и стратегии предотвращения отражают усилия CDC, штата и округа по смягчению последствий в четырех областях: маскирование внутри помещений, тестирование, проверка вакцинации и вентиляция. Федеральные требования и требования штата включены в приведенную ниже таблицу и будут изменены, если и когда эти требования изменятся.
Федеральные требования и требования штата включены в приведенную ниже таблицу и будут изменены, если и когда эти требования изменятся.
*Примечание. При наличии циркуляции нового COVID-19вызывающий озабоченность вариант, демонстрирующий потенциальные доказательства отказа от вакцинации, эти рекомендации и требования будут изменены.
Стратегии в таблице охватывают низкий, средний и высокий уровни сообщества. Пожалуйста, , прокрутите по горизонтали , чтобы просмотреть все стратегии на уровне сообщества.
| Уровень сообщества CDC 1 : НИЗКИЙ | Уровень сообщества CDC 1 : СРЕДНИЙ | CDC, уровень сообщества 1 : ВЫСОКИЙ | ||||
| Передача сообщества: Еженедельная заболеваемость | >100/100К | >100/100К | Все уровни передачи | |||
| Маски для помещений Примечание: Работодатели должны предлагать подходящие медицинские маски и респираторы работникам, работающим в помещении в тесном контакте с другими работниками и/или клиентами на всех объектах | Все лица, независимо от прививочного статуса | Индивидуальные предпочтения 3 , если это не требуется на месте | Настоятельно рекомендуется, если это не требуется сайтом | Индивидуальные предпочтения 3 , если это не требуется на месте | Настоятельно рекомендуется, если это не требуется сайтом | Требуется во всех общественных местах и на предприятиях |
| Лица с повышенным риском 2 | Настоятельно рекомендуется в условиях повышенного риска, если только это не требуется сайтом | .Настоятельно рекомендуется, если это не требуется сайтом | Настоятельно рекомендуется в условиях повышенного риска, если только это не требуется сайтом | .Настоятельно рекомендуется, если это не требуется сайтом | Обязательно | |
| Для тех, кто подвергается воздействию | Необходимо носить маску в помещении в течение 10 дней после последнего дня воздействия | Необходимо носить маску в помещении в течение 10 дней после последнего дня воздействия | Необходимо носить маску в помещении в течение 10 дней после последнего дня воздействия | Необходимо носить маску в помещении в течение 10 дней после последнего дня воздействия | Необходимо носить маску в помещении в течение 10 дней после последнего дня воздействия | |
| Для тех, у кого подтвержден положительный результат | Требуется изоляция на срок до 10 дней, если на 6-й день тест не будет отрицательным; маска настоятельно рекомендуется тем, у кого отрицательный результат на 6-10 день; в условиях рабочего места маски требуются в течение полных 10 дней | Требуется изоляция на срок до 10 дней, если на 6-й день тест не будет отрицательным; маска настоятельно рекомендуется тем, у кого отрицательный результат на 6-10 день; в условиях рабочего места маски необходимы в течение полных 10 дней | Требуется изоляция на срок до 10 дней; маска настоятельно рекомендуется для тех, у кого отрицательный результат на 6-10 день, на рабочем месте маски требуются в течение полных 10 дней | Требуется изоляция на срок до 10 дней, если на 6-й день тест не будет отрицательным; маска настоятельно рекомендуется тем, у кого отрицательный результат на 6-10 день; в условиях рабочего места маски требуются в течение полных 10 дней | ||
| Все медицинские учреждения и учреждения коллективного ухода | Обязательно | Обязательно | Обязательно | Обязательно | Обязательно | |
| Исправительные учреждения и места содержания под стражей, приюты для бездомных и временные приюты | Индивидуальные предпочтения 3 за пределами клинических областей, если это не требуется агентством | Настоятельно рекомендуется, если этого не требует агентство | Обязательно | Обязательно | Обязательно | |
| Общественный транспорт, транспортный узел и транспортные услуги (включая маршрутные автобусы, такси, медицинский транспорт) | Настоятельно рекомендуется, если этого не требует агентство | Обязательно | Настоятельно рекомендуется, если этого не требует агентство | Требуется | Обязательно | |
| Тестирование | Обычные скрининговые испытания на рабочих местах | По требованию участка или сектора | По требованию участка или сектора | По требованию участка или сектора | По требованию участка или сектора | По требованию участка или сектора |
| Для тех, кто подвергается воздействию | Требуется, если не помещен в карантин | Требуется, если не помещен в карантин | Требуется, если не помещен в карантин | Требуется, если не помещен в карантин | Требуется, если не помещен в карантин | |
| Для тех, у кого есть симптомы / подтвержденный положительный результат | Требуется для выхода из изоляции на 5-й день; в противном случае Изоляция на 10 дней | Требуется для выхода из изоляции на 5-й день; в противном случае изолировать на 10 дней | Требуется для выхода из изоляции на 5-й день; в противном случае Изоляция на 10 дней | Требуется для выхода из изоляции на 5-й день; в противном случае Изоляция на 10 дней | Требуется для выхода из изоляции на 5-й день; в противном случае Изоляция на 10 дней | |
| Перед общением с людьми из группы повышенного риска | Индивидуальные предпочтения 3 , если это не требуется на месте | Настоятельно рекомендуется, если этого не требует сайт/хост | Индивидуальные предпочтения 3 , если это не требуется на месте | Настоятельно рекомендуется, если этого не требует сайт/хост | Настоятельно рекомендуется, если этого не требует сайт/хост | |
| Вакцинация / Подтверждение отрицательного результата теста | Для медицинских работников, сотрудников медицинских учреждений и сотрудников учреждений коллективного ухода с высоким уровнем риска и приютов для бездомных | Требуется вакцинация | Требуется вакцинация | Требуется вакцинация | Требуется вакцинация | Требуется вакцинация |
| Для посетителей в помещениях медицинских учреждений, включая места массового ухода | Настоятельно рекомендуется, если это не требуется сайтом | Настоятельно рекомендуется, если это не требуется сайтом | Настоятельно рекомендуется, если это не требуется сайтом | Настоятельно рекомендуется, если этого не требует сайт | .Требовать, чтобы все посетители, независимо от статуса вакцинации от COVID, проходили тестирование (ПЦР или антиген) перед посещением помещений | |
| Мероприятия на открытом воздухе | Индивидуальные настройки сайта | Индивидуальные настройки сайта | Индивидуальные настройки сайта | Индивидуальные настройки сайта | Индивидуальные настройки сайта | |
| Мероприятия в помещении | Индивидуальные настройки сайта | Индивидуальные настройки сайта | Индивидуальные настройки сайта | Индивидуальные настройки сайта | Настоятельно рекомендуется, если это не требуется сайтом | |
| Для персонала приютов для бездомных | Требуется вакцинация | Требуется вакцинация | Требуется вакцинация | Требуется вакцинация | Требуется вакцинация | |
| Снижение воздействия на окружающую среду | Вентиляция во всех помещениях | Настоятельно рекомендуется обеспечить и поддерживать улучшенную вентиляцию во всех помещениях | Настоятельно рекомендуется обеспечить и поддерживать улучшенную вентиляцию во всех помещениях | Настоятельно рекомендуется обеспечить и поддерживать улучшенную вентиляцию во всех помещениях | Настоятельно рекомендуется обеспечить и поддерживать улучшенную вентиляцию во всех помещениях | Настоятельно рекомендуется обеспечить и поддерживать улучшенную вентиляцию во всех помещениях |


 Уровень сообщества CDC может быть низким, средним или высоким и определяется более высоким уровнем нового COVID-19.показатели госпитализации и стационарных коек, занятых пациентами с COVID-19, исходя из текущего уровня новых случаев на 100 000 населения за последние 7 дней.
Уровень сообщества CDC может быть низким, средним или высоким и определяется более высоким уровнем нового COVID-19.показатели госпитализации и стационарных коек, занятых пациентами с COVID-19, исходя из текущего уровня новых случаев на 100 000 населения за последние 7 дней. (2) К группе повышенного риска относятся: пожилые люди, непривитые, люди с сопутствующими заболеваниями, люди с ослабленным иммунитетом и люди, живущие в бедных сообществах. Это означает, что человек с одним или несколькими из этих состояний, который тяжело заболел от COVID-19 (имеет тяжелое заболевание от COVID-19), с большей вероятностью: будет госпитализирован, ему потребуется интенсивная терапия, ему потребуется искусственная вентиляция легких, чтобы помочь ему дышать, или умереть. Посетите веб-сайт CDC для получения более подробной информации об основных заболеваниях, связанных с повышенным риском тяжелого течения COVID-19.болезнь.
(3) Индивидуальное предпочтение означает, что ни одному лицу не может быть запрещено носить маску в качестве условия участия в деятельности или начала бизнеса.
Общественное здравоохранение будет отслеживать следующие сигналы, чтобы обеспечить раннее выявление тревожных тенденций, которые могут привести к будущим высоким темпам передачи и/или увеличению тяжести заболевания. Сигналы в приведенной ниже таблице включают три (3) меры для всего сообщества (варианты, вызывающие обеспокоенность,число посещений, кумулятивный уровень заболеваемости в общинах с высоким уровнем бедности) и четыре (4) отраслевых меры (вспышки в учреждениях квалифицированного сестринского ухода [SNFs], вспышки в детских садах до 12-х классов школ, вспышки в учреждениях, оказывающих помощь бездомным [PEH], кластеры на рабочих местах ). Предупреждение по любому показателю, достигающему порога средней или высокой степени беспокойства, вызовет углубленный анализ способствующих факторов и возможность внесения изменений в стратегии профилактики сообщества, как указано в таблице «Приоритетные ответные меры сектора» ниже.
| Пороговые значения индикатора | Низкая забота | Средний концерн | Высокая озабоченность | Текущие значения округа Лос-Анджелес |
Процент секвенированных образцов, идентифицированных как новый интересующий подвариант* (на основе ВОЗ, CDC или местного обозначения) 1 В настоящее время отслеживается: BA. 4.6 4.6 | 10%-20% | > 20% | 4% | |
| Среднее за 7 дней процент обращений в отделение неотложной помощи (ED), классифицированных как связанные с коронавирусом 2 | 5%-10% | > 10% | 4% | |
| 7-дневный кумулятивный показатель заболеваемости с поправкой на возраст для районов с самым низким доходом (30-100% бедности) 3 | 100-200 на 100 000 | > 200 на 100 000 | 48 на 100 000 | |
| Текущая концентрация SARS-CoV2 в сточных водах в процентах от пикового значения концентрации летом 2022 г. 4 | 30-60% | > 60% | 21% | |
| Количество новых вспышек в учреждениях квалифицированного сестринского ухода за последние 7 дней 5 | ≤10 | 11-20 | >20 | 7 |
| Количество новых вспышек в классах школ ТК-12 за последние 7 дней 5 | ≤ 10 | 11-20 | >20 | 11 |
| Количество новых вспышек в условиях ПОБ за последние 7 дней 5 | ≤ 10 | 11-20 | >20 | 5 |
| Количество отчетов кластера рабочей площадки за последние 7 дней 6 | 150-350 | > 350 | 36 |
 Если новый вариант, представляющий интерес, также определяется как вариант с серьезными последствиями на основании обозначения ВОЗ или CDC, это приведет к немедленному обозначению на уровне «высокой озабоченности».
Если новый вариант, представляющий интерес, также определяется как вариант с серьезными последствиями на основании обозначения ВОЗ или CDC, это приведет к немедленному обозначению на уровне «высокой озабоченности». (1) Текущий 7-дневный период: 18.09.22 – 24.09.22.
Варианты пропорций основаны на образце всех положительных по rt-PCR образцов, собранных у жителей округа Лос-Анджелес. В среднем данные секвенирования доступны через 2–3 недели после сбора образцов. Пропорции вариантов рассчитываются с недельными интервалами с использованием даты сбора образцов. Еженедельные оценки могут пересматриваться по мере поступления дополнительных данных.
(2) Текущий 7-дневный период: 03.10.22 — 09.10.22.
Классификация коронавируса определяется произвольным текстовым поиском упоминаний о коронавирусе COVID-19., а также синонимы в выдержках из основных жалоб пациентов и диагнозов из больниц, участвующих в синдромном наблюдении. Некоторые встречи могут быть пропущены из-за неполной и/или запоздалой информации о причине посещения или неправильно классифицированы из-за скрининга на COVID-19 во время посещения, независимо от фактической основной причины посещения. Этот процент от больниц, участвующих в Syndromic Surveillance, не предназначен для точного отражения процента населения, но является полезным инструментом для отслеживания тенденций с течением времени.
Этот процент от больниц, участвующих в Syndromic Surveillance, не предназначен для точного отражения процента населения, но является полезным инструментом для отслеживания тенденций с течением времени.
(3) Текущий 7-дневный период: с 03.10.22 по 09.10.22.
Случаи подсчитываются по дате эпизода, которая является самым ранним существующим значением: даты начала заболевания, даты постановки диагноза, даты смерти, даты получения или даты сбора образцов. Региональная бедность отражает процент домохозяйств, живущих за федеральной чертой бедности или ниже нее. «Районы с самым низким доходом», используемые в этом показателе, представляют собой переписные районы, в которых 30–100% домохозяйств живут за федеральной чертой бедности или ниже нее. Оценки бедности в районе получены на основе пятилетнего (2013–2017 гг.) Обследования американского сообщества переписи населения США на уровне переписных участков. Заболеваемость скорректирована по возрасту и составляет на 100 000 человек. Оценки населения получены из демографических файлов LAC PEPS 2018.
(4) Текущий период для среднего недельного значения: 25.09.22 – 01.10.22.
Средневзвешенное значение рассчитывается с использованием концентраций вируса SARS-CoV-2 в 4 канализационных системах, которые отслеживают вирус в округе Лос-Анджелес, при этом веса представляют численность населения, обслуживаемого каждой канализационной системой. Затем средневзвешенные значения за неделю сравниваются с максимальной средневзвешенной концентрацией вируса SARS-CoV-2 за неделю, наблюдаемой во время всплеска активности летом 2022 года.
(5) Текущий 7-дневный период: с 05.10.22 по 11.10.22.
Подсчеты включают расследования вспышек, начатые за последние 7 дней в учреждениях квалифицированного ухода, школьных классах TK-12 и в местах бездомных. Подсчеты исключают расследования, начатые более чем через 28 дней после даты тестирования или начала заболевания первого случая (для учета задержки с отчетом в DPH). По состоянию на 16.06.22 порог для открытия вспышки в условиях бездомности (PEH) изменился с как минимум 1 случая или 2 человек под следствием на как минимум 2 случая или 3 человека под следствием. По состоянию на 08.07.22 порог для открытия вспышки в условиях ЛПБ изменился как минимум с 2 случаев или 3 человек под следствием на 3 случая или 3 человека под следствием.
По состоянию на 08.07.22 порог для открытия вспышки в условиях ЛПБ изменился как минимум с 2 случаев или 3 человек под следствием на 3 случая или 3 человека под следствием.
(6) Текущий 7-дневный период: с 05.10.22 по 11.10.22.
Кластеры на рабочем месте — это 3 или более случаев в течение 14 дней, о которых сообщает работодатель. Кластеры рабочих мест могут представлять собой вспышки или не вспышки и, если есть дополнительные случаи, могут представлять собой несколько отчетов об одном и том же месте. По состоянию на 10.05.22 крайний срок подачи отчетов за последний день 7-дневного отчетного периода был скорректирован с 23:59 до полудня. 23.05.22 были внесены изменения в обработку данных кластерного отчета, включая исключение повторяющихся отчетов с менее чем 3 случаями.
Для каждого из четырех (4) секторов в приведенной ниже таблице представлены дополнительные общие меры по смягчению последствий и другие меры, которые должны быть реализованы в зависимости от уровня беспокойства, указанного в приведенной выше таблице сигналов раннего оповещения округа Лос-Анджелес. Могут быть рассмотрены дополнительные меры помимо тех, которые перечислены в таблице ниже.
Могут быть рассмотрены дополнительные меры помимо тех, которые перечислены в таблице ниже.
ПРИМЕЧАНИЕ. Все секторы должны соблюдать минимальные требования, касающиеся маскирования помещений, тестирования, проверки вакцинации и вентиляции , изложенные выше в разделе «Меры защиты и стратегии безопасности сообщества».
| Сектор | Общие меры по смягчению последствий | Низкая забота | Средний концерн | Высокая озабоченность |
| ОЯТ* Меры в этой таблице являются общей информацией для населения. Медицинские учреждения должны следовать конкретным рекомендациям LAC DPH и требованиям CDPH и CMS. |
|
| ||
| Переходный детский сад (ТК)-12 класс |
|
|
|
|
| Бездомные (PEH) |
|
|
|
|
| Рабочие площадки |
|
|
|
|
| Сильно пострадавшие сообщества |
|
|
|
|










В соответствии с уровнями сообщества CDC
Большая часть Плана реагирования на COVID-19 зависит от нашей способности сосредоточить ресурсы на ответных действиях и действиях по обеспечению готовности. Реагирующие действия включают в себя устранение растущих уровней риска для населения и сигналы раннего оповещения, которые могут быть специфическими для сектора. Действия по обеспечению готовности сосредоточены на стратегиях, которые позволяют округу последовательно реагировать на меняющиеся условия.
| Действия по обеспечению готовности | Требуется улучшение | Адекватный | Выдающийся | Текущие значения округа Лос-Анджелес |
| Доступ к вакцинам: Количество мобильных пунктов вакцинации в неделю | Менее 200 | 200-300 | Более 300 | 767 (07.10.22 — 14.10.22) |
| Доступ к вакцинам: Количество стационарных пунктов вакцинации | Менее 900 | 900-1100 | Более 1100 | 1040 (05.  10.22 — 11.11.22) 10.22 — 11.11.22) |
| Доступ к вакцинам: Процент соответствующих требованиям проживающих на дому жителей, направленных в DPH и вакцинированных, которые получили вакцину в течение 2 недель с даты направления | Менее 60% | 60%-75% | Более 75% | 88% (11.09.22 — 17.09.22) |
| Доступ к вакцинам: Процент соответствующих критериям жителей 5+ в наиболее уязвимых сообществах 1 , получивших 1+ дополнительную дозу/ревакцинацию | Менее 45% | 45%-60% | Более 60% | Н/Д* |
| Борьба со вспышками: Процент вспышек на рабочих местах с высоким риском 2 , которые были по крайней мере один раз посещены DPH | Менее 75% | 75%-85% | Более 85% | 86% (06.09.22 — 04.10.22) |
| Общедоступный доступ к тестированию: Среднее время ожидания на всех участках тестирования округа | Более 1 часа | От 30 до 60 минут | Менее 30 минут | 10 минут (02.  10.22 — 10.08.22) 10.22 — 10.08.22) |
| Доступ к тестированию в школах: Процент государственных школ TK-12, в которых есть возможности для проведения тестирования 3 | Менее 80% | 80%-90% | Более 90% | 91% (на 11.10.22) |
| Доступ к тестированию в SNF: Процент учреждений квалифицированного сестринского ухода (SNF), которые имеют возможности для планового тестирования реагирования | Менее 90% | 90%-99% | 100% | 100% (02.10.22 — 10.08.22) |
| Доступ к терапевтическим средствам: Количество пунктов в наиболее уязвимых сообществах 1 , которые выдают терапевтические средства 4 | Менее 150 сайтов | 150-250 сайтов | Более 250 сайтов | 557 (по состоянию на 07.10.22) |
| Доступ к терапевтическим средствам: Процент жителей, имеющих право на использование колл-центра и получивших рекомендованные терапевтические средства | Менее 80% | 80%-90% | Более 90% | 100% (27.  09.22 — 03.10.22) 09.22 — 03.10.22) |
| Эпиднадзор – секвенирование: Количество положительных образцов, секвенированных в неделю | Менее 300 | 300-1200 | Более 1200 | 725 (18.09.22 — 24.09.22) |
| Наблюдение – сточные воды: Количество зон планирования обслуживания (SPA), представленных в сборе и тестировании сточных вод 3 раза в неделю | Менее 5 | 5-7 | 8 | 7 (по состоянию на 12.10.22) |
| Эпиднадзор – отделения неотложной помощи: Процент отделений неотложной помощи, сообщающих данные о COVID-подобных заболеваниях | Менее 80% | 80%-90% | Более 90% | 81% (на 09.10.22) |
 09.22 до получения данных от обновленной бустерной вакцины осенью 2022 года. Обновление этого показателя возобновится, как только эти данные станут доступны.
09.22 до получения данных от обновленной бустерной вакцины осенью 2022 года. Обновление этого показателя возобновится, как только эти данные станут доступны. (1) «Наиболее уязвимые» сообщества классифицируются на основе как минимум 11 характеристик сообщества, включая экономические, жилищные, экологические, социальные, образовательные, транспортные и медицинские услуги, собранные в метрике равенства вакцин Департамента общественного здравоохранения штата Калифорния или охват полностью вакцинированным населением меньше, чем общая оценка округа Лос-Анджелес.
(2) Вспышки на рабочих местах с высоким риском определяются как 1) производственные или складские рабочие места с участием не менее 50 сотрудников; 2) установка авиакомпании/аэропорта; или 3) любая отрасль с 5 или более эпидемиологически связанными случаями. Определения вспышек на рабочих местах с высоким риском могут меняться при более высоких уровнях передачи инфекции в сообществе и/или когда количество вспышек превышает местные возможности для проведения расследований.