| AVR для DeMark DMG 8800FE | под заказ |
| Автомат защиты для DeMark DMG 8800FE | в наличии |
| Аккумулятор для DeMark DMG 8800FE | под заказ |
| Бак топливный для DeMark DMG 8800FE | в наличии |
| Барабан стартера для DeMark DMG 8800FE | в наличии |
| Блок диодов для DeMark DMG 8800FE | в наличии |
| Блок заряда АКБ для DeMark DMG 8800FE | в наличии |
| Блок индикации для DeMark DMG 8800FE | под заказ |
| Болт для слива масла для DeMark DMG 8800FE | в наличии |
| Болт крепления клапанной крышки для DeMark DMG 8800FE | под заказ |
| Болт крепления крышки головки цилиндра для DeMark DMG 8800FE | под заказ |
| Болт крепления ротора для DeMark DMG 8800FE | в наличии |
| Болт крышки генератора для DeMark DMG 8800FE | в наличии |
| Болт цилиндра для DeMark DMG 8800FE | под заказ |
| Вал балансира (противовес) для DeMark DMG 8800FE | в наличии |
| Вал рычага регулятора оборотов для DeMark DMG 8800FE | в наличии |
| Виброгаситель для DeMark DMG 8800FE | в наличии |
| Виброгаситель генератора и двигателя для DeMark DMG 8800FE | под заказ |
| Вилка для DeMark DMG 8800FE | в наличии |
| Вилка штепсельная для DeMark DMG 8800FE | под заказ |
| Вольтметр для DeMark DMG 8800FE | в наличии |
| Воронка для заливки масла в картер для DeMark DMG 8800FE | в наличии |
| Выключатель для DeMark DMG 8800FE | в наличии |
| Выключатель зажигания для DeMark DMG 8800FE | под заказ |
| Гайка крепления головки цилиндра для DeMark DMG 8800FE | в наличии |
| Гайка маховика для DeMark DMG 8800FE | под заказ |
| Гайка шпильки цилиндра для DeMark DMG 8800FE | в наличии |
| Генератор для DeMark DMG 8800FE | под заказ |
| Глушитель для DeMark DMG 8800FE | в наличии |
| Головка блока цилиндра для DeMark DMG 8800FE | под заказ |
| Головка блока цилиндра в сборе для DeMark DMG 8800FE | под заказ |
| Датчик импульсный для DeMark DMG 8800FE | в наличии |
| Датчик уровня масла для DeMark DMG 8800FE | под заказ |
| Двигатель для DeMark DMG 8800FE | в наличии |
| Двигатель сервопривода для DeMark DMG 8800FE | под заказ |
| Двигатель шаговый привода заслонки карбюратора для DeMark DMG 8800FE | под заказ |
| Дефлектор для DeMark DMG 8800FE | под заказ |
| Диод для DeMark DMG 8800FE | под заказ |
| Диод якоря генератора для DeMark DMG 8800FE | под заказ |
| Диодный мост для DeMark DMG 8800FE | в наличии |
| Дисплей цифровой для DeMark DMG 8800FE | в наличии |
| Замок зажигания для DeMark DMG 8800FE | под заказ |
| Индикатор уровня топлива для DeMark DMG 8800FE | в наличии |
| Индикатор уровня топлива в баке круглый для DeMark DMG 8800FE | под заказ |
| Индикатор уровня топлива в баке прямоугольный для DeMark DMG 8800FE | в наличии |
| Искрогаситель глушителя для DeMark DMG 8800FE | под заказ |
| Карбюратор для DeMark DMG 8800FE | в наличии |
| Картер для DeMark DMG 8800FE | под заказ |
| Катушка активная для DeMark DMG 8800FE | в наличии |
| Катушка заряда АКБ для DeMark DMG 8800FE | в наличии |
| Катушка зажигания для DeMark DMG 8800FE | в наличии |
| Клапан впускной для DeMark DMG 8800FE | под заказ |
| Клапан впускной и выпускной для DeMark DMG 8800FE | под заказ |
| Клапан выпускной для DeMark DMG 8800FE | в наличии |
| Клапан электромагнитный карбюратора для DeMark DMG 8800FE | под заказ |
| Клемы для АКБ для DeMark DMG 8800FE | под заказ |
| Ключ замка зажигания для DeMark DMG 8800FE | под заказ |
| Кожух маховика для DeMark DMG 8800FE | в наличии |
| Коленвал для DeMark DMG 8800FE | в наличии |
| Колесо для DeMark DMG 8800FE | в наличии |
| Колодка генератора для DeMark DMG 8800FE | под заказ |
| Коллектор глушителя выпускной для DeMark DMG 8800FE | под заказ |
| Колпачок клапана для DeMark DMG 8800FE | под заказ |
| Колпачок клапана выпускного под коромысло для DeMark DMG 8800FE | в наличии |
| Колпачок маслосъемный для DeMark DMG 8800FE | в наличии |
| Колпачок свечи зажигания для DeMark DMG 8800FE | в наличии |
| Кольца поршневые для DeMark DMG 8800FE | в наличии |
| Кольцо стопорное подшипника коленвала для DeMark DMG 8800FE | в наличии |
| Кольцо стопорное поршневого пальца для DeMark DMG 8800FE | в наличии |
| Кольцо уплотнительное щупа уровня масла для DeMark DMG 8800FE | в наличии |
| Комплект колес для DeMark DMG 8800FE | в наличии |
| Конденсатор для DeMark DMG 8800FE | в наличии |
| Контроллер зажигания для DeMark DMG 8800FE | под заказ |
| Коромысло для DeMark DMG 8800FE | под заказ |
| Коромысло клапана для DeMark DMG 8800FE | в наличии |
| Кожух защитный для DeMark DMG 8800FE | в наличии |
| Кожух маховика для DeMark DMG 8800FE | в наличии |
| Кран топливный для DeMark DMG 8800FE | в наличии |
| Кран топливный угловой для DeMark DMG 8800FE | под заказ |
| Крыльчатка для DeMark DMG 8800FE | под заказ |
| Крыльчатка маховика для DeMark DMG 8800FE | под заказ |
| Крыльчатка ротора для DeMark DMG 8800FE | под заказ |
| Крышка генератора для DeMark DMG 8800FE | в наличии |
| Крышка клапанов для DeMark DMG 8800FE | в наличии |
| Крышка панели управления для DeMark DMG 8800FE | в наличии |
| Крышка фильтра воздушного для DeMark DMG 8800FE | под заказ |
| Магнето для DeMark DMG 8800FE | под заказ |
| Маслоотражатель впускного клапана для DeMark DMG 8800FE | в наличии |
| Маховик для DeMark DMG 8800FE | под заказ |
| Механизм управления воздушной заслонкой карбюратора для DeMark DMG 8800FE | в наличии |
| Направляющая двигателя сервопривода для DeMark DMG 8800FE | в наличии |
| Направляющая толкателей клапана для DeMark DMG 8800FE | в наличии |
| Направляющая штанга толкателей клапанов для DeMark DMG 8800FE | в наличии |
| Насос топливный для DeMark DMG 8800FE | под заказ |
| Ножка рамы генератора для DeMark DMG 8800FE | под заказ |
| Опора рамы для генератора для DeMark DMG 8800FE | под заказ |
| Ось крепления колеса для DeMark DMG 8800FE | в наличии |
| Палец поршня для DeMark DMG 8800FE | в наличии |
| Панель контрольная для DeMark DMG 8800FE | под заказ |
| Подшипник игольчатый для DeMark DMG 8800FE | в наличии |
| Подшипник коленвала для DeMark DMG 8800FE | в наличии |
| Подшипник ротора для DeMark DMG 8800FE | под заказ |
| Подшипник ротора со стопорным кольцом для DeMark DMG 8800FE | под заказ |
| Поршень для DeMark DMG 8800FE | в наличии |
| Поршневая группа для DeMark DMG 8800FE | в наличии |
| Пробка топливного бака для DeMark DMG 8800FE | под заказ |
| Провода 12В для DeMark DMG 8800FE | в наличии |
| Провода с разъемами для DeMark DMG 8800FE | под заказ |
| Прокладка воздушного фильтра для DeMark DMG 8800FE | в наличии |
| Прокладка глушителя для DeMark DMG 8800FE | под заказ |
| Прокладка головки цилиндра для DeMark DMG 8800FE | в наличии |
| Прокладка карбюратора для DeMark DMG 8800FE | под заказ |
| Прокладка теплоизолятора для DeMark DMG 8800FE | под заказ |
| Прокладка цилиндра для DeMark DMG 8800FE | в наличии |
| Противовес для DeMark DMG 8800FE | в наличии |
| Пружина для DeMark DMG 8800FE | под заказ |
| Пружина карбюратора для DeMark DMG 8800FE | в наличии |
| Пружина клапана для DeMark DMG 8800FE | под заказ |
| Пружина ручного стартера для DeMark DMG 8800FE | под заказ |
| Радиатор блока диодов для DeMark DMG 8800FE | под заказ |
| Рама для DeMark DMG 8800FE | в наличии |
| Распредвал для DeMark DMG 8800FE | в наличии |
| Реле датчика уровня масла для DeMark DMG 8800FE | в наличии |
| Реле отключения магнето для DeMark DMG 8800FE | под заказ |
| Реле стартера для DeMark DMG 8800FE | в наличии |
| Ремкомплект для ручного стартера для DeMark DMG 8800FE | под заказ |
| Розетка для DeMark DMG 8800FE | в наличии |
| Ротор (якорь) генератора для DeMark DMG 8800FE | в наличии |
| Ручка для переноски для DeMark DMG 8800FE | в наличии |
| Ручка ручного стартера для DeMark DMG 8800FE | под заказ |
| Рычаг воздушной заслонки карбюратора для DeMark DMG 8800FE | в наличии |
| Рычаг регулятора оборотов для DeMark DMG 8800FE | в наличии |
| Сальник для DeMark DMG 8800FE | под заказ |
| Сальник коленвала для DeMark DMG 8800FE | под заказ |
| Сапун, патрубок для DeMark DMG 8800FE | под заказ |
| Свеча зажигания для DeMark DMG 8800FE | в наличии |
| Седло пружины клапана для DeMark DMG 8800FE | под заказ |
| Собачка стартера для DeMark DMG 8800FE | под заказ |
| Стартер ручной для DeMark DMG 8800FE | под заказ |
| Статор генератора для DeMark DMG 8800FE | под заказ |
| Стопор пружины клапана для DeMark DMG 8800FE | в наличии |
| Счетчик моточасов для DeMark DMG 8800FE | под заказ |
| Сухарь клапана для DeMark DMG 8800FE | под заказ |
| Тарелка пружины клапана для DeMark DMG 8800FE | в наличии |
| Тарелка толкателя клапана для DeMark DMG 8800FE | в наличии |
| Теплоизолятор для DeMark DMG 8800FE | под заказ |
| Теплоизолятор карбюратора для DeMark DMG 8800FE | под заказ |
| Толкатель клапана для DeMark DMG 8800FE | в наличии |
| Тяга регулятора оборотов для DeMark DMG 8800FE | под заказ |
| Фильтр воздушный для DeMark DMG 8800FE | под заказ |
| Фильтр масляный для DeMark DMG 8800FE | в наличии |
| Фильтр топливный для DeMark DMG 8800FE | в наличии |
| Храповик стартера для DeMark DMG 8800FE | под заказ |
| Цилиндр для DeMark DMG 8800FE | в наличии |
| Шатун для DeMark DMG 8800FE | под заказ |
| Шатунно-поршневая группа для DeMark DMG 8800FE | под заказ |
| Шестерня механического регулятора для DeMark DMG 8800FE | в наличии |
| Шестерня регулятора оборотов для DeMark DMG 8800FE | под заказ |
| Шланг вакуумного регулятора с клапаном для DeMark DMG 8800FE | в наличии |
| Шланг топливный для DeMark DMG 8800FE | в наличии |
| Шланг сапуна для DeMark DMG 8800FE | под заказ |
| Шнур ручного стартера для DeMark DMG 8800FE | в наличии |
| Шпилька головки цилиндра для DeMark DMG 8800FE | под заказ |
| Шпилька крепления карбюратора для DeMark DMG 8800FE | в наличии |
| Шпонка маховика для DeMark DMG 8800FE | в наличии |
| Штанга толкателя клапана для DeMark DMG 8800FE | в наличии |
| Штифт головки цилиндра для DeMark DMG 8800FE | в наличии |
| Штифт крышки картера для DeMark DMG 8800FE | под заказ |
| Штуцер топливного шланга для DeMark DMG 8800FE | под заказ |
| Щетки коллекторные для DeMark DMG 8800FE | в наличии |
| Щуп уровня масла для DeMark DMG 8800FE | под заказ |
| AVR для DeMark DMG 3500FE | в наличии |
| Автомат защиты для DeMark DMG 3500FE | в наличии |
| Аккумулятор для DeMark DMG 3500FE | в наличии |
| Бак топливный для DeMark DMG 3500FE | в наличии |
| Барабан стартера для DeMark DMG 3500FE | под заказ |
| Блок диодов для DeMark DMG 3500FE | под заказ |
| Блок заряда АКБ для DeMark DMG 3500FE | в наличии |
| Блок индикации для DeMark DMG 3500FE | в наличии |
| Болт для слива масла для DeMark DMG 3500FE | в наличии |
| Болт крепления клапанной крышки для DeMark DMG 3500FE | в наличии |
| Болт крепления крышки головки цилиндра для DeMark DMG 3500FE | под заказ |
| Болт крепления ротора для DeMark DMG 3500FE | в наличии |
| Болт крышки генератора для DeMark DMG 3500FE | в наличии |
| Болт цилиндра для DeMark DMG 3500FE | под заказ |
| Вал балансира (противовес) для DeMark DMG 3500FE | под заказ |
| Вал рычага регулятора оборотов для DeMark DMG 3500FE | в наличии |
| Виброгаситель для DeMark DMG 3500FE | под заказ |
| Виброгаситель генератора и двигателя для DeMark DMG 3500FE | в наличии |
| Вилка для DeMark DMG 3500FE | под заказ |
| Вилка штепсельная для DeMark DMG 3500FE | под заказ |
| Вольтметр для DeMark DMG 3500FE | в наличии |
| Воронка для заливки масла в картер для DeMark DMG 3500FE | в наличии |
| Выключатель для DeMark DMG 3500FE | в наличии |
| Выключатель зажигания для DeMark DMG 3500FE | под заказ |
| Гайка крепления головки цилиндра для DeMark DMG 3500FE | в наличии |
| Гайка маховика для DeMark DMG 3500FE | под заказ |
| Гайка шпильки цилиндра для DeMark DMG 3500FE | в наличии |
| Генератор для DeMark DMG 3500FE | под заказ |
| Глушитель для DeMark DMG 3500FE | под заказ |
| Головка блока цилиндра для DeMark DMG 3500FE | в наличии |
| Головка блока цилиндра в сборе для DeMark DMG 3500FE | в наличии |
| Датчик импульсный для DeMark DMG 3500FE | под заказ |
| Датчик уровня масла для DeMark DMG 3500FE | в наличии |
| Двигатель для DeMark DMG 3500FE | под заказ |
| Двигатель сервопривода для DeMark DMG 3500FE | в наличии |
| Двигатель шаговый привода заслонки карбюратора для DeMark DMG 3500FE | в наличии |
| Дефлектор для DeMark DMG 3500FE | в наличии |
| Диод для DeMark DMG 3500FE | под заказ |
| Диод якоря генератора для DeMark DMG 3500FE | в наличии |
| Диодный мост для DeMark DMG 3500FE | в наличии |
| Дисплей цифровой для DeMark DMG 3500FE | под заказ |
| Замок зажигания для DeMark DMG 3500FE | в наличии |
| Индикатор уровня топлива для DeMark DMG 3500FE | под заказ |
| Индикатор уровня топлива в баке круглый для DeMark DMG 3500FE | в наличии |
| Индикатор уровня топлива в баке прямоугольный для DeMark DMG 3500FE | под заказ |
| Искрогаситель глушителя для DeMark DMG 3500FE | под заказ |
| Карбюратор для DeMark DMG 3500FE | в наличии |
| Картер для DeMark DMG 3500FE | под заказ |
| Катушка активная для DeMark DMG 3500FE | под заказ |
| Катушка заряда АКБ для DeMark DMG 3500FE | под заказ |
| Катушка зажигания для DeMark DMG 3500FE | под заказ |
| Клапан впускной для DeMark DMG 3500FE | под заказ |
| Клапан впускной и выпускной для DeMark DMG 3500FE | в наличии |
| Клапан выпускной для DeMark DMG 3500FE | под заказ |
| Клапан электромагнитный карбюратора для DeMark DMG 3500FE | под заказ |
| Клемы для АКБ для DeMark DMG 3500FE | под заказ |
| Ключ замка зажигания для DeMark DMG 3500FE | под заказ |
| Кожух маховика для DeMark DMG 3500FE | под заказ |
| Коленвал для DeMark DMG 3500FE | под заказ |
| Колесо для DeMark DMG 3500FE | в наличии |
| Колодка генератора для DeMark DMG 3500FE | в наличии |
| Коллектор глушителя выпускной для DeMark DMG 3500FE | под заказ |
| Колпачок клапана для DeMark DMG 3500FE | под заказ |
| Колпачок клапана выпускного под коромысло для DeMark DMG 3500FE | под заказ |
| Колпачок маслосъемный для DeMark DMG 3500FE | под заказ |
| Колпачок свечи зажигания для DeMark DMG 3500FE | в наличии |
| Кольца поршневые для DeMark DMG 3500FE | в наличии |
| Кольцо стопорное подшипника коленвала для DeMark DMG 3500FE | под заказ |
| Кольцо стопорное поршневого пальца для DeMark DMG 3500FE | под заказ |
| Кольцо уплотнительное щупа уровня масла для DeMark DMG 3500FE | в наличии |
| Комплект колес для DeMark DMG 3500FE | в наличии |
| Конденсатор для DeMark DMG 3500FE | под заказ |
| Контроллер зажигания для DeMark DMG 3500FE | под заказ |
| Коромысло для DeMark DMG 3500FE | под заказ |
| Коромысло клапана для DeMark DMG 3500FE | под заказ |
| Кожух защитный для DeMark DMG 3500FE | в наличии |
| Кожух маховика для DeMark DMG 3500FE | в наличии |
| Кран топливный для DeMark DMG 3500FE | в наличии |
| Кран топливный угловой для DeMark DMG 3500FE | в наличии |
| Крыльчатка для DeMark DMG 3500FE | под заказ |
| Крыльчатка маховика для DeMark DMG 3500FE | в наличии |
| Крыльчатка ротора для DeMark DMG 3500FE | под заказ |
| Крышка генератора для DeMark DMG 3500FE | под заказ |
| Крышка клапанов для DeMark DMG 3500FE | в наличии |
| Крышка панели управления для DeMark DMG 3500FE | под заказ |
| Крышка фильтра воздушного для DeMark DMG 3500FE | под заказ |
| Магнето для DeMark DMG 3500FE | в наличии |
| Маслоотражатель впускного клапана для DeMark DMG 3500FE | в наличии |
| Маховик для DeMark DMG 3500FE | под заказ |
| Механизм управления воздушной заслонкой карбюратора для DeMark DMG 3500FE | в наличии |
| Направляющая двигателя сервопривода для DeMark DMG 3500FE | в наличии |
| Направляющая толкателей клапана для DeMark DMG 3500FE | под заказ |
| Направляющая штанга толкателей клапанов для DeMark DMG 3500FE | в наличии |
| Насос топливный для DeMark DMG 3500FE | в наличии |
| Ножка рамы генератора для DeMark DMG 3500FE | под заказ |
| Опора рамы для генератора для DeMark DMG 3500FE | под заказ |
| Ось крепления колеса для DeMark DMG 3500FE | в наличии |
| Палец поршня для DeMark DMG 3500FE | под заказ |
| Панель контрольная для DeMark DMG 3500FE | в наличии |
| Подшипник игольчатый для DeMark DMG 3500FE | под заказ |
| Подшипник коленвала для DeMark DMG 3500FE | в наличии |
| Подшипник ротора для DeMark DMG 3500FE | под заказ |
| Подшипник ротора со стопорным кольцом для DeMark DMG 3500FE | в наличии |
| Поршень для DeMark DMG 3500FE | под заказ |
| Поршневая группа для DeMark DMG 3500FE | в наличии |
| Пробка топливного бака для DeMark DMG 3500FE | в наличии |
| Провода 12В для DeMark DMG 3500FE | в наличии |
| Провода с разъемами для DeMark DMG 3500FE | под заказ |
| Прокладка воздушного фильтра для DeMark DMG 3500FE | в наличии |
| Прокладка глушителя для DeMark DMG 3500FE | в наличии |
| Прокладка головки цилиндра для DeMark DMG 3500FE | под заказ |
| Прокладка карбюратора для DeMark DMG 3500FE | в наличии |
| Прокладка теплоизолятора для DeMark DMG 3500FE | в наличии |
| Прокладка цилиндра для DeMark DMG 3500FE | под заказ |
| Противовес для DeMark DMG 3500FE | под заказ |
| Пружина для DeMark DMG 3500FE | под заказ |
| Пружина карбюратора для DeMark DMG 3500FE | в наличии |
| Пружина клапана для DeMark DMG 3500FE | в наличии |
| Пружина ручного стартера для DeMark DMG 3500FE | под заказ |
| Радиатор блока диодов для DeMark DMG 3500FE | под заказ |
| Рама для DeMark DMG 3500FE | в наличии |
| Распредвал для DeMark DMG 3500FE | в наличии |
| Реле датчика уровня масла для DeMark DMG 3500FE | под заказ |
| Реле отключения магнето для DeMark DMG 3500FE | в наличии |
| Реле стартера для DeMark DMG 3500FE | в наличии |
| Ремкомплект для ручного стартера для DeMark DMG 3500FE | под заказ |
| Розетка для DeMark DMG 3500FE | в наличии |
| Ротор (якорь) генератора для DeMark DMG 3500FE | в наличии |
| Ручка для переноски для DeMark DMG 3500FE | в наличии |
| Ручка ручного стартера для DeMark DMG 3500FE | в наличии |
| Рычаг воздушной заслонки карбюратора для DeMark DMG 3500FE | в наличии |
| Рычаг регулятора оборотов для DeMark DMG 3500FE | в наличии |
| Сальник для DeMark DMG 3500FE | в наличии |
| Сальник коленвала для DeMark DMG 3500FE | под заказ |
| Сапун, патрубок для DeMark DMG 3500FE | под заказ |
| Свеча зажигания для DeMark DMG 3500FE | под заказ |
| Седло пружины клапана для DeMark DMG 3500FE | под заказ |
| Собачка стартера для DeMark DMG 3500FE | в наличии |
| Стартер ручной для DeMark DMG 3500FE | в наличии |
| Статор генератора для DeMark DMG 3500FE | в наличии |
| Стопор пружины клапана для DeMark DMG 3500FE | в наличии |
| Счетчик моточасов для DeMark DMG 3500FE | под заказ |
| Сухарь клапана для DeMark DMG 3500FE | под заказ |
| Тарелка пружины клапана для DeMark DMG 3500FE | под заказ |
| Тарелка толкателя клапана для DeMark DMG 3500FE | в наличии |
| Теплоизолятор для DeMark DMG 3500FE | под заказ |
| Теплоизолятор карбюратора для DeMark DMG 3500FE | в наличии |
| Толкатель клапана для DeMark DMG 3500FE | под заказ |
| Тяга регулятора оборотов для DeMark DMG 3500FE | в наличии |
| Фильтр воздушный для DeMark DMG 3500FE | в наличии |
| Фильтр масляный для DeMark DMG 3500FE | под заказ |
| Фильтр топливный для DeMark DMG 3500FE | в наличии |
| Храповик стартера для DeMark DMG 3500FE | в наличии |
| Цилиндр для DeMark DMG 3500FE | в наличии |
| Шатун для DeMark DMG 3500FE | под заказ |
| Шатунно-поршневая группа для DeMark DMG 3500FE | в наличии |
| Шестерня механического регулятора для DeMark DMG 3500FE | в наличии |
| Шестерня регулятора оборотов для DeMark DMG 3500FE | под заказ |
| Шланг вакуумного регулятора с клапаном для DeMark DMG 3500FE | под заказ |
| Шланг топливный для DeMark DMG 3500FE | в наличии |
| Шланг сапуна для DeMark DMG 3500FE | в наличии |
| Шнур ручного стартера для DeMark DMG 3500FE | в наличии |
| Шпилька головки цилиндра для DeMark DMG 3500FE | под заказ |
| Шпилька крепления карбюратора для DeMark DMG 3500FE | в наличии |
| Шпонка маховика для DeMark DMG 3500FE | под заказ |
| Штанга толкателя клапана для DeMark DMG 3500FE | под заказ |
| Штифт головки цилиндра для DeMark DMG 3500FE | в наличии |
| Штифт крышки картера для DeMark DMG 3500FE | под заказ |
| Штуцер топливного шланга для DeMark DMG 3500FE | под заказ |
| Щетки коллекторные для DeMark DMG 3500FE | в наличии |
| Щуп уровня масла для DeMark DMG 3500FE | в наличии |

DeMARK DMG 6800F – Генераторы бензиновые – GoodGarden.
 ru
ruОписание
Безно-генератор DeMARK DMG 6800F
Портативная электростанция от производителя DeMark, который становится все более популярным на рынке бензиновых генераторов. Силовая техника под брендом DeMark выделяется высоким качеством и способна обеспечивать электроэнергией все основные энергопотребляющие приборы и различную технику.
Технические характеристики
| Максимальная мощность | 5.5 [кВт] |
| Номинальная мощность | 5.0 [кВт] |
| Объем двигателя | 389 [см3] |
| Объем картера | 1.1 [л] |
| Выходная мощность | 13 [л.с.] |
| Объем топливного бака | 25 [л] |
| Время работы при 50% нагрузке | 15 [ч] |
| Напряжение | 230 [В] |
| Частота | 50 [Гц] |
| Розетки | 1-16/1-32 [А] |
| Уровень шума | 69 [дБ] |
| Габариты | 690x530x590 [мм] |
| Вес | 82 [кг] |
- Двигатель – бензиновый, 4-тактный
- Запуск – ручной
- Защитная окраска бака – Car Painting
- Обмотка альтернатора – медная
- Автоматический регулятор напряжения – есть
- Выход 12В/8.
 3А – есть
3А – есть - Антивибрационные ножки подставки
- Усиленная рама
- Усиленные подушки крепления ДВС
- Транспортировочные колеса – есть
- Ручки – складные
- Датчик низкого уровня масла с функцией аварийной остановки двигателя
- Современный дисплей – счетчик моточасов, напряжение, частота, счетчик времени рабочей сессии
- Гарантия – 1 год
Особенности и преимущества
Дизельные генераторы Pramac с доставкой — большой выбор электростанций на официальном сайте www.sklad-generator.ru
Сегодня концерн, выпускающий бензиновые и дизельные генераторы Pramac, является ведущей компанией отрасли в Европе. ДГУ Pramac, которые предлагает ГК «ЭнергоПроф», высоко оценивают потребители и эксперты за оптимальное соотношение цены и качества. О высоком доверии к продукции этого производителя со стороны российских покупателей свидетельствует постоянный рост продаж Pramac электростанций дизельных и прочей техники итальянского производства.
Дизельные электростанции Pramac от компании ЭнергоПроф
Наша компания рада предложить своим клиентам в Москве современное, надежное и доступное оборудование, которое может быть использовано в качестве альтернативного или резервного источника энергии. Среди прочего, нами реализуются установки итальянского производства. Вы можете купить дизельные электростанции Pramac или оборудование, работающее на светлых сортах топлива, которое выпускается заводом более 50 лет. Столь внушительный возраст предприятия красноречиво свидетельствует о конкурентоспособности его продукции на мировом рынке и массе позитивных потребительских качеств.
Модельный ряд оборудования данного бренда рассчитан на самый широкий круг потребителей. Это установки различной производительности, габаритов и других технических параметров. В ассортименте представлены однофазные и трехфазные электростанции.
Дизельный генератор Pramac
Это установки, которые широко используются строительными фирмами, коммунальными предприятиями и промышленными компаниями Москвы. Pramac электростанции дизельные экономичны, надежны, неприхотливы. Относительно высокая стоимость электростанций с лихвой окупается уже в первые месяцы активной эксплуатации.
Генераторы Stamford AvK
Jump to Navigation- Информация
- Производители
- Каталог
- Назад
- Насосное оборудование
- Насосы центробежные
- Apex Pumps
- Насосы винтовые
- Насосы высокого давления
- BFT
- GEA
- Погружные насосы
- Houttuin
- Горизонтальные насосы
- Apex Pumps
- Houttuin
- Inoxihp
- Moyno
- Vipom
- Насосы герметичные
- Hermetic Pumpen
- Zenith
- Насосное оборудование прочее
- AX System
- Sanco
- Servi Group
- Насосы центробежные
- Фильтровальное оборудование
- Воздушные фильтры
- AAF
- Jonell
- Масляные и гидравлические фильтры
- Parker Hannifin Corporation
- Servi Group
- Коалесцирующие фильтры
- ASCO Filtri
- Buhler Technologies
- EUROFILL
- Hydac
- Jonell
- Petrogas
- Scam Filltres
- Vokes Air
- Водоподготовка
- Grunbeck
- Фильтры КВОУ
- AAF
- Осушители
- Воздушные фильтры
- Компрессорное оборудование
- Поршневые компрессоры
- Винтовые компрессоры
- GEA
- Howden
- Stewart & Stevenson
- Центробежные компрессоры
- Baker Hughes
- Stewart & Stevenson
- Thermodyn
- Поршневые компрессоры
- Трубопроводная арматура
- Запорная, регулирующая, запорно-регулирующая арматура
- Предохранительная арматура
- Sapag Industrial valves
- Schroedahl
- Servi Group
- Приводы трубопроводной арматуры
- Biffi
- Keystone
- Запорная, регулирующая, запорно-регулирующая арматура
- Гидравлика
- Гидроцилиндры
- Servi Group
- Гидроклапаны
- Meggitt
- Servi Group
- Гидронасосы
- Riverhawk
- Servi Group
- Гидрораспределители
- Servi Group
- Пневмоцилиндры
- Artec
- Mec Fluid 2
- Гидроцилиндры
- Станочное оборудование
- Станки шлифовальные
- LOESER
- Хонинговальные станки
- CAR srl
- Станки зубо- и резьбо- обрабатывающие
- Nagel Maschinen
- Карусельные станки
- Star Micronics
- Шпиндели и фрезерные головки
- Cytec
- Станки шлифовальные
- Приводная техника
- Электрические приводы
- Servi Group
- Гидравлические приводы
- Biffi
- Пневматические приводы
- Keystone
- Вентиляторы
- Reitz
- Электромагнитные приводы
- Danfoss
- ECONTROL
- Редукторы
- Renk
- VAR-SPE
- Турборедукторы
- Flender-Graffenstaden
- Renk
- Электрические приводы
- КИП (измерительное оборудование)
- Анализаторы влажности
- Belimo
- Scantech
- Приборы измерения уровня
- Endress+Hauser
- Приборы контроля и регулирования технологических процессов
- Reuter-Stokes
- Приборы измерения уровня расхода (расходомеры)
- Belimo
- Itron
- Servi Group
- Системы измерения неразрушающего контроля
- HBM
- Kavlico
- Marposs
- Устройства измерения температуры
- Устройства измерения давления
- Autrol
- Servi Group
- Устройства измерения перемещения и положения
- Анализаторы влажности
- Лабораторное оборудование
- Микроскопия и спектроскопия
- Keyence
- Микроскопия и спектроскопия
- Электрооборудование
- Аккумуляторные батареи
- Hoppecke
- Противопожарное оборудование
- Reuter-Stokes
- Sanco
- Spectrex
- Выключатели
- Metrol
- Источники питания
- LAM Technologies
- Кабели и коннекторы
- Axon’ Cable
- HiRel Connectors
- Murrplastik
- RIO
- Лампы
- Nic
- Parat
- Серийные преобразователи
- LAM Technologies
- Электродвигатели
- Gamak Motors
- LAM Technologies
- Электроника
- DUCATI Energia
- JOVYATLAS
- Luvata
- Murrplastik
- Аккумуляторные батареи
- Прочее оборудование
- Абразивные изделия
- Abrasivos Manhattan
- Atto Abrasives
- Буровое оборудование
- BVM Corporation
- Den-Con Tool
- MI Swaco
- Top-co
- WestCo
- Валы
- GKN
- Jaure
- Rotar
- Вибротехника
- JOST
- Газовые турбины
- Alba Power
- Baker Hughes
- Meggitt
- Score Energy
- Siemens energy
- Solar turbines
- Горелки
- Зажимные устройства
- Restech Norway
- SPIETH
- Защита от износа, налипания, коррозии
- Rema Tip Top
- Инструмент
- Deprag
- Knipex
- Клапаны
- Baker Hughes
- Mec Fluid 2
- Top-co
- Velan
- W.
 T.A.
T.A. - Zimmermann & Jansen (Z&J)
- Крановое оборудование
- Facco
- Маркировочное оборудование
- Couth
- Espera
- Мельницы
- Eirich
- Металлообработка
- Agrati
- Муфты
- Coremo Ocmea
- Esco Couplings
- Jaure
- John Crane
- Kendrion Linnig
- Top-co
- ZERO-MAX
- Оси
- Jaure
- Подшипники
- John Crane
- NTN-SNR
- SPIETH
- Производственные линии
- Espera
- FIBRO
- Masa Henke
- Робототехника
- Motoman Robotics
- Системы обогрева
- Helios
- TYCO Thermal Controls
- Системы охлаждения
- Gohl
- Системы смазки
- Lincoln
- Строительные леса
- HAKI
- Сушильные печи
- Eirich
- Такелажное оборудование
- Casar
- Easy Mover
- Fetra
- Тормоза и сцепления
- Coremo Ocmea
- Упаковочное оборудование
- Espera
- Thimonnier
- Уплотнения
- Flexitallic
- John Crane
- Форсунки и эжекторы
- Exair
- Центраторы
- Top-co
- Электрографитовые щетки
- Morgan Advanced Materials
- Абразивные изделия
- AX System
- A.
 O. Smith – Century Electric
O. Smith – Century Electric - A.S.T.
- AAF
- Abrasivos Manhattan
- Advanced Energy
- Agilent Technologies
- Agrati
- Alba Power
- Algi
- Allweiler
- Alphatron Marine
- Amot
- Anderson Greenwood
- Apex Pumps
- Apollo Valves
- Ariana Industrie
- Ariel
- Artec
- ASCO Filtri
- Ashcroft
- ATAS elektromotory
- Atos
- Atto Abrasives
- Autrol
- Autronica
- Axis
- Axon’ Cable
- Baker Hughes
- Baker Hughes
- Bando
- Baruffaldi
- BAUER Kompressoren
- Belimo
- Bently Nevada
- Berarma
- BFT
- BHDT
- Biffi
- Bifold Group
- Brinkmann pumps
- Buhler Technologies
- BVM Corporation
- Camfil FARR
- Campen Machinery
- CanaWest Technologies
- CAR srl
- Carif
- Casar
- CAT
- Celduc Relais
- Center Line
- Clif Mock
- Comagrav
- Compressor Controls Corporation
- CoorsTek
- Coral engineering
- Coremo Ocmea
- Couth
- CRANE
- Crosby
- Cytec
- Danaher Motion
- Danfoss
- Danobat Group
- David Brown Hydraulics
- Den-Con Tool
- DenimoTECH
- Deprag
- Destaco
- Dixon Valve
- Donaldson
- Donaldson осушители, адсорбенты
- DUCATI Energia
- Duplomatic
- Duplomatic Oleodinamica
- Dustcontrol
- Dynasonics
- E-tech Machinery
- Easy Mover
- Ebro Armaturen
- ECONTROL
- Eirich
- EMIT
- Endress+Hauser
- Esco Couplings
- Espera
- Estarta
- Euchner
- EUROFILL
- EuroSMC
- Exair
- Facco
- FANUC
- Farris
- Fema
- Ferjovi
- Fetra
- FIBRO
- Fisher
- Flender-Graffenstaden
- Flexitallic
- Flowserve
- Fluenta
- Flux
- FPZ
- Freudenberg
- Fritz STUDER
- Gali
- Gamak Motors
- GEA
- GEORGIN
- GKN
- Gohl
- Goulds Pumps
- GPM Titan International
- Graco
- Grunbeck
- Grundfos
- Gustav Gockel
- HAKI
- Harting technology
- HAWE Hydraulik SE
- HBM
- Heimbach
- Helios
- Hermetic Pumpen
- Herose
- HiRel Connectors
- Hohner
- Holland-Controls
- Honsberg Instruments
- Hoppecke
- Horton
- Houttuin
- Howden
- Howden CKD Compressors s.
 r.o.
r.o. - HTI-Gesab
- Hydac
- Hydrotechnik
- IMO
- Inoxihp
- iNPIPE Products
- ISOG
- Italmagneti
- Itron
- ITW Dynatec
- Jaure
- JDSU
- Jenoptik
- John Crane
- Jonell
- JOST
- JOVYATLAS
- K-TEK
- Kadia
- Kavlico
- Kellenberger
- Kendrion
- Kendrion Linnig
- Keyence
- Keystone
- Kitagawa
- Knipex
- Knoll
- Kordt
- Krombach Armaturen
- KSB
- Kumera
- Labor Security System
- LAM Technologies
- Lapmaster Wolters
- Lincoln
- LOESER
- Lufkin Industries
- Luvata
- Mahle
- Marposs
- Masa Henke
- Masoneilan
- Mec Fluid 2
- MEDIT Inc.

- Meggitt
- Mercotac
- Metrol
- MI Swaco
- Minco
- MMC International Corporation
- MOOG
- Moore Industries
- Morgan Advanced Materials
- Motoman Robotics
- Moyno
- Mud King
- MULTISERW-Morek
- Munters
- Murr elektronik
- Murrplastik
- Nagel Maschinen
- National Oilwell Varco
- Netzsch
- Nexoil srl
- Nic
- NOV Mono
- NTN-SNR
- Ntron
- Nuovo Pignone
- O’Drill/MCM
- Oerlikon
- Oilgear
- Omal Automation
- Omni Flow Computers
- OMT
- Opcon
- Orange Research
- Orwat filtertechnik
- OTECO
- Pacific valves
- Pageris AG
- Paktech
- PALL
- Panametrics
- Parat
- Parker Hannifin Corporation
- PENTAIR
- Peter Wolters
- Petrogas
- ProMinent
- Quick Soldering
- Reitz
- Rema Tip Top
- Renk
- Renold
- Repar2
- Resatron
- Resistoflex
- Restech Norway
- Reuter-Stokes
- Revo
- Rexnord
- Rheonik
- Rineer Hydraulics
- RIO
- Riverhawk
- RMG Honeywell
- Ro-Flo Compressors
- Robbi
- ROS
- Rota Engineering
- Rotar
- Rotoflow
- Rotork
- Ruhrpumpen
- S.
 Himmelstein
Himmelstein - Sanco
- Sapag Industrial valves
- Saunders
- Scam Filltres
- Scantech
- Schroedahl
- Score Energy
- Sermas Industrie
- Servi Group
- Settima
- Siekmann Econosto
- Siemens
- Siemens energy
- Simaco
- Solar turbines
- Solberg
- SOR
- Spectrex
- SPIETH
- SPX
- Stamford | AvK
- Star Micronics
- Stewart & Stevenson
- Stockham
- Sumitomo
- Supertec Machinery
- Tamagawa Seiki
- Tartarini
- TEAT
- TEKA
- Thermodyn
- Thimonnier
- Top-co
- Truflo
- Turbotecnica
- Tuthill
- TYCO Thermal Controls
- Vanessa
- VAR-SPE
- VDO
- Velan
- Versa
- Vibra Schultheis
- Vipom
- Vokes Air
- Voumard
- W.
 T.A.
T.A. - Warren
- Waukesha
- Weatherford
- Weiss GmbH
- Wenglor
- WestCo
- Woodward
- Xomox
- Yarway
- Zenith
- ZERO-MAX
- Zimmermann & Jansen (Z&J)
Бесплатный генератор записей DMARC — Создание записи DMARC
Шаг
Для какого домена вы хотите создать запись?
Введите ваш домен
Какой тип политики DMARC вам нужен?
Как вы хотите обрабатывать почту, не прошедшую проверку DMARC?
Пока ничего, просто соберите данные.
Поместить его в карантин для дальнейшего анализа.
Отклонить сразу.
DMARC позволяет применять различные «политики» к электронной почте, которая не соответствует вашему домену.При первой публикации вашей записи мы предлагаем вам начать с «нет». Это позволяет собирать данные, не влияя на потоки электронной почты.
Куда вы хотите отправлять сводные отчеты?
Данные — движущая сила DMARC. Если адрес указан, сводные отчеты DMARC будут доставлены на указанный адрес электронной почты для дальнейшей обработки.
Вы можете продолжить работу с этим мастером, если хотите получать отчеты напрямую, или вы можете создать бесплатную учетную запись dmarcian, и мы создадим вашу запись для вас.
Если вы хотите получить отчет напрямую, вам все равно понадобится способ визуализации данных. Добавьте в закладки наш инструмент преобразования XML в Human после того, как вы начали получать отчеты.
Добавьте в закладки наш инструмент преобразования XML в Human после того, как вы начали получать отчеты.
DMARC Сводные отчеты данных имеют решающее значение для оценки, мониторинга и защиты ваших почтовых потоков. Эти сводные отчеты на основе XML создаются с 24-часовым циклом и включают исчерпывающую статистику о том, как получатель электронной почты видит, что ваш домен используется, включая всю электронную почту, которая полностью прошла DMARC.
Пользователи Dmarcian могут автоматически получать и обрабатывать эти отчеты по мере их создания и просматривать их в Обзоре домена.
Хотите получать индивидуальные отчеты о сбоях?
Индивидуальные отчеты о сбоях или отчеты судебной экспертизы — это копии отдельных частей электронной почты, не прошедших проверку DMARC. Эти отчеты не требуются или необходимы для развертывания DMARC, но могут дать дополнительную информацию о возможном злоупотреблении вашим доменом.
№
Есть
Изучите электронную почту, создав отчеты Forensic DMARC.Хотя это не требуется для установки DMARC, он может быть полезен при выявлении источников или шаблонов злоупотреблений электронной почтой. Для получения дополнительной информации об отчетах DMARC Forensic ознакомьтесь с этой статьей.
Dmarcian предоставляет адрес электронной почты для зарегистрированных пользователей, по которому отчеты судебной экспертизы могут доставляться и анализироваться в Forensic Viewer.
DMARC Forensic состоит из отредактированных копий отдельных писем, для которых не удалось выполнить SPF, DKIM или и то, и другое. Эти отчеты не всегда доступны из-за проблем с конфиденциальностью, проблем с объемом или из-за того, что они не требуются для точного развертывания DMARC.
Когда вы хотите создавать отчеты о криминалистике?
Когда оба SPF и DKIM не работают.
При сбое SPF или DKIM.
Когда DKIM выходит из строя.
Когда SPF не работает.
Расслабленные или строгие механизмы?
Здесь вы можете изменить выравнивание идентификатора для каждого механизма.
Выравнивание идентификатора указывает, насколько строго оцениваются политики DKIM и SPF. Расслабленный режим позволяет доменам с проверкой подлинности SPF, которые используют общий домен организации с доменом отправителя электронного письма, пройти проверку DMARC.Строгий режим требует точного соответствия между доменом SPF и доменом отправителя электронного письма.
Выравнивание
по умолчанию
Расслабленный
Строгий
Хотите другую политику для поддоменов?
По умолчанию политика, примененная к example. com , будет применена к Department.example.com .
com , будет применена к Department.example.com .
Если вы не отправляете электронную почту с субдомена, установка политики отклонения субдомена поможет предотвратить злоупотребление электронной почтой в отношении субдоменов.
Если вы не уверены, идет ли ваша электронная почта от поддомена, выберите «Нет», пока не будут собраны дополнительные данные.
Какую политику вы хотите применить к электронной почте из поддомена этой записи DMARC, не прошедшей проверку DMARC? Это будет по умолчанию для любой политики, применяемой к домену организации.
К какому проценту писем вы хотите применить это?
DMARC позволяет пользователям постепенно наращивать свою политику, позволяя пользователям применять данную политику DMARC к определенному проценту потоков электронной почты. Если вы укажете процентное значение, отличное от 100, ваша политика DMARC будет применяться только к заданному проценту ваших сообщений.
Если вы укажете процентное значение, отличное от 100, ваша политика DMARC будет применяться только к заданному проценту ваших сообщений.
Начать создание
Предыдущий
Далее
Создать запись
Старт более
Тип записи
Цель
Хост
Расположение
Исправьте неверный домен, чтобы продолжить.
Исправьте неверные адреса, чтобы продолжить.
Исправьте недопустимый процент, чтобы продолжить.
Разрешите нашему приложению обрабатывать и визуализировать ваши данные DMARC таким образом, чтобы выявить пробелы в аутентификации (SPF / DKIM) и несанкционированное использование ваших доменов.
АДРЕС | |
| улица (индекс | )Jomfrustien 83 |
| Город | Драгёр |
| Штат / провинция / область | Область Sjælland |
| Почтовый индекс | 2791 |
| Телефон | 23-51-25340 |
| Временная почта | Это настоящий адрес электронной почты, нажмите здесь, чтобы получать письма. |
ФИНАНСЫ | |
| Тип кредитной карты | Visa Генератор кредитных карт |
| Номер кредитной карты | 4532115001441091 |
| CVV2 / CVV | 168 |
| Срок действия истекает | |
ЗАНЯТОСТЬ | |
| Промышленность | Архитектура и инженерное дело |
| Статус занятости | Работа на полную ставку |
| Месячная заработная плата | $ 7 800 |
| Компания | Музыка Камелот |
| Размер компании | 1-10 |
| Род занятий (должность) | Специалист по землеотводам |
БОЛЬШЕ | |
| Высота | 6 футов 1 дюйм (186 см) |
| Масса | 248. 4 фунта (112,9 кг) 4 фунта (112,9 кг) |
| Имя пользователя | lindsay.ok1980 Генератор имен пользователей |
| Пароль | aogh5eiSh6ph Генератор паролей |
| Контрольный вопрос | Какая девичья фамилия у вашей матери? |
| Ответ системы безопасности | Йона |
| Пользовательский агент браузера | Mozilla / 5.0 (Windows NT 10.0; WOW64; rv: 53.0) Gecko / 20100101 Firefox / 53.0 |
| Система | Окна 10 |
| GUID | 0453929d-484e-4cc3-af88-23724a84d0aa |
Что такое DMARC? Что нужно знать в 2021 году
Посмотрите короткометражный фильм
DMARC (Domain-based Message Authentication Reporting and Conformance) — это система проверки электронной почты, предназначенная для защиты домена электронной почты вашей компании от использования для подделки электронной почты, фишинга и других киберпреступлений. DMARC использует существующие методы аутентификации электронной почты SPF (Sender Policy Framework) DKIM (Domain Keys Identified Mail). DMARC добавляет важную функцию — отчетность. Когда владелец домена публикует запись DMARC в своей записи DNS, он получает представление о том, кто отправляет электронную почту от имени своего домена. Эта информация может быть использована для получения подробной информации о канале электронной почты. С помощью этой информации владелец домена может получить контроль над электронной почтой, отправляемой от его имени. Вы можете использовать DMARC для защиты своих доменов от фишинга или спуфинговых атак.
DMARC использует существующие методы аутентификации электронной почты SPF (Sender Policy Framework) DKIM (Domain Keys Identified Mail). DMARC добавляет важную функцию — отчетность. Когда владелец домена публикует запись DMARC в своей записи DNS, он получает представление о том, кто отправляет электронную почту от имени своего домена. Эта информация может быть использована для получения подробной информации о канале электронной почты. С помощью этой информации владелец домена может получить контроль над электронной почтой, отправляемой от его имени. Вы можете использовать DMARC для защиты своих доменов от фишинга или спуфинговых атак.
Как владелец веб-сайта вы хотите точно знать, что ваши посетители или клиенты будут видеть только электронные письма, которые вы отправили сами. Следовательно, DMARC необходим каждому владельцу домена. Защита вашей электронной почты с помощью DMARC дает получателям электронной почты уверенность в том, является ли электронное письмо законным и отправлено ли оно от вас. Это положительно сказывается на доставке электронной почты, а также предотвращает отправку электронной почты другими пользователями через ваш домен.
Это положительно сказывается на доставке электронной почты, а также предотвращает отправку электронной почты другими пользователями через ваш домен.
Индивидуальные услуги DMARC
Получите индивидуальный план и расценки.
Узнать цену
История DMARC
Стандарт DMARC был впервые опубликован в 2012 году для предотвращения злоупотреблений электронной почтой. Несколько лидеров отрасли работали вместе над созданием спецификации DMARC, DMARC был создан PayPal вместе с Google, Microsoft и Yahoo! Эти лидеры отрасли собрались вместе, чтобы разработать рабочую спецификацию, желая, чтобы она могла достичь официального статуса стандартов. Они создали стандарт DMARC на основе существующих методов аутентификации электронной почты SPF (Sender Policy Framework) и DKIM (Domain Keys Identified Mail).
DMARC изначально разрабатывался как протокол безопасности электронной почты. Сначала DMARC в основном использовался экспертами по безопасности в финансовой индустрии. С тех пор внедрение DMARC растет и становится все более распространенным в онлайн-ландшафте. На данный момент DMARC все больше и больше признается маркетологами электронной почты как аспект онлайн-безопасности и улучшенной доставляемости.
Сначала DMARC в основном использовался экспертами по безопасности в финансовой индустрии. С тех пор внедрение DMARC растет и становится все более распространенным в онлайн-ландшафте. На данный момент DMARC все больше и больше признается маркетологами электронной почты как аспект онлайн-безопасности и улучшенной доставляемости.
DMARC в настоящее время поддерживается всеми основными интернет-провайдерами (такими как Google, Microsoft, Yahoo! и т. Д.). В настоящее время DMARC ожидает утверждения, чтобы стать открытым стандартом, одобренным IETF (Internet Engineering Task Force).
Почему DMARC?
Имея почти 5 миллиардов учетных записей электронной почты по всему миру, нет канала с более широким охватом, чем канал электронной почты. Это гарантирует, что киберпреступники захотят использовать этот канал в злонамеренных целях. Несмотря на то, что в последние годы были приняты более строгие меры безопасности для защиты этого канала, преступность на этом канале растет с каждым годом. 95% всех хакерских атак и утечек данных связаны с электронной почтой.
95% всех хакерских атак и утечек данных связаны с электронной почтой.
Это область, в которой отчет об аутентификации на основе домена и соответствие требованиям (DMARC) добавляют ценность.DMARC не только обеспечивает полную информацию о каналах электронной почты, но и делает видимыми фишинговые атаки. DMARC более мощный: DMARC способен смягчать воздействие фишинга и атак вредоносного ПО, предотвращать спуфинг, защищать от злоупотреблений торговой маркой, мошенничества и предотвращать компрометацию корпоративной электронной почты. DMARC Analyzer позволяет организациям развернуть DMARC и упростить процесс развертывания DMARC.
Исследования показывают, почему DMARC имеет решающее значение:
- В среднем 1,6 миллиона долларов — это стоимость одной целевой фишинг-атаки для организаций
- С 2013 по 2016 годы убытки компаний приблизились к 1 доллару США.6 миллиардов
- Фишинговые атаки на 500 миллионов долларов ежегодно обманываются
- Обработка фишинговых атак обходится компании с 10 000 сотрудников в 3,7 миллиона долларов в год
- В среднем сотрудник тратит 4,16 часа в год на фишинговые атаки
- С января 2015 года по декабрь 2016 года долларовые показатели резко выросли на 2370% из-за фишинговых атак
- Только 3% всех пользователей сообщат своему руководству о фишинговых письмах
- Более 400 предприятий становятся жертвами мошенничества BEC каждый день
- 76% организаций сообщили, что они стали жертвами фишинг-атаки в 2016 году
- Организации, которые сообщили о том, что они стали жертвами фишинг-атаки в 2016 году: 76%
- Каждая третья компания стала жертвой мошенничества со стороны генерального директора по электронной почте
- 70% всех электронных писем в мире являются вредоносными
- Объем спам-писем в 2016 году увеличился в 4 раза
- Q3 2016 количество фишинговых писем с программами-вымогателями выросло до 97.
 25% по сравнению с 92% в 1 квартале 2016 г.
25% по сравнению с 92% в 1 квартале 2016 г. - В марте 2016 г. 9 из 10 фишинговых писем содержали вымогатели в той или иной форме
- Поддельные сообщения-фактуры — это фишинговая приманка №1
- Количество зарегистрированных фишинговых писем W-2 в 2017 году увеличилось на 870%
- 78% людей утверждают, что знают о рисках неизвестных ссылок в электронных письмах. И все равно нажимают
- В 2016 году ежемесячно в среднем наблюдалось более 400 000 фишинговых сайтов
- Открывается 30 процентов фишинговых писем
- В 2016 году 1 из 131 электронного письма содержало вредоносное ПО, это самый высокий показатель за 5 лет
DMARC Analyzer предоставляет удобное для пользователя программное обеспечение для анализа DMARC и выступает в качестве вашего экспертного руководства, чтобы как можно быстрее перейти к политике отклонения.
Где помогает DMARC?
Организациям и их клиентам наносят вред вредоносные электронные письма, рассылаемые от их имени, DMARC может блокировать эти атаки./yaootaweb-production-ng/media/crawledproductimages/6ccb3b9b01af5eee49a1aab5259b209fbe442e2a.jpg) С помощью DMARC организация может получить представление о своем канале электронной почты. Основываясь на этом понимании, организации могут работать над развертыванием и применением политики DMARC.
С помощью DMARC организация может получить представление о своем канале электронной почты. Основываясь на этом понимании, организации могут работать над развертыванием и применением политики DMARC.
Когда для политики DMARC задано значение p = reject, организации защищены от:
- Фишинг на клиентов организации
- Злоупотребление торговой маркой и мошенничество
- Атаки вредоносных программ и программ-вымогателей
- Случаи целевого фишинга и мошенничества со стороны генерального директора
С помощью анализатора DMARC Analyzer организации могут получить полное представление о своем канале электронной почты.Поскольку раньше организации могли получить информацию о фишинговых атаках только тогда, когда атака уже произошла, получение полного представления о канале электронной почты является большим преимуществом DMARC. С помощью DMARC становится возможным анализировать фишинговые атаки. Таким образом, клиенты могут быть проинформированы заранее и, следовательно, знать об этих атаках.
DMARC на практике
Основная цель DMARC — обнаруживать и предотвращать подделку электронной почты. Например, фишинговые мошенничества с использованием доменов банков для рассылки электронной почты от их имени.Клиенты из этого банка думают, что они получили правильное электронное письмо, что их банковская карта больше не действительна. Ссылка, по которой нужно щелкнуть, приведет к мошенническому веб-сайту. Этот веб-сайт точно такой же, как настоящий веб-сайт, и вход в систему предоставит киберпреступникам возможность использовать ваши учетные данные.
Изначально методы аутентификации электронной почты DKIM и SPF помогали защитить ваши домены от подобного мошенничества. Однако киберпреступники могут обойти эти меры безопасности. Чтобы полностью защитить ваш домен и канал электронной почты, DMARC создаст связь между SPF и DKIM.При внедрении DMARC в свою запись DNS вы получаете представление о своем канале электронной почты. Интернет-провайдеры будут предоставлять сводные (RUA) и криминалистические (RUF) отчеты DMARC на ежедневной основе, и эти отчеты можно отправлять на адрес электронной почты, опубликованный в вашей записи DMARC. Существует два доступных типа отчетов DMARC: сводные отчеты (RUA) и отчеты судебной экспертизы DMARC (RUF):
Существует два доступных типа отчетов DMARC: сводные отчеты (RUA) и отчеты судебной экспертизы DMARC (RUF):
Сводные отчеты DMARC (RUA)
- Отправляется ежедневно
- Предоставляет обзор трафика электронной почты
- Включает все IP-адреса, которые пытались передать электронную почту получателю, используя ваше доменное имя.
Пожалуйста, обратитесь к нашей статье об агрегированных отчетах DMARC для получения более подробной информации об агрегированных отчетах DMARC.
Криминалистические отчеты DMARC (RUF)
- Реальное время
- Отправляется только при сбоях
- Включает исходные заголовки сообщений
- Может включать исходное сообщение
Пожалуйста, обратитесь к нашей статье о криминалистических отчетах DMARC для получения более подробной информации о криминалистических отчетах DMARC.
DMARC Analyzer предоставляет панель инструментов для мониторинга и анализа результатов SPF, DKIM и DMARC. Публикации записи DMARC в вашем DNS недостаточно для защиты ваших доменов.DMARC работает с 3 политиками DMARC, это позволяет вам решать, что будет происходить с вашими электронными письмами. Политика «нет» предназначена только для сбора данных и мониторинга вашего текущего канала электронной почты. Чтобы обеспечить соблюдение вашего канала электронной почты, есть еще две политики DMARC. Политика «карантина» доставляет вредоносное электронное письмо в папку для спама получателя, а политика «отклонения» идет еще дальше, не доставляя это электронное письмо вообще.
Публикации записи DMARC в вашем DNS недостаточно для защиты ваших доменов.DMARC работает с 3 политиками DMARC, это позволяет вам решать, что будет происходить с вашими электронными письмами. Политика «нет» предназначена только для сбора данных и мониторинга вашего текущего канала электронной почты. Чтобы обеспечить соблюдение вашего канала электронной почты, есть еще две политики DMARC. Политика «карантина» доставляет вредоносное электронное письмо в папку для спама получателя, а политика «отклонения» идет еще дальше, не доставляя это электронное письмо вообще.
Таким образом, DMARC позволяет защитить ваши домены и позволить вам решить, что должно произойти, когда серверы от интернет-провайдера получают вредоносную электронную почту.Пожалуйста, обрати внимание! что DMARC — очень мощное решение для полной защиты вашего почтового домена при правильной настройке. Немедленный переход в карантин или отказ от политики может привести к множеству ложных срабатываний. Перед принудительным использованием ваших доменов убедитесь, что все настроено правильно. DMARC Analyzer предлагает 5 простых этапов, которые помогут вам защитить свой канал электронной почты.
DMARC Analyzer предлагает 5 простых этапов, которые помогут вам защитить свой канал электронной почты.
Снижайте влияние спуфинга с помощью DMARC
В DMARC можно указать получателям электронной почты, что делать с электронным письмом, которое не проходит проверку DMARC.В записи DMARC может быть определена политика DMARC, которая, в зависимости от настройки, указывает интернет-провайдеру, как обрабатывать электронные письма, не прошедшие проверки DMARC. Получатели электронной почты проверяют, имеют ли входящие сообщения допустимые записи SPF и DKIM и соответствуют ли они домену отправителя. После этих проверок сообщение можно рассматривать как DMARC-совместимое или как DMARC не удалось. После того, как получатель электронной почты проверит статус аутентификации сообщения, он будет обрабатывать сообщение по-разному в зависимости от установленной политики DMARC.
Доступны 3 возможные политики DMARC: Нет (только мониторинг), Карантин и Отклонить.
Политика монитора:
p = none Первая политика — это политика отсутствия (наблюдения): p = none . Политика DMARC none предписывает получателям электронной почты отправлять отчеты DMARC на адрес, опубликованный в тегах RUA или RUF записи DMARC. его называют политикой «Только мониторинг», потому что с помощью этой (рекомендованной для начала) политики вы получаете представление о своем канале электронной почты.Политика none дает представление о канале электронной почты, но не инструктирует получателей электронной почты обрабатывать электронные письма, не прошедшие проверку DMARC, по-другому, поэтому она также известна как политика мониторинга. Политика none дает представление только о том, кто отправляет электронную почту от имени домена, и не влияет на доставляемость.
Политика карантина:
p = карантин Вторая политика — это политика карантина: p = карантин . Помимо отправки отчетов DMARC, карантин политики DMARC предписывает получателям электронной почты помещать электронные письма, не прошедшие проверку DMARC, в папку спама получателя.Письма, прошедшие проверку DMARc, будут доставлены в основной почтовый ящик получателя. Политика карантина уже смягчит влияние спуфинга, но поддельные электронные письма по-прежнему будут доставляться получателю (в папку спама).
Политика карантина уже смягчит влияние спуфинга, но поддельные электронные письма по-прежнему будут доставляться получателю (в папку спама).
Политика отклонения:
p = отклонитьТретья политика — это политика отклонения: p = отклонить . Политика DMARC отклонена. Помимо отправки отчетов DMARC, отклонение политики DMARC предписывает получателям электронной почты вообще не доставлять электронные письма, не прошедшие проверки DMARC.Письма, прошедшие проверку DMARC, будут доставлены в основной почтовый ящик получателя. Эта политика снижает влияние спуфинга. Поскольку отклонение политики DMARC гарантирует, что все неверные установочные электронные письма (поддельные электронные письма) будут удалены получателем электронной почты, а не попадут во входящие сообщения получателя.
Политика DMARC — это запрос, а не обязательство.
Важно отметить, что политика DMARC предписывает обрабатывать электронные письма в соответствии с политикой DMARC, но получатели электронной почты не обязаны принимать во внимание политику DMARC. Получатели электронной почты иногда используют собственную локальную политику. Когда получатель электронной почты имеет разумные основания полагать, что электронное письмо является законным, он иногда применяет свою собственную локальную политику. Это означает, что электронное письмо, не прошедшее проверку DMARC, может попасть в основной почтовый ящик получателя, несмотря на то, что вы принудительно отклонили политику DMARC. Иногда получатели электронной почты заменяют политики DMARC локальной политикой.
Получатели электронной почты иногда используют собственную локальную политику. Когда получатель электронной почты имеет разумные основания полагать, что электронное письмо является законным, он иногда применяет свою собственную локальную политику. Это означает, что электронное письмо, не прошедшее проверку DMARC, может попасть в основной почтовый ящик получателя, несмотря на то, что вы принудительно отклонили политику DMARC. Иногда получатели электронной почты заменяют политики DMARC локальной политикой.
Посмотрите наше видео: объяснение DMARC
Недоразумения относительно DMARC
С помощью DMARC организация может блокировать вредоносные программы, фишинговые атаки и повышать доставляемость электронной почты! DMARC — мощный инструмент, но есть некоторые недопонимания по поводу DMARC:
.
DMARC не является средством быстрой доставки
Размещая запись DMARC (и применяя ее), интернет-провайдеры, внедрившие DMARC, заметят, что вы работаете над повышением безопасности своего канала электронной почты.Поэтому интернет-провайдеры, использующие DMARC, с большей вероятностью позволят вашим электронным письмам попадать в основной почтовый ящик получателя. Однако простое развертывание политики DMARC — это не просто быстрое решение проблемы доставки электронной почты. Развертывая и применяя политику DMARC, ваша доставляемость может улучшиться, однако это не является гарантией.
Немедленное применение политики отклонения — не лучшая идея
Когда организация сталкивается с фишинговой атакой от ее имени, она часто может немедленно заблокировать свой канал электронной почты, разместив запись DMARC и немедленно применив ее к политике 100% p = reject.Это действительно эффективно для немедленного блокирования фишинговых атак, однако это также приведет к потере законной электронной почты. Анализатор DMARC показывает, что в 99% случаев организации не достигают уровня соответствия почти 100%, когда они начинают использовать DMARC. DMARC Analyzer советует начать с политики p = none и отслеживать результаты, улучшать аутентификацию SPF и DKIM, а затем применять политику. В зависимости от инфраструктуры организации этот процесс может занять от одного до двенадцати месяцев.Мы настоятельно не рекомендуем немедленно применять политику отказа.
Анализатор DMARC показывает, что в 99% случаев организации не достигают уровня соответствия почти 100%, когда они начинают использовать DMARC. DMARC Analyzer советует начать с политики p = none и отслеживать результаты, улучшать аутентификацию SPF и DKIM, а затем применять политику. В зависимости от инфраструктуры организации этот процесс может занять от одного до двенадцати месяцев.Мы настоятельно не рекомендуем немедленно применять политику отказа.
DMARC не защищает потоки входящей электронной почты
DMARC не предназначен для защиты входящей части канала электронной почты, DMARC защищает исходящую часть канала электронной почты. Однако DMARC влияет на небольшую часть канала входящей электронной почты. На электронные письма, отправляемые коллегам, влияет DMARC. Поскольку это отправляемые электронные письма (хотя электронное письмо остается входящим), DMARC может влиять на эти электронные письма.
Удобное программное обеспечение для анализа DMARC
DMARC Analyzer предоставляет удобное для пользователя программное обеспечение для анализа DMARC и выступает в качестве вашего экспертного руководства, чтобы как можно быстрее перейти к политике отклонения. DMARC Analyzer предоставляет решение SaaS, которое позволяет организациям легко управлять сложным развертыванием DMARC. Решение обеспечивает 360-градусный обзор и управление по всем каналам электронной почты. Все сделано для того, чтобы сделать это как можно проще. Загрузите техническое описание для получения дополнительной информации о DMARC Analyzer
DMARC Analyzer предоставляет решение SaaS, которое позволяет организациям легко управлять сложным развертыванием DMARC. Решение обеспечивает 360-градусный обзор и управление по всем каналам электронной почты. Все сделано для того, чтобы сделать это как можно проще. Загрузите техническое описание для получения дополнительной информации о DMARC Analyzer
DMARC Analyzer предоставляет удобное для пользователя программное обеспечение для анализа DMARC и выступает в качестве вашего экспертного руководства, чтобы как можно быстрее перейти к политике отклонения.
• Дания: наиболее распространенные фамилии 2021 г.
• Дания: наиболее распространенные фамилии 2021 г. | StatistaДругая статистика по теме
Пожалуйста, создайте учетную запись сотрудника, чтобы иметь возможность отмечать статистику как избранную.
Затем вы можете получить доступ к своей любимой статистике через звездочку в заголовке.
Пожалуйста, авторизуйтесь, перейдя в «Моя учетная запись» → «Администрирование».После этого вы сможете отмечать статистику как избранную и использовать персональные статистические оповещения.
АутентифицироватьБазовая учетная запись
Познакомьтесь с платформой
У вас есть доступ только к базовой статистике.
Единая учетная запись
Идеальная учетная запись начального уровня для индивидуальных пользователей
- Мгновенный доступ к статистике за 1 мес
- Скачать в форматах XLS, PDF и PNG
- Подробные ссылки
$ 59 39 $ / месяц *
в первые 12 месяцев
Корпоративный аккаунт
Полный доступ
Корпоративное решение, включающее все функции.
* Цены не включают налог с продаж.
Узнайте больше о том, как Statista может поддержать ваш бизнес.
Статистическое управление Дании. (19 января 2021 г.). Самые распространенные фамилии в Дании по состоянию на январь 2021 г. [График]. В Statista. Получено 21 сентября 2021 г. с сайта https://www.statista.com/statistics/745971/most-common-surnames-in-denmark/
Statistics Denmark. «Самые распространенные фамилии в Дании по состоянию на январь 2021 года.»Диаграмма. 19 января 2021 года. Statista. По состоянию на 21 сентября 2021 года. Https://www.statista.com/statistics/745971/most-common-surnames-in-denmark/
Статистика Дании. (2021). Большинство общие фамилии в Дании по состоянию на январь 2021 г. Statista. Statista Inc .. Дата обращения: 21 сентября 2021 г. https://www.statista.com/statistics/745971/most-common-surnames-in-denmark/
Статистика Дании . «Самые распространенные фамилии в Дании по состоянию на январь 2021 г. » Statista, Statista Inc., 19 января 2021 г., https: // www.statista.com/statistics/745971/most-common-surnames-in-denmark/
» Statista, Statista Inc., 19 января 2021 г., https: // www.statista.com/statistics/745971/most-common-surnames-in-denmark/
Статистика Дании, Наиболее распространенные фамилии в Дании по состоянию на январь 2021 года Statista, https://www.statista.com/statistics/745971/most- common-surnames-in-denmark / (последнее посещение: 21 сентября 2021 г.)
Произошла ошибка при настройке вашего пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.

- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Гиппокамп как генератор последовательности
Trends Cogn Sci. Авторская рукопись; доступно в PMC 1 октября 2019 г.
Опубликован в окончательной отредактированной форме как:
PMCID: PMC6166479
NIHMSID: NIHMS1500443
Дьёрдь Бужаки
1 Институт неврологии, 3067, 435, Медицинский центр Лангоне, Нью-Йоркский университет, Нью-Йорк, Нью-Йорк 10016, США
2 Отделение неврологии, Медицинский центр Лангоне, Нью-Йоркский университет, Нью-Йорк, Нью-Йорк 10016, США
3 Центр неврологии, New York University, New York, NY 10003, USA
David Tingley
1 Neuroscience Institute, 435 East 30 th St. , Langone Medical Center, New York University, New York, NY 10016, USA
, Langone Medical Center, New York University, New York, NY 10016, USA
1 Институт неврологии, 435 East 30 th St., Медицинский центр Лангоне, Нью-Йоркский университет, Нью-Йорк, Нью-Йорк 10016, США
2 Отделение неврологии, Медицинский центр Лангоне, Нью-Йоркский университет, Нью-Йорк, Нью-Йорк 10016, США
3 Центр неврологии, Нью-Йоркский университет, Нью-Йорк, штат Нью-Йорк 10003, США
4 Контактное лицо
Окончательная отредактированная версия этой статьи доступна на сайте Trends Cogn Sci. См. Другие статьи в PMC, в которых цитируется опубликованная статья.Abstract
Нейронные вычисления часто сравнивают с измеренным прибором расстоянием или продолжительностью, и такие отношения интерпретируются человеком-наблюдателем.Однако нейронные цепи не зависят от инструментов, созданных руками человека, но выполняют вычисления относительно внутренней скорости изменения. В то время как нейронные корреляции с внешними показателями, такими как расстояние или продолжительность, можно наблюдать в скорости всплесков или других показателях нейронной активности, для мозга важно то, как такие паттерны активности используются нижележащими нейронными наблюдателями. Мы предполагаем, что операции гиппокампа можно описать последовательной активностью нейронных сборок и их внутренне определенной скоростью изменения, не прибегая к концепции пространства или времени.
Мы предполагаем, что операции гиппокампа можно описать последовательной активностью нейронных сборок и их внутренне определенной скоростью изменения, не прибегая к концепции пространства или времени.
Ключевые слова: Клетки места, временные клетки, тета-колебания, фазовое кодирование, латеральная перегородка
Пространство, время и память в системе гиппокампа
Возможно, наиболее важным вычислением, приписываемым гиппокампальной системе, является эпизодическая память [1] . Эпизодическая память относится к важному понятию, которое описывает события, пережитые от первого лица, в контексте пространства и времени («что случилось со мной, где и когда?» [2]. Как такие личные воспоминания создаются и воссоздаются, является предметом интенсивных исследований. .Если бы мозг должен был хранить отдельное представление каждого индивидуального опыта нашей жизни, то есть каждую комбинацию «что», «где» и «когда», список был бы чрезвычайно длинным и потребовал бы чрезвычайно большой емкости памяти. Вызов эпизода из такого длинного списка был бы сложным и трудоемким. Вымышленный персонаж Борхеса, Фунес Мемориус, обладал безупречной памятью и мог вспомнить каждый момент своей деятельности в предыдущий день, но на это ему потребовался еще целый день [3].Альтернативным решением является хранение компонентов «что», «где» и «когда» отдельно и воссоздание исходного эпизода путем повторного встраивания «что» в порядковую структуру «где» и «когда». Такое разделение компонентов памяти очень приятно. Допуская независимые пространственные и временные рамки и добавляя их к кодированию «чего», может показаться, что нейробиология определила дорожную карту для раскрытия нейрофизиологических механизмов эпизодической памяти и понимания глобальной функции системы гиппокампа (вставка 1).
Вызов эпизода из такого длинного списка был бы сложным и трудоемким. Вымышленный персонаж Борхеса, Фунес Мемориус, обладал безупречной памятью и мог вспомнить каждый момент своей деятельности в предыдущий день, но на это ему потребовался еще целый день [3].Альтернативным решением является хранение компонентов «что», «где» и «когда» отдельно и воссоздание исходного эпизода путем повторного встраивания «что» в порядковую структуру «где» и «когда». Такое разделение компонентов памяти очень приятно. Допуская независимые пространственные и временные рамки и добавляя их к кодированию «чего», может показаться, что нейробиология определила дорожную карту для раскрытия нейрофизиологических механизмов эпизодической памяти и понимания глобальной функции системы гиппокампа (вставка 1).
Вставка 1.
Эволюция взглядов на пространство и время в гиппокампе
Два ключевых эпизода предоставили критическую информацию о работе гиппокампа и связанных с ним структур: открытие амнезии у пациента H. M. после двустороннего удаления гиппокампа [1] и открытия клеток места у крыс [14]. Тем не менее, как эти наблюдения соотносятся друг с другом и со многими другими функциями, приписываемыми гиппокампу, остается загадкой [5]. Существует общее мнение, что гиппокамп необходим для использования пространственных отношений окружающей среды и для способности запоминать конкретные личные события [2].Эпизодическая память определяется как пережитые от первого лица события в контексте пространства и времени («что случилось со мной, где и когда?»; [2]). Основываясь на исследованиях на животных, O’Keefe & Nadel (1978) концептуализировали когнитивную карту гиппокампа и предположили, что «гиппокамп является ядром системы нейронной памяти, обеспечивающей объективную пространственную структуру, в которой расположены элементы и события опыта организма и взаимосвязаны ». Таким образом, когнитивная карта может обеспечить необходимую пространственную основу для эпизодов.Недавно Говард Эйхенбаум утверждал, что структура эпизодической памяти также нуждается во временном контексте, и назвал гиппокампальные и энторинальные нейроны «временными клетками» [21,74,126,127]; «Временные ячейки могут играть роль в эпизодической памяти, отмечая события, происходящие во времени, точно так же, как ячейки места отображают места, где события происходят в пространстве».
M. после двустороннего удаления гиппокампа [1] и открытия клеток места у крыс [14]. Тем не менее, как эти наблюдения соотносятся друг с другом и со многими другими функциями, приписываемыми гиппокампу, остается загадкой [5]. Существует общее мнение, что гиппокамп необходим для использования пространственных отношений окружающей среды и для способности запоминать конкретные личные события [2].Эпизодическая память определяется как пережитые от первого лица события в контексте пространства и времени («что случилось со мной, где и когда?»; [2]). Основываясь на исследованиях на животных, O’Keefe & Nadel (1978) концептуализировали когнитивную карту гиппокампа и предположили, что «гиппокамп является ядром системы нейронной памяти, обеспечивающей объективную пространственную структуру, в которой расположены элементы и события опыта организма и взаимосвязаны ». Таким образом, когнитивная карта может обеспечить необходимую пространственную основу для эпизодов.Недавно Говард Эйхенбаум утверждал, что структура эпизодической памяти также нуждается во временном контексте, и назвал гиппокампальные и энторинальные нейроны «временными клетками» [21,74,126,127]; «Временные ячейки могут играть роль в эпизодической памяти, отмечая события, происходящие во времени, точно так же, как ячейки места отображают места, где события происходят в пространстве».gz100vkozhuhe.jpg) [5] Согласно этой структуре, фундаментальная цель нейробиологии — раскрыть нейронные «репрезентации» пространства и времени, в которые могут быть встроены вещи и события.
[5] Согласно этой структуре, фундаментальная цель нейробиологии — раскрыть нейронные «репрезентации» пространства и времени, в которые могут быть встроены вещи и события.
В этом обзоре мы ставим под сомнение эту общую схему как на концептуальной, так и на экспериментальной основе.Современные концептуальные рамки нейробиологии основаны на идеях классической физики о пространстве и времени. Однако в современной физике «больше нет пространства,« содержащего »мир, и нет времени,« в котором »происходят события» [4]. Мы предполагаем, что нейробиология требует аналогичного обновления парадигмы. Когда концепции пространства и времени подвергаются тщательному изучению, они оказываются просто придуманными людьми терминами, удобными для классификации событий мира, а не независимыми сущностями. На экспериментальном уровне мы считаем, что концептуализация системы гиппокампа как устройства, вычисляющего пространство и время, не учитывает многие экспериментальные наблюдения [5], потому что гиппокамп может быть «слепым» в отношении модальности своих входов. Какая бы информация ни была передана ему из каких бы то ни было частей неокортекса, он активирует одни и те же вычислительные алгоритмы. Таким образом, конкретные термины, которые мы склонны относить к обычным вычислениям гиппокампа, могут в значительной степени отражать экспериментальные условия и задействованные неокортикальные входы, а не внутреннее вычисление пространства или времени.
Какая бы информация ни была передана ему из каких бы то ни было частей неокортекса, он активирует одни и те же вычислительные алгоритмы. Таким образом, конкретные термины, которые мы склонны относить к обычным вычислениям гиппокампа, могут в значительной степени отражать экспериментальные условия и задействованные неокортикальные входы, а не внутреннее вычисление пространства или времени.
Пространство и время как организующие координаты по сравнению с пространством-временем в физике
Концепции пространства и времени можно рассматривать как аксиомы вселенной, независимые друг от друга и всего остального.Они также являются фундаментальными организаторами наших идей [6]. Их привлекательность восходит к ньютоновской структуре, в которой события происходят в большом «театре» или «контейнере» и разворачиваются на временной шкале. Однако концепции пространства и времени безразмерны и неизмеримы, поэтому их нельзя изучать напрямую. Современная наука преобразовала эти абстрактные концепции, заменив их измеримыми вариантами: расстояние и продолжительность . Постоянно растущее распространение и точность измерительных инструментов изменили человеческое представление о пространстве и времени.В сегодняшнем промышленно развитом мире было бы трудно представить и организовать человеческую жизнь без часов, хотя несколько закрытых человеческих обществ дожили до наших дней без развития лингвистических структур, связанных со временем. Тем не менее, эти люди понимают последовательность событий [7].
Постоянно растущее распространение и точность измерительных инструментов изменили человеческое представление о пространстве и времени.В сегодняшнем промышленно развитом мире было бы трудно представить и организовать человеческую жизнь без часов, хотя несколько закрытых человеческих обществ дожили до наших дней без развития лингвистических структур, связанных со временем. Тем не менее, эти люди понимают последовательность событий [7].
Предполагаемая независимость пространства и времени обсуждалась как лингвистами, так и физиками. В наших повседневных разговорах эти размеры часто используются как синонимы: «Ресторан находится в пяти минутах ходьбы отсюда».Единица расстояния определяется временем; световая секунда — это расстояние, которое свет проходит за одну секунду. Долготы Земли (то есть расстояния) известны как часовые пояса. Сегодня мы ориентируемся с помощью GPS, в котором нет метрики. Он вычисляет положение, определяя временной интервал (разность фаз), который требуется сигналам, чтобы достичь приемника от нескольких спутников.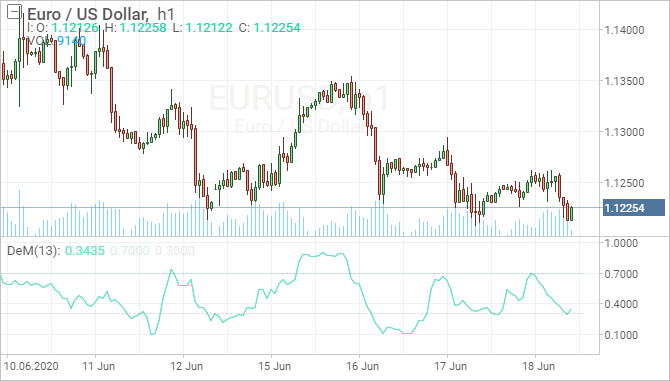
Даже в классической механике эта независимость пространства и времени начала подвергаться сомнению. Если время — это среда, через которую перемещаются вещи, определение времени требует, чтобы что-то перемещалось в пространстве.Таким образом, время записывается материей. В 20-м веке классические «контейнерные» перспективы пространства и времени были заменены пространственно-временной моделью общей теории относительности, в которой ось времени представляет собой четвертое измерение. В общей теории относительности прошлое и будущее полностью симметричны. Понятие «сейчас» не имеет значения.
Эти дискуссии внутри физики и между физикой и философией [8] мало повлияли на нейробиологию. Исследования пространства и времени в мозге продолжились в традициях классической физики, где пространство и время сохраняют независимость друг от друга и всего остального.
Концептуальные вопросы
Постоянное изменение активности — фундаментальное правило мозга. В своей исследовательской практике мы сравниваем эволюцию паттернов возбуждения нейронов, последовательностей сборки клеток или других показателей с единицами инструментов и часто находим надежные корреляции. Таким образом, экспериментатор интерпретирует как значение нейронных ответов, измеренных с помощью инструментов, так и значение единиц, измеренных этими инструментами, без независимого обоснования.Это сравнение создает иллюзию того, что мозг вычисляет или «представляет» время, но то, что мы называем временем, — это просто набор правил, управляющих изменениями. Эта проблематичная практика — интерпретация как значения ответов мозга, измеренных с помощью инструментов, так и значения единиц этих инструментов — может быть фундаментальной ошибкой в нашем текущем экспериментальном подходе. Наблюдатель не найдет смысла в ответах за пределами переменных, измеряемых приборами. Если расстояние и продолжительность определены и поняты с помощью созданных человеком измерительных приборов, можно задаться вопросом, как пространство и время вычисляются и понимаются другими животными, которые не могут читать эти инструменты [9].
Таким образом, экспериментатор интерпретирует как значение нейронных ответов, измеренных с помощью инструментов, так и значение единиц, измеренных этими инструментами, без независимого обоснования.Это сравнение создает иллюзию того, что мозг вычисляет или «представляет» время, но то, что мы называем временем, — это просто набор правил, управляющих изменениями. Эта проблематичная практика — интерпретация как значения ответов мозга, измеренных с помощью инструментов, так и значения единиц этих инструментов — может быть фундаментальной ошибкой в нашем текущем экспериментальном подходе. Наблюдатель не найдет смысла в ответах за пределами переменных, измеряемых приборами. Если расстояние и продолжительность определены и поняты с помощью созданных человеком измерительных приборов, можно задаться вопросом, как пространство и время вычисляются и понимаются другими животными, которые не могут читать эти инструменты [9].
Мы должны осознавать разницу между пространственными и временными свойствами процесса представления чего-либо и представлениями пространства и времени. Даже если нейронная активность надежно коррелирует с пространственно-временной последовательностью событий («временная последовательность представлений»), такие корреляции не обязательно означают, что нейронная активность вычисляет продолжительность («представление временных последовательностей»; [10-12]). Другими словами, мы не должны отождествлять описание событий с их субъективной интерпретацией.Ни инструменты, ни мозг не создают пространство или время. Время ничего не значит для часов, а тиканье не имеет внутреннего значения без наблюдателя. Поскольку люди определили единицы измерения стержней и часов, этот процесс неизбежно определяет наши представления о пространстве и времени.
Даже если нейронная активность надежно коррелирует с пространственно-временной последовательностью событий («временная последовательность представлений»), такие корреляции не обязательно означают, что нейронная активность вычисляет продолжительность («представление временных последовательностей»; [10-12]). Другими словами, мы не должны отождествлять описание событий с их субъективной интерпретацией.Ни инструменты, ни мозг не создают пространство или время. Время ничего не значит для часов, а тиканье не имеет внутреннего значения без наблюдателя. Поскольку люди определили единицы измерения стержней и часов, этот процесс неизбежно определяет наши представления о пространстве и времени.
Классическая основанная на физике структура породила две в значительной степени независимые области нейробиологии, посвященные проблемам пространства и времени с отдельной литературой [9,13]. Можно возразить, что законы физики неприменимы к «психологическому» времени.Оживленное пространство и время могут фундаментально отличаться от микроскопических и астрономических миров, управляемых теорией относительности и квантовой физикой. В качестве альтернативы можно принять позицию, согласно которой разрешение отношений между пространством и временем является задачей нейробиологии: что означают пространство и время для мозга? Возможно, хорошее место для начала — обсуждение нейронных механизмов гиппокампа и его союзников.
В качестве альтернативы можно принять позицию, согласно которой разрешение отношений между пространством и временем является задачей нейробиологии: что означают пространство и время для мозга? Возможно, хорошее место для начала — обсуждение нейронных механизмов гиппокампа и его союзников.
Анатомическая организация гиппокампа делает его слепым к модальностям
Было предложено, чтобы гиппокамп и родственные ему структуры «кодировали» пространственную карту [14,15], а в последнее время «представляли» время [16,17 ].Эти теории приводят к постулатам клеток места в гиппокампе [18], ячеек сетки в энторинальной коре [19] и временных клеток в обеих структурах [20,21], очевидно, прокладывая путь к пониманию нейронные механизмы эпизодической памяти [2,16,17]. Чтобы критически оценить эту структуру, поучительно осмотреть анатомические субстраты, которые, как считается, «представляют» пространство и время.
Гиппокампально-энторинальная система имеет топографически организованную двунаправленную связь с большим неокортексом. У грызунов основная часть неокортекса обрабатывает сенсорные входы и генерирует моторные выходы. Напротив, у приматов большая часть неокортекса предназначена для вычисления более сложных функций (). Следовательно, несенсорные входы в гиппокамп прогрессивно увеличиваются параллельно с увеличением неокортекса во время эволюции млекопитающих. Перегородочная (дорсальная) и средневисочная трети гиппокампа грызунов получают зрительно-пространственные и другие сенсорные сигналы от дорсальной энторинальной коры, тогда как височный полюс сообщается в основном с гипоталамической, миндалевидной и префронтальной областями [22,23].
У грызунов основная часть неокортекса обрабатывает сенсорные входы и генерирует моторные выходы. Напротив, у приматов большая часть неокортекса предназначена для вычисления более сложных функций (). Следовательно, несенсорные входы в гиппокамп прогрессивно увеличиваются параллельно с увеличением неокортекса во время эволюции млекопитающих. Перегородочная (дорсальная) и средневисочная трети гиппокампа грызунов получают зрительно-пространственные и другие сенсорные сигналы от дорсальной энторинальной коры, тогда как височный полюс сообщается в основном с гипоталамической, миндалевидной и префронтальной областями [22,23].
Вентральный квадрант гиппокампа грызунов у приматов непропорционально увеличен, чтобы не отставать от все большей доли неокортекса более высокого порядка. Только относительно небольшая хвостовая часть гиппокампа приматов сообщается с зрительно-пространственными областями. Этот хвост гомологичен спинному промежуточному гиппокампу грызунов. Показаны различные связи видов с сегментами перегородочно-височной оси и от них.Большинство записей и манипуляций с мозгом грызунов производились в спинном гиппокампе. Адаптировано из [29], Изображение: Debbie Maizels Springer Nature.
Показаны различные связи видов с сегментами перегородочно-височной оси и от них.Большинство записей и манипуляций с мозгом грызунов производились в спинном гиппокампе. Адаптировано из [29], Изображение: Debbie Maizels Springer Nature.
В головном мозге приматов вентральный полюс формирует ункус и тело и становится непропорционально увеличенным, чтобы не отставать от большой доли не родственного неокортекса. Только относительно небольшая часть хвоста гиппокампа приматов сообщается с зрительно-пространственными областями, и этот хвост гомологичен спинному гиппокампу грызунов.Напротив, оставшаяся основная часть гиппокампа приматов получает и отправляет нейронные сигналы в не-родственный (также называемый более высоким или ассоциативным) неокортекс. У людей также были продемонстрированы различия в функциях левого и правого гиппокампа, соизмеримые с полушарной специализацией неокортекса [24]. Если взять за основу анатомию, сенсорные «репрезентации» могут быть лишь неполной работой для гиппокампа. Поскольку в мозгу нет датчиков ни пространства, ни времени, неясно, как расстояние и продолжительность измеряются мозгом и «моделируются» системой гиппокампа.
Поскольку в мозгу нет датчиков ни пространства, ни времени, неясно, как расстояние и продолжительность измеряются мозгом и «моделируются» системой гиппокампа.
В отличие от неокортекса, организованного по модульному принципу, гиппокамп можно рассматривать как единый гигантский модуль с чрезвычайно большой повторяющейся системой возбуждения [25,26], даже если принять во внимание относительно постепенные анатомические, физиологические и молекулярные изменения от перегородки к перегородке. височный полюс [22,27–29]. Сильно связанный граф внутригиппокампального контура служит для объединения и смешивания нейронных сообщений из обширных областей неокортекса, независимо от их модальностей и происхождения.Эта структурная организация и скоординированные тета-осцилляции во всей гиппокампально-энторинальной системе [30–32] предполагают, что одни и те же общие вычисления выполняются для всех входящих сигналов в гиппокамп, в значительной степени независимо от их неокортикального источника. То, подразумевает ли конкретное экспериментальное наблюдение, что гиппокамп вычисляет пространство, время, память, планирование, абстрактные концепции или другие отношения [33–35], может зависеть от плана эксперимента и коркового сигнала, а не от вычислений гиппокампа как таковых.Это безразмерное реляционное вычисление в гиппокампе, возможно, не так уж сильно отличается от идеи Говарда Эйхенбаума о «представлении многомерных отношений между событиями в пространстве памяти» [17,36,37], хотя вполне может быть, что «многомерность» предопределена через анатомия и связь (). Однако, даже если мы предположим, что эти кажущиеся дискретными представления рассчитываются выделенными нейронами и цепями, необходимо решить, могут ли эти якобы отдельные информационные каналы по-разному интерпретироваться нижестоящими механизмами считывания.
То, подразумевает ли конкретное экспериментальное наблюдение, что гиппокамп вычисляет пространство, время, память, планирование, абстрактные концепции или другие отношения [33–35], может зависеть от плана эксперимента и коркового сигнала, а не от вычислений гиппокампа как таковых.Это безразмерное реляционное вычисление в гиппокампе, возможно, не так уж сильно отличается от идеи Говарда Эйхенбаума о «представлении многомерных отношений между событиями в пространстве памяти» [17,36,37], хотя вполне может быть, что «многомерность» предопределена через анатомия и связь (). Однако, даже если мы предположим, что эти кажущиеся дискретными представления рассчитываются выделенными нейронами и цепями, необходимо решить, могут ли эти якобы отдельные информационные каналы по-разному интерпретироваться нижестоящими механизмами считывания.
Эквивалентность расстояния и продолжительности в гиппокампе
Классическая физика уже показала, что расстояние и продолжительность связаны через скорость: знание любых двух переменных может идентифицировать третью, поэтому одна всегда избыточна. Давайте исследуем их отношения в гиппокампе.
Давайте исследуем их отношения в гиппокампе.
Местные поля нейронов гиппокампа могут быть относительно большими, от 20 до 300 сантиметров по сравнению с типичным размером тела ~ 20 см у крыс. Таким образом, поля мест от нескольких нейронов могут перекрываться друг с другом в течение нескольких тета-циклов (; [38–41]).Во время каждого тета-цикла происходит примерно семь циклов гамма , и внутри каждого из них находится клеточная сборка, чья активность коррелирует с данным пространственным положением [42–45]. Пиковая последовательность нейронных сборок в каждом тета-цикле предсказывает последовательность прошлых и будущих местоположений на пути крысы. По мере движения крысы новые клетки места присоединяются к существующей сборке в тета-цикле, в то время как шипы клеток места, поле которых только что покинуло животное, исчезают. Нейроны с предстоящими полями мест срабатывают на поздней фазе тета-цикла, тогда как нейроны с недавно пройденными полями мест действуют на ранних фазах. Таким образом, по мере того, как животное движется вперед, положение последовательности конкретного нейрона перемещается из восходящей (поздней) фазы в нисходящую (раннюю) фазу тета-цикла (известную как «фазовая прецессия»; [46];). Такое смещение членства «один-за-один раз» поддерживает относительно постоянным количество клеточных сборок в пределах тета-циклов [39]. В более крупных средах пространственные поля становятся больше, как и их расстояния друг от друга [40,47,48]. Следовательно, коэффициент «сжатие» расстояние-длительность больше в больших средах, а пространственное разрешение хуже.Другими словами, гиппокамп, таким образом, может «уменьшать» или «увеличивать» в зависимости от размера окружающей среды [40,41]. Скорее, чем исключение, эта форма упорядоченной активности, по-видимому, является общим принципом организации в областях мозга, которые, как считается, несут различные типы информации [49–52].
Таким образом, по мере того, как животное движется вперед, положение последовательности конкретного нейрона перемещается из восходящей (поздней) фазы в нисходящую (раннюю) фазу тета-цикла (известную как «фазовая прецессия»; [46];). Такое смещение членства «один-за-один раз» поддерживает относительно постоянным количество клеточных сборок в пределах тета-циклов [39]. В более крупных средах пространственные поля становятся больше, как и их расстояния друг от друга [40,47,48]. Следовательно, коэффициент «сжатие» расстояние-длительность больше в больших средах, а пространственное разрешение хуже.Другими словами, гиппокамп, таким образом, может «уменьшать» или «увеличивать» в зависимости от размера окружающей среды [40,41]. Скорее, чем исключение, эта форма упорядоченной активности, по-видимому, является общим принципом организации в областях мозга, которые, как считается, несут различные типы информации [49–52].
A. Пиковая активность двухместных клеток и тета-ритм потенциала местного поля во время ходьбы по лабиринту. Временная длительность T — это время, необходимое крысе, чтобы пробежать расстояние между пиками двух полей мест (шкала поведенческого времени).τ — временной сдвиг между двумя нейронами в тета-цикле («тета-шкала времени»). B. Идеализированные перекрывающиеся поля мест трех ячеек с идентичной частотой тета-колебаний, иллюстрирующие взаимосвязь между T и τ. Связь между расстояниями пространственных полей и временными сдвигами (τ) показывает линейную зависимость в пределах шкалы тета-времени (100–160 мс). C. Траектории движения крысы по полю места в двух испытаниях с разными скоростями. Д. Шипы одной ячейки места и соответствующий тета-ритм тех же двух испытаний, что и в C. Горизонтальные двойные стрелки указывают время, которое потребовалось крысе, чтобы пробежать через поле места. E. Количество всплесков в поле места нейрона аналогично тестам на медленном и быстром беге.
Пиковая активность двухместных клеток и тета-ритм потенциала местного поля во время ходьбы по лабиринту. Временная длительность T — это время, необходимое крысе, чтобы пробежать расстояние между пиками двух полей мест (шкала поведенческого времени).τ — временной сдвиг между двумя нейронами в тета-цикле («тета-шкала времени»). B. Идеализированные перекрывающиеся поля мест трех ячеек с идентичной частотой тета-колебаний, иллюстрирующие взаимосвязь между T и τ. Связь между расстояниями пространственных полей и временными сдвигами (τ) показывает линейную зависимость в пределах шкалы тета-времени (100–160 мс). C. Траектории движения крысы по полю места в двух испытаниях с разными скоростями. Д. Шипы одной ячейки места и соответствующий тета-ритм тех же двух испытаний, что и в C. Горизонтальные двойные стрелки указывают время, которое потребовалось крысе, чтобы пробежать через поле места. E. Количество всплесков в поле места нейрона аналогично тестам на медленном и быстром беге. Испытания отсортированы по скорости от самых медленных до самых быстрых. F. В дополнение к текущему положению, начальная и конечная позиции также могут быть считаны из тета-фазы спайка.Слева, декодированная вероятность (высокая вероятность: красный) того, что крыса занимает каждую позицию трека (ось y), вычисляется в каждой фазе тета (ось x, белая синусоида). На каждом подзаголовке диапазон белой синусоидальной волны обозначает фактическое положение крысы. Как правило, велика вероятность того, что крыса займет свое реальное положение. Последовательность тета может быть визуализирована диагональными полосами с высокой вероятностью, которые начинаются в позиции НАЧАЛО в фазе падения тета и заканчиваются в позиции КОНЕЦ в фазе роста.Правильно, одни и те же данные усреднены по всем позициям, которые фактически занимала крыса. Обратите внимание, что тета-последовательности заполнены представлениями линейных позиций НАЧАЛА и КОНЕЦ дорожки в фазах спада и подъема, соответственно.
Испытания отсортированы по скорости от самых медленных до самых быстрых. F. В дополнение к текущему положению, начальная и конечная позиции также могут быть считаны из тета-фазы спайка.Слева, декодированная вероятность (высокая вероятность: красный) того, что крыса занимает каждую позицию трека (ось y), вычисляется в каждой фазе тета (ось x, белая синусоида). На каждом подзаголовке диапазон белой синусоидальной волны обозначает фактическое положение крысы. Как правило, велика вероятность того, что крыса займет свое реальное положение. Последовательность тета может быть визуализирована диагональными полосами с высокой вероятностью, которые начинаются в позиции НАЧАЛО в фазе падения тета и заканчиваются в позиции КОНЕЦ в фазе роста.Правильно, одни и те же данные усреднены по всем позициям, которые фактически занимала крыса. Обратите внимание, что тета-последовательности заполнены представлениями линейных позиций НАЧАЛА и КОНЕЦ дорожки в фазах спада и подъема, соответственно. G. Сборки клеток места в гиппокампе организованы тета-колебаниями («время гиппокампа»). Каждый ряд точек — это проба пиковой активности десятизначных ячеек (обозначенных разными цветами). Верхняя панель: Испытания показаны с привязкой к расстоянию через лабиринт. Средняя панель: Испытания показаны в зависимости от времени, прошедшего с начала. Нижняя панель: Испытания показаны как синхронизированная по фазе активность нейронов в последовательных тета-циклах. A-E, после [58]; F, по [128]; G, любезно предоставлено Кариной Курто и Евой Пасталковой.
G. Сборки клеток места в гиппокампе организованы тета-колебаниями («время гиппокампа»). Каждый ряд точек — это проба пиковой активности десятизначных ячеек (обозначенных разными цветами). Верхняя панель: Испытания показаны с привязкой к расстоянию через лабиринт. Средняя панель: Испытания показаны в зависимости от времени, прошедшего с начала. Нижняя панель: Испытания показаны как синхронизированная по фазе активность нейронов в последовательных тета-циклах. A-E, после [58]; F, по [128]; G, любезно предоставлено Кариной Курто и Евой Пасталковой.
Следовательно, из наблюдаемых экспериментатором корреляций между положениями в окружающей среде и данными фазы спайк-тета, можно сделать вывод, что гиппокамп измеряет расстояния [47]. Однако одних лишь внешних стимулов недостаточно для обеспечения метрики, необходимой для пространственной карты.Для калибровки расстояний животное должно двигаться, чтобы понять отношения между ориентирами [53]. В отсутствие движения поля мест не появляются или становятся очень большими [54,55].
В отсутствие движения поля мест не появляются или становятся очень большими [54,55].
В отличие от абстрактного пространства и времени, мозг может непосредственно определять скорость и направление головы и комбинировать их для определения скорости [53,56,57]. Сигнал скорости может быть передан в гиппокамп от вестибулярной системы, мышечных афферентов или оптического потока [53], а пройденное расстояние можно рассчитать как произведение количества циклов тета-колебаний (вместо продолжительности) и скорости усиление [58].
Доступ к информации о скорости движения критически важен для поддержания надежной взаимосвязи между фазами всплеска ячеек места относительно предстоящих и предыдущих позиций. Когда крыса пересекает поле места нейрона за одну секунду в одном испытании, а затем за полсекунды во время другого испытания, клетка места будет активна в течение 8 и 4 тета-циклов соответственно (при условии тета-частоты 8 Гц;). Количество шипов в поле места меняется относительно мало, даже если скорость крысы меняется. По этой причине количество всплесков на тета-волну удваивается. Из-за этого увеличения скорости возбуждения, которое отражает более сильное возбуждение нейрона по мере увеличения скорости, величина сдвига тета-фазы от цикла к циклу увеличивается пропорционально. В результате, усиление скорости компенсирует более короткое время, проведенное в поле места, оставляя связь между фазой выброса и пространственным положением относительно неизменной (; [41,58]). Скоростная чувствительность нейронов гиппокампа снижается от перегородки к височному полюсу, и в результате поля места соответственно становятся больше [29,59,60].
По этой причине количество всплесков на тета-волну удваивается. Из-за этого увеличения скорости возбуждения, которое отражает более сильное возбуждение нейрона по мере увеличения скорости, величина сдвига тета-фазы от цикла к циклу увеличивается пропорционально. В результате, усиление скорости компенсирует более короткое время, проведенное в поле места, оставляя связь между фазой выброса и пространственным положением относительно неизменной (; [41,58]). Скоростная чувствительность нейронов гиппокампа снижается от перегородки к височному полюсу, и в результате поля места соответственно становятся больше [29,59,60].
Таким образом, как и в физике, мозг способен уравнивать расстояния и длительности посредством модуляции, зависящей от скорости [12,61]. Вычисление времени, измеряемое отдельными нейронами или сборками, может быть динамически деформировано относительно времени часов. Последующее сжатие или декомпрессия может быть вызвано их динамически изменяющейся и зависимой от состояния чувствительностью к сигналам скорости. В отсутствие движения механизмы, лежащие в основе субъективного термина «внимание», могут действовать как усиление и усиление критических аспектов закодированного или вызванного содержания мысленного путешествия.Таким образом, можно предположить, что внимание — это интернализованная мозгом версия скорости, которая может влиять на скорость изменения в последовательно организованной деятельности [9,62].
В отсутствие движения механизмы, лежащие в основе субъективного термина «внимание», могут действовать как усиление и усиление критических аспектов закодированного или вызванного содержания мысленного путешествия.Таким образом, можно предположить, что внимание — это интернализованная мозгом версия скорости, которая может влиять на скорость изменения в последовательно организованной деятельности [9,62].
Гиппокамп как генератор последовательности
Какое значение имеет постулированная выше эквивалентность пространства-времени для мозга? Могут ли пространство и время взять на себя роль друг друга? Ниже мы выдвигаем гипотезу о том, что основная функция гиппокампа — генерировать постоянно меняющиеся последовательности без необходимости ссылаться на концепции пространства или времени.
Внешне навязанные и генерируемые внутри последовательности сборки клеток
Обширные экспериментальные данные показывают, что нейроны гиппокампа могут реагировать на ориентиры [47,63] и сигналы от тела [53], что приводит к молчаливому предположению, что последовательности клеток места вызваны изменения внешних входов [63,64] по мере того, как животное исследует окружающую среду.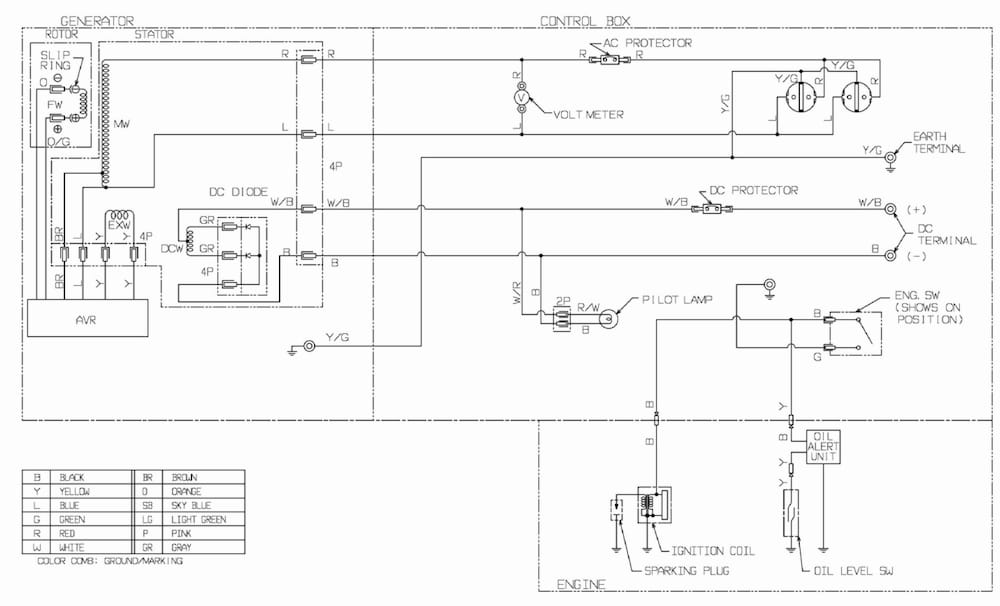 В соответствии с этой структурой теория когнитивных карт предсказывает, что небольшой набор клеток-мест должен активироваться непрерывно и в одной и той же фазе тета-цикла, пока голова и тело животного не смещены.Напротив, если последовательности сборки генерируются внутренними механизмами, нейронная активность должна постоянно меняться. Ключевым механизмом в последовательной организации ансамблей клеток гиппокампа является тета-осцилляция [65].
В соответствии с этой структурой теория когнитивных карт предсказывает, что небольшой набор клеток-мест должен активироваться непрерывно и в одной и той же фазе тета-цикла, пока голова и тело животного не смещены.Напротив, если последовательности сборки генерируются внутренними механизмами, нейронная активность должна постоянно меняться. Ключевым механизмом в последовательной организации ансамблей клеток гиппокампа является тета-осцилляция [65].
Тета-ритм представляет собой самоорганизующийся паттерн в септо-гиппокампальной системе и обеспечивает механизм сцепления нейронных сборок в соответствующих последовательностях (). В отсутствие скоординированных тета-осцилляций сжатые последовательности нейронных траекторий сильно нарушаются, что сопровождается поведенческими нарушениями в задачах, зависящих от гиппокампа [65–70].Поскольку тета-ритм представляет собой внутренний «таймер», единицы которого могут сжиматься и расширяться в зависимости от состояния мозга, он не точно имитирует ход времени часов (). Эта способность искажать время часов (и, следовательно, пространство) может объяснить, почему BOLD-сигнал в исследованиях на людях показывает различия между гиппокампальным представлением объективных пространственных расстояний по сравнению с запомненными расстояниями [71].
Эта способность искажать время часов (и, следовательно, пространство) может объяснить, почему BOLD-сигнал в исследованиях на людях показывает различия между гиппокампальным представлением объективных пространственных расстояний по сравнению с запомненными расстояниями [71].
Ранний эксперимент, демонстрирующий, что последовательная активность гиппокампа может быть внутренне связана с тета-колебаниями, показан на рис.Крыс обучали чередовать левое и правое руки модифицированного Т-образного лабиринта, и от них требовалось бегать в центральном колесе от 10 до 20 с примерно с той же скоростью, смотря в одном направлении в течение периода задержки между испытаниями. Поскольку и сигналы окружающей среды, и сигналы тела оставались постоянными благодаря этой конструкции, животное не могло использовать внешние сигналы, чтобы поддерживать информацию о выборе. Вместо этого он должен был полагаться на память о ранее выбранной правильной руке.
Последовательности сборки, сгенерированные внутри задачи памяти.gz50vkozhuheavr.jpg)
А. Центр: Крыса должна была бежать в бегущем колесе во время задержки между испытаниями, вспоминая свой последний выбор между левым и правым рукавами лабиринта. Точки представляют собой спайки одновременно зарегистрированных нейронов гиппокампа. Слева: нормализованная траектория частоты срабатывания нейронов во время вращения колеса, упорядоченная по латентности их пиковых частот срабатывания во время левых испытаний (каждая линия представляет собой отдельный нейрон). Справа: нормализованных скоростей возбуждения тех же нейронов во время испытаний справа.Экспериментатор может легко определить разницу между будущим левым или правым выбором, просто глядя на вектор сборки нейронов в любой момент во время работы колеса. B. Средние частоты срабатывания, каждая нормализованная до их собственной максимальной частоты выполнения задачи (синий-красный = 0–1), всех зарегистрированных нейронов BF (ось y) на всех этапах задачи (ось x). Зеленая, синяя, пурпурная и красная стрелки обозначают, соответственно, фазы световой вспышки, движения носа, перекрещивания пластины и остановки / вознаграждения задачи. C. То же, что и B, для нейронов, зарегистрированных в задней теменной коре. D. Образцы возбуждения пирамидных нейронов C A1 во время предпочтительных (первый столбец) и не предпочтительных (второй столбец) испытаний выбора. В третьем столбце показаны сегменты со значительно более высокой скоростью разряда в испытаниях «выбор-L» (красный) или «выбор-R» (синий). E. Кривые настройки клеток «целевое расстояние» в гиппокампе летучих мышей. Обратите внимание, что интервалы между фазами задач имеют разную и переменную длительность часов и организуют нейроны в соответствии с внутренними потребностями, а не с внешним (часовым) временем. A, по [72]; B-E, воспроизведено из [34,82,83].
Зеленая, синяя, пурпурная и красная стрелки обозначают, соответственно, фазы световой вспышки, движения носа, перекрещивания пластины и остановки / вознаграждения задачи. C. То же, что и B, для нейронов, зарегистрированных в задней теменной коре. D. Образцы возбуждения пирамидных нейронов C A1 во время предпочтительных (первый столбец) и не предпочтительных (второй столбец) испытаний выбора. В третьем столбце показаны сегменты со значительно более высокой скоростью разряда в испытаниях «выбор-L» (красный) или «выбор-R» (синий). E. Кривые настройки клеток «целевое расстояние» в гиппокампе летучих мышей. Обратите внимание, что интервалы между фазами задач имеют разную и переменную длительность часов и организуют нейроны в соответствии с внутренними потребностями, а не с внешним (часовым) временем. A, по [72]; B-E, воспроизведено из [34,82,83].
Когда одновременно регистрируется достаточное количество нейронов, весь путь в колесе связан с нейронной траекторией постоянно меняющихся клеточных ансамблей. Пирамидные нейроны срабатывают в определенное время во время бега колеса в каждом испытании или на тех же определенных расстояниях, пробегаемых с начала каждого испытания. Поскольку при беге в колесе животное не смещается, эти активные нейроны не соответствуют критериям клеток места. Вместо этого развивающаяся нейронная траектория должна отражать некоторый когнитивный контент.В поддержку этой гипотезы, нейронные траектории уникальны для испытаний левого или правого выбора, указывая на то, что начальное условие — установленное вознаграждением или запланированным действием — определяет образец траектории нейрона и отражает будущий выбор с высокой точностью, включая ошибки. (; [72]). Важно отметить, что содержимое нейронных сборок последовательных тета-циклов сдвигается во время движения колеса в задаче памяти так же, как и во время ходьбы при исследовании лабиринта. В целом, эти находки показывают, что даже несмотря на то, что заметные пространственные или идеотетические сигналы могут влиять на паттерны активации пространственных клеток, они не являются необходимыми для индукции последовательностей сборки клеток.
Пирамидные нейроны срабатывают в определенное время во время бега колеса в каждом испытании или на тех же определенных расстояниях, пробегаемых с начала каждого испытания. Поскольку при беге в колесе животное не смещается, эти активные нейроны не соответствуют критериям клеток места. Вместо этого развивающаяся нейронная траектория должна отражать некоторый когнитивный контент.В поддержку этой гипотезы, нейронные траектории уникальны для испытаний левого или правого выбора, указывая на то, что начальное условие — установленное вознаграждением или запланированным действием — определяет образец траектории нейрона и отражает будущий выбор с высокой точностью, включая ошибки. (; [72]). Важно отметить, что содержимое нейронных сборок последовательных тета-циклов сдвигается во время движения колеса в задаче памяти так же, как и во время ходьбы при исследовании лабиринта. В целом, эти находки показывают, что даже несмотря на то, что заметные пространственные или идеотетические сигналы могут влиять на паттерны активации пространственных клеток, они не являются необходимыми для индукции последовательностей сборки клеток. Фактически, динамика гиппокампа по умолчанию — это постоянное изменение. Даже во время сна гиппокамп неоднократно генерирует нейронные последовательности [73].
Фактически, динамика гиппокампа по умолчанию — это постоянное изменение. Даже во время сна гиппокамп неоднократно генерирует нейронные последовательности [73].
Расставить ячейки и ячейки времени?
Хотя пробег на расстояние не является важным параметром в задаче памяти бега колеса (), расстояние и прошедшее время можно вычислить либо из эволюции векторов скорости возбуждения нейронных сборок, либо из их векторов тета-фазы [20]. Другими словами, одни и те же нейронные данные могут быть связаны с расстоянием или временем [16,71], в зависимости от того, какой измерительный прибор используется для сравнения.Обширные наборы данных, полученные во время поведения в лабиринте и принудительного бега по беговой дорожке или ходьбы по мячу, демонстрируют, что развивающиеся нейронные сборки, полученные в гиппокампе, префронтальной коре, энторинальной коре и теменной коре, можно использовать для определения расстояния или продолжительности бега [74–77] . В первую очередь, единицы измерительных инструментов определяют, интерпретирует ли экспериментатор нейрофизиологические данные как пространство или время и относится к ним как к ячейкам места или времени.
Во многих поведенческих ситуациях организм должен действовать с определенной степенью гибкости, достигая целей, которые широко варьируются в пространственных и временных масштабах.Оптимальной стратегией для таких задач может быть «нормализация по событию» расстояния или длительности интервалов между поведенческими стимулами. Такие специфичные для задачи интервалы изменения могут не иметь большого отношения к времени часов, но могут быть важны при объединении последовательных нейронных событий, где скорость изменения зависит от требований задачи или внутренних переменных (). В поддержку этой гипотезы, клеточные сборки, сформированные в фазе задачи, были описаны в базальных ганглиях [78,79], базальном переднем мозге [80], теменной коре [81,82] и гиппокампе [34,83].
Другой пример независимости нейронных последовательностей от времени — это переменная скорость, с которой такие последовательности могут воспроизводиться. Такая же последовательность нейронных событий может происходить по крайней мере на трех порядках во временном масштабе. Во время локомоторного поведения клетки места будут активировать поведенческих последовательностей , которые изменяются в пределах 1-2 секунд (определяется скоростью бега животных). Одновременно те же траектории воспроизводятся во временном масштабе 100–200 миллисекунд в течение тета-последовательностей , которые сами по себе адаптируются во времени к скорости бега и целевой дистанции ([41,84];).Во время сна и других проявлений поведения в сочетании с резкой волновой рябью в гиппокампе эти последовательности пульсаций могут «воспроизводить» предыдущий опыт во временном масштабе 10–20 миллисекунд. Это не уникально для пространственных траекторий и гиппокампа, поскольку сборки направления головы в таламусе и связанные с задачами последовательности в неокортексе могут реактивироваться во время не-REM-сна при многократном сжатии [85,86]. Таким образом, отдельные сети и их динамика не имеют единой «тактовой частоты», которая может быть напрямую связана с внешним миром.Скорее, нейронная динамика является гибкой и адаптивной, изменяя темп вычислений по мере того, как этого требуют факторы (например, поведение или состояние мозга).
Поскольку в мозгу нет датчиков для непосредственного определения расстояния или продолжительности, ни пространство, ни время не могут «вызвать» какие-либо изменения в мозгу. Следовательно, расстояние и продолжительность не могут быть выведены из первых принципов. Напротив, эти концепции неизбежно логичны и зависят от инструментов, созданных руками человека [9]. Хотя часто можно найти корреляцию между эволюцией некоторых показателей мозга и единиц часов и удобно разместить измеренные параметры на временной шкале, такую корреляцию может интерпретировать только экспериментатор, имеющий доступ как к измерениям мозга, так и к калиброванным приборам. продолжительность.Тем не менее, такая корреляция не дает оснований сделать вывод о том, что мозговые цепи либо чувствуют, либо вычисляют время.
Отслеживание времени или последовательные нейронные вычисления
Трудно оправдать гипотезу, согласно которой существуют выделенные нейронные сети, единственная функция которых — вычислять время. При тщательном изучении нейронные сборки всегда оказываются для вычисления чего-то еще, помимо отслеживания продолжительности. Например, было предложено, чтобы нейроны теменной коры вычисляли время [87–90].В то же время образцы возбуждения этих нейронов могут быть привязаны к нескольким опорным кадрам; включая эгоцентрические действия [91], объекты [92,93] и изученные маршруты [81]. Имея в виду такой разнообразный набор систем координат, кажется, что способность декодировать «время» из таких сигналов является просто следствием системы, способной нести «систему отсчета, определяемую любым произвольным набором точек» [94] . В этом ключе и аналогично гиппокампу [34] базальные нейроны переднего мозга работают на уровне абстракции от расстояний и длительностей в физическом мире [52].В то время как время потенциала действия в этих структурах может коррелировать с пройденным расстоянием и истекшей продолжительностью, количество нейронов и сила их корреляций возрастают по отношению к фазе задачи, а не к единицам инструментов.
Генерация последовательности
Вместо таймера можно рассматривать гиппокамп как генератор последовательностей общего назначения, который несет ограниченную по содержанию порядковую структуру и покрывает промежутки между событиями или местами, которые необходимо связать [11]. Навигация в реальном или ментальном пространстве по своей природе представляет собой последовательность событий.Возможно, это все, что делает система гиппокампа: производит ограниченные по содержимому последовательности сборки клеток без кодирования деталей конкретных событий и предсказывает лучшие варианты на основе предыдущего опыта [95]. Классическое исследование Говарда Эйхенбаума «транзитивный вывод» также указывает на критическую важность порядка нейронов [96,97]. Крыс обучали различать пары запахов, которые перекрываются друг с другом (то есть запах «А»> запах «В»; запах «В»> запах «С» и т. Д.). После изучения парных ассоциаций крыса могла «построить» последовательность, которая позволяла ей делать выводы о взаимосвязях между парами запахов, которые ранее не были изучены (например,g., запах «А»> запах «В»). Крысы с поражениями гиппокампа изучили индивидуальные различия, но не смогли правильно определить взаимосвязь между новой парой (запах «А»> запах «С»).
Будь то стержень или часы , последовательности в системе гиппокампа могут просто указывать на элементы (что), хранящиеся в неокортексе, в том же порядке, в каком они были испытаны во время обучения. Подобные теории предполагают, что это разделение труда аналогично роли библиотекаря (гиппокампа, указывающего на желаемые предметы) в библиотеке (неокортекс, где хранятся семантические знания; [98–102].Ключевое различие между нашей текущей точкой зрения и теорией индексации [98] — это порядок событий. Вместо единственного индекса, который « указывает » на корковые модули, « единица », с нашей точки зрения, представляет собой последовательность индексов (« мультиплексированный указатель »), так что гиппокампальная система отвечает за конкатенацию неокортикальных блоков информации в последовательности как для кодирования, так и для кодирования. извлечение (). С этой точки зрения ковариация активности между модальностями приводит к формированию последовательности гиппокампа, которая может быть отделена от индивидуальных модальностей, так что последовательности будут лучше коррелировать с этими ковариатами, которые часто представляют собой расстояние или продолжительность с текущими экспериментальными парадигмами.Однако продемонстрированная способность таких последовательностей искажаться относительно измеренных извне расстояния и продолжительности предполагает, что это внутреннее «представление» пространства или времени было бы крайне неточным отражением внешнего мира. В рамках предлагаемой теории секвенирования эпизодическая память — это просто упорядоченная последовательность трансляционно-инвариантных «что» и связанных с ними контекстов (еще одно «что») без явного внутреннего «представления» пространства или времени.
Гипотеза секвенирования гиппокампа.Индексы, указывающие на корковые и подкорковые модули для различных входов, упорядочиваются паттернами активности гиппокампа, таким образом сохраняя порядковую структуру, по которой происходит переживание.
Подтверждение исследований на людях
Гипотеза о генераторе последовательностей гиппокампа подтверждается данными как поражений, так и изображений. Как и в экспериментальных исследованиях на животных, в большинстве исследований на людях отдельно изучались нейронные корреляты пространства [24,103-108] или времени [16,17,109-112], постулируемых контекстуальных компонентов эпизодической памяти.
У пациентов с амнезией с повреждением гиппокампа гораздо меньше проблем с оценкой и запоминанием расстояний и продолжительности, чем с запоминанием последовательного порядка событий, в которых они произошли [103,113]. Точно так же крысы с повреждением гиппокампа демонстрируют нарушение памяти на порядок запаховых стимулов, в отличие от их сохраненной способности распознавать запахи, которые возникли недавно [114].
Эксперименты по визуализации человека также предоставили поддержку функции генератора последовательности гиппокампа, без явных карт пространства или времени.Недавний эксперимент сравнил прошедшую продолжительность между представлениями объектов, встроенных в последовательность. Полученные данные показали, что паттерны активности гиппокампа несут информацию о ранговых позициях объектов в усвоенных, но не случайных последовательностях, даже когда временные положения объектов были одинаковыми [111]. В другом недавнем исследовании использовались временные метки GPS и камеры для моментальных снимков событий, автоматически сделанных участниками, которые носили камеру в течение четырех недель. Впоследствии, когда участников сканировали с помощью фМРТ, вспоминая свой реальный жизненный опыт, их гиппокампальный паттерн активации был подобен пространственным и временным расстояниям в различных масштабах [115].В исследовании виртуальной реальности испытуемые перемещались по улицам виртуального города, и их просили вспомнить, были ли два объекта близко или далеко. В этом эксперименте нейронное сходство также масштабировалось с учетом близости воспоминаний о событиях как в пространственном, так и во временном измерении, поддерживая представление об общем механизме кодирования гиппокампа для пространства и времени [71].
Независимая серия исследований изучала, как можно понять опыт через последовательность событий [116]. Было обнаружено, что память последовательностей более точна для элементов, представленных в одном контексте, по сравнению с событиями, разделенными границами [117], и что элементы, появляющиеся в одном контексте, позже оцениваются как более близкие друг к другу [110], даже если фактическое расстояние было такой же.Более высокое сходство паттернов BOLD-активности наблюдалось во время испытаний заданий для заданий, которые были субъективно оценены как « близкие », по сравнению с заданиями, которые позже были оценены как « далеко », даже если пары пунктов были разделены таким же количеством промежуточных испытаний в задании. [110]. В целом, эксперименты с поведением, повреждениями и визуализацией подтверждают гипотезу о том, что кодирование и сохранение последовательного порядка переживаний является важной функцией гиппокампа. Изучение последовательного порядка событий позволяет нам формировать прогнозы о надвигающемся будущем и соответствующим образом планировать предстоящие действия.
Трансдукция последовательностей сборки гиппокампа в действие
Классификация нейронов гиппокампа на клетки места, клетки времени или и то и другое экспериментатором может не иметь отношения к мозгу. Важно то, как нижестоящие механизмы чтения классифицируют сообщения гиппокампа. Если не считать демонстрации того, что существуют отдельные методы дешифрования для расстояния и продолжительности, мы не можем заключить, что гиппокамп «кодирует» ни пространство, ни время. Чтобы продемонстрировать полезность гипотетической когнитивной карты, нам необходимо изучить, как такая информация считывается нижележащими механизмами считывания, чтобы направлять действия животного.
Поскольку нейроны гиппокампа не имеют прямых проекций на мышцы, их вычисления должны быть перенесены в другое место, чтобы информировать механизмы выбора действия. Возможные выходные структуры включают энторинальную кору, субикулум и ретросплениальную кору, префронтальную кору и ядра перегородки [25]. Из них наиболее вероятным каналом является латеральная перегородка [118]. Из всех мишеней он получает самые плотные проекции гиппокампа. В то время как единственной кортикофугальной проекцией из гиппокампа является область CA1, нейроны в латеральной перегородке получают входы как CA1, так и CA3 [23].Боковая перегородка также имеет прямые проекции на мотивационные центры гипоталамуса [119, 120], средней линии таламуса [121] и ствола мозга [23]; потенциально позволяя выбирать программы действий. Если тета-шаговая последовательность клеточных сборок в гиппокампе релевантна для такой гипотетической трансформации карты в действие, то латеральная перегородка должна быть способна читать эту синтаксическую структуру. Однако предыдущие исследования записи обнаружили только сильно деградированную пространственную информацию в частотах возбуждения нейронов латеральной перегородки [122–125].
Ключевым аспектом этой цепочки рассуждений является то, что механизм считывания нейронов в боковой перегородке должен быть связан не с внешним миром, а с последовательно упорядоченной активностью его входов. В этом случае формат нейронной активности в боковой перегородке может отличаться от формата пространственных корреляций внутри гиппокампа. Действительно, выброс нейронов латеральной перегородки показывает очень надежную корреляцию между их тета-фазой и положением животного. Это чистый «фазовый код», потому что частота возбуждения нейронов латеральной перегородки практически не несет информации о месте, которое занимает животное, и требует, чтобы фазовая информация от гиппокампа наблюдалась ().В пределах одного тета-цикла потенциалы действия нейронных сборок латеральной перегородки коррелируют как с текущим положением, так и с расстоянием от начала и до конца изученной траектории в пространстве. Этот не зависящий от скорости фазовый код не наследуется от информации о фазе или скорости отдельных вышестоящих клеток места гиппокампа, но отражает вычисление постоянно изменяющейся взаимосвязи между сборками клеток места СА1 и СА3 [118]. С точки зрения нейронов латеральной перегородки, тета-последовательности нейронных ансамблей CA3 и CA1 являются «единицей» передачи информации из гиппокампа.Предполагая аналогичные механизмы трансформации в нижележащих цепях гипоталамуса и ствола мозга, такой механизм «селектора действия» может быстро идентифицировать источник информации и, таким образом, опосредовать действие с карты более высокого порядка.
Частотно-независимые корреляции между фазой и положением нейронов латеральной перегородки.A. Нейрон в боковой перегородке демонстрирует фазовую прецессию относительно тета-фазы CA1. Цветные точки указывают на возникновение потенциалов действия от одного нейрона латеральной перегородки, когда крыса пересекает круговой путь (~ 3.2 метра; N = 32 испытания). Цвета соответствуют фазам тета-колебаний C A1. B. Реконструкция положения крысы (среднеквадратичная ошибка, MSE) по скорости стрельбы (красный) или по соотношению фаза-положение спайка (синий) относительно контроля (перетасовка). Воспроизведено из [118]
Заключительные замечания
Пространство и время, как организующие вместилища исследований классической физики и нейробиологии, будут и дальше служить источником множества исследований. Конечно, нет ничего плохого в том, чтобы использовать измерительные приборы и соотносить наши наблюдения с их единицами.Формулировка пространства-времени физиками не дискредитировала часы. Напротив, физика и все другие науки все больше полагаются на точность измерительных инструментов для калибровки экспериментальных наблюдений. Было бы немыслимо проводить современные нейробиологические исследования без обращения к пространственным и временным единицам. Однако заявить на основе таких сравнений, что какая-то область мозга или механизм «представляет» пространство и время или вычисляет расстояние и продолжительность, — это другое дело. Во многих случаях мы могли просто не найти правильные правила преобразования, которые могли бы более эффективно объяснить динамику срабатывания нейронов, чем расстояние или продолжительность [52] (вставка для нерешенных вопросов).
Чтобы осмысленно интерпретировать значение любого нейронного паттерна, очень важно изучить, как такие паттерны используются нижестоящими механизмами нейронного наблюдателя. Несущественно вызывать наборы нейронов гиппокампа, клетки места или клетки времени без демонстрации их дифференциального воздействия на нейроны-партнеры ниже по течению. Мы проиллюстрировали эту стратегию здесь, продемонстрировав, что преобразование паттернов активности нейронов гиппокампа, концептуализированных как когнитивная карта, в чистый фазовый код в латеральной перегородке зависит от нейронных последовательностей, управляемых тета-колебаниями.Изучение правил трансформации между структурами мозга не требует обращения к концепциям пространства или времени. Фактически, это преобразование, вероятно, существует для всех последовательностей, продуцируемых в гиппокампе или других структурах, независимо от того, относим ли мы эти последовательности к пространству, времени или другим когнитивным конструкциям.
Основные моменты
Мы предлагаем, чтобы гиппокамп выполнял общий, но единичный алгоритм: создание последовательной структуры без содержания для доступа и организации сенсорных ощущений, распределенных по кортикальным модулям
Нейронные «репрезентации» можно ссылаться на множество кадров , и прямые сравнения между системами отсчета могут использоваться при попытке идентифицировать лежащие в основе нейронные вычисления
Нейронную динамику и преобразования можно описать, не прибегая к концепциям пространства и времени
Дальнейшие исследования должны сосредоточиться на правилах преобразования между конструкции, а не на тюнинг.Подход, ориентированный на нейронного наблюдателя, который сравнивает две внутренние переменные, может быть более плодотворным, чем корреляция внешнего сигнала с нейронными паттернами.
Поле для нерешенных вопросов
Можно ли приписывать пространству и времени двойные роли: пространство-время в физике и ортогональные координаты пространства и времени для вычислений мозга?
Если концепции пространства и времени зависят от измеряемых приборами вариантов расстояния и продолжительности, как животные, не относящиеся к человеку, могут концептуализировать или использовать эти концепции?
Можем ли мы описать нейронные вычисления, не обращаясь к внешней пространственной и временной структуре?
Необходимо ли предполагать внутреннюю временную структуру для определения нейронных вычислений?
Существует ли активное поведение, при котором гиппокамп не обеспечивает надежной последовательной активности, связанной с заданием?
Активность населения в любой структуре мозга позволяет отслеживать время по часам.Существуют ли нейроны и сети, единственная функция которых состоит в отслеживании времени, или это тот случай, когда развивающаяся нейронная активность всегда вычисляет что-то еще (также)?
Если в гиппокампе есть выделенные ячейки места, ячейки времени, ячейки отслеживания тона, ячейки цели или ячейки вознаграждения, кто их читает? Постулирование таких специализированных функций полезно для мозга только в том случае, если существуют последующие классификаторы таких предполагаемых уникальных функций.
Доминирующий взгляд на консолидацию памяти предполагает, что опыт временно сохраняется в гиппокампе и с течением времени передается в неокортекс.Это потребует достоверного кодирования информации о пробуждении в гиппокампе. В качестве альтернативы, возможно ли, что информация о бодрствовании всегда хранится в неокортексе, а критическая роль гиппокампа состоит в организации упорядоченной последовательности событий?
Благодарности:
Мы посвящаем нашу перспективу покойному Говарду Эйхенбауму, который предупредил нас о нечеткой природе пространства и времени в исследованиях нейробиологии. Мы также хотели бы поблагодарить Кэтрин МакКлейн за критические отзывы о предыдущей версии этой рукописи.Эта работа была поддержана грантами Национального института здравоохранения США ({«type»: «entrez-nucleotide», «attrs»: {«text»: «Mh207396», «term_id»: «1386680410»}} Mh207396, NS099705, NS104590), Фонд Мазерса и Фонд Саймонса. NSF PIRE.
Глоссарий
| Поведенческая последовательность | Последовательность нейронной активности, которая изменяется со скоростью, с которой организмы ведут себя и взаимодействуют с окружающей средой |
| Расстояние | Кратчайший путь между двумя положениями.Это определение делает взаимосвязь между позициями и расстоянием круговой |
| Продолжительность | Интервал между двумя временными точками. Это определение устанавливает взаимосвязь между временными точками (например, рождение Христа и сегодняшний день) и продолжительностью круговой |
| Гамма-ритм / колебание | Паттерны локального поля ритма 30–120 Гц. Гамма-колебания — признак конкуренции между возбуждающими и тормозящими нейронами. Он отражает временный баланс (колеблющиеся выигрыш и проигрыш) в любом мозговом контуре с этими типами нейронов.Амплитуда гамма-колебаний часто модулируется фазой более медленных колебаний (таких как тета) |
| Ячейки сетки | Нейроны в медиальной энторинальной коре и парапубикулуме, чья повышенная активность в вершинах воображаемых равносторонних треугольников на полу метки окружающей среды, положение животного обозначается клетками сетки |
| Гиппокампально-энторинальная система | Компоненты систем работают не изолированно, а во взаимодействии.Гиппокамп и энторинальная кора часто работают вместе как единое целое, дополняемое субикулярным комплексом. |
| Поместите клетки | Пирамидные нейроны и гранулярные клетки в гиппокампе, которые последовательно активируются в определенных частях окружающей среды, определены как клетки места |
| Пульсирующая последовательность | Последовательность нервной активности, которая коррелирует с опытом организма, но происходит во временной шкале, сжатой в ~ 10–20 раз.Примеры этого в некоторых случаях называют «воспроизведением». См. [129–131] |
| Стержни и часы | Все измерительные приборы имеют единицы, которые произвольно определяются их разработчиком. Интервалы этих единиц всегда являются относительными и всегда относятся к мере, предоставляемой другим инструментом (например, период маятника может быть откалиброван другими более быстрыми часами) или естественным событием (например, лунный цикл, частота сердечных сокращений, окружность мира ) |
| Тета-ритм / колебания | Колебательный паттерн от 4 до 9 Гц в локальном поле гиппокампа, когда животное занято исследовательским поведением или быстрым сном.Он отражает когерентные, ритмичные колебания трансмембранного потенциала во многих нейронах |
| Тета-последовательность | Последовательность нейронной активности, которая коррелирует с восприятием организма, но происходит в ~ 10-кратном сжатом временном масштабе. Примеры этого часто называют «прогнозированием». См. [38,41,84] |
| Временные клетки | В гиппокампе и энторинальной коре основные клетки, которые генерируют кластеры спайков в отсутствие смещения животного (например,ж., бег в колесе или на беговой дорожке) называются ячейками времени. Частота возбуждения отдельных нейронов в нескольких структурах мозга часто увеличивается в зависимости от некоторой экспериментальной переменной (например, задержки) до некоторого порогового значения. В принципе, накопление потенциалов действия нейрона или популяции нейронов можно использовать для отслеживания времени. |
Сноски
Заявление издателя: Это PDF-файл неотредактированной рукописи, принятой для публикации.В качестве услуги для наших клиентов мы предоставляем эту раннюю версию рукописи. Рукопись будет подвергнута копированию, верстке и рассмотрению полученного доказательства, прежде чем она будет опубликована в окончательной форме для цитирования. Обратите внимание, что во время производственного процесса могут быть обнаружены ошибки, которые могут повлиять на содержание, и все юридические оговорки, относящиеся к журналу, имеют отношение.
Ссылки
1. Scoville WB и Milner B (1957) Потеря недавней памяти после двусторонних поражений гиппокампа.J. Neurol. Нейрохирургия. Психиатрия 20, 11–21 [Бесплатная статья PMC] [PubMed] [Google Scholar] 2. Tulving E (1985) Элементы эпизодической памяти, Clarendon. [Google Scholar] 4. Rovelli C (2018) Реальность — это не то, чем кажется: путешествие к квантовой гравитации. [Google Scholar] 7. Sinha C и другие. (2011) Когда время — это не пространство: социальное и лингвистическое построение временных интервалов и временных отношений событий в амазонской культуре. Lang. Cogti. 3, 137–169 [Google Scholar] 8. Каналес Дж. (2016) Физик и философ: Эйнштейн, Бергсон и дебаты, изменившие наше понимание времени, Princeton University Press.[Google Scholar] 10. Деннетт Д.К. и Кинсборн М. (1992) Время и наблюдатель: где и когда возникает сознание в мозгу. Behav. Brain Sci. 15, 183–201 [Google Scholar] 11. Фристон К. и Бужаки Г. (2016) Функциональная анатомия времени: что и когда в мозге. Trends Cogn. Sci. 20, 500–511 [PubMed] [Google Scholar] 13. Толман EC (1948) Когнитивные карты у крыс и людей. Psychol. Ред. 55, 189–208 [PubMed] [Google Scholar] 14. О’Киф Дж. И Надел Л. (1978) Гиппокамп как когнитивная карта, Оксфорд: Clarendon Press.[Google Scholar] 15. Мозер Э.И. и другие. (2014) Сетчатые клетки и корковое представление. Nat. Rev. Neurosci. 15, 466–481 [PubMed] [Google Scholar] 18. О’Киф Дж. И Достровский Дж. (1971) Гиппокамп как пространственная карта. Предварительные свидетельства активности единицы у свободно передвигающейся крысы Brain Res. 34, 171–175 [PubMed] [Google Scholar] 19. Hafting T и другие. (2005) Микроструктура пространственной карты энторинальной коры Природа 436, 801–806 [PubMed] [Google Scholar] 20. Ицков В и другие. (2011) Последовательности сборки клеток, возникающие в результате адаптации к спайковому порогу. Следите за временем в гиппокампе.J. Neurosci. 31, 2828–2834 [Бесплатная статья PMC] [PubMed] [Google Scholar] 22. Amaral DG и Lavenex P (2006) Нейроанатомия гиппокампа. В Книге Гиппокампа; С. 45–66 [Google Scholar] 23. Risold PY и Swanson LW (1997) Соединения латерального перегородочного комплекса крысы. Brain Res. Ред. 24, 115–195 [PubMed] [Google Scholar] 24. Иглой К и другие. (2010) Латерализованная активность гиппокампа человека предсказывает навигацию на основе памяти последовательности или места. Proc. Natl. Акад. Sci. 107, 14466–14471 [Бесплатная статья PMC] [PubMed] [Google Scholar] 25.Амарал Д.Г. и Виттер М.П. (1989) Трехмерная организация образования гиппокампа: обзор анатомических данных. Неврология 31, 571–591 [PubMed] [Google Scholar] 26. Виттнер Л и другие. (2007) Трехмерная реконструкция ветви аксона пирамидной клетки CA3, записанная и заполненная in vivo. Структура мозга. Фонд. 212, 75–83 [Бесплатная статья PMC] [PubMed] [Google Scholar] 27. Маурер А.П. и другие. (2005) Самодвижение и происхождение дифференциального пространственного масштабирования вдоль перегородочно-временной оси гиппокампа.Гиппокамп 15, 841–852 [PubMed] [Google Scholar] 28. Ranganath C и Ritchey M (2012) Две корковые системы для поведения, управляемого памятью. Nat. Rev. Neurosci. 13, 713–726 [PubMed] [Google Scholar] 30. Buzsáki G (2002) Тета-колебания в гиппокампе. Нейрон 33, 325–340 [PubMed] [Google Scholar] 31. Любенов Е.В., Сиапас А.Г. (2009) Тета-колебания гиппокампа — это бегущие волны. Природа 459, 534–9 [PubMed] [Google Scholar] 34. Терада С и другие. (2017) Временное и скоростное кодирование дискретных последовательностей событий в гиппокампе.Нейрон 94, 1248–1262.e4 [PubMed] [Google Scholar] 35. Миливоевич Б. и Доеллер К.Ф. (2013) Мнемонические сети в формировании гиппокампа: от пространственных карт до временных и концептуальных кодов. J. Exp. Psychol. Gen. 142, 1231–1241 [PubMed] [Google Scholar] 36. Эйхенбаум Х и другие. (1999) Гиппокамп, клетки памяти и места: это пространственная память или пространство памяти? Нейрон 23, 209–226 [PubMed] [Google Scholar] 37. Маккензи С и другие. (2014) Гиппокампальное представление связанных и противоположных воспоминаний развивается в рамках отдельных, иерархически организованных нейронных схем.Нейрон 83, 202–215 [Бесплатная статья PMC] [PubMed] [Google Scholar] 38. Skaggs WE и другие. (1996) Прецессия тета-фазы в нейрональных популяциях гиппокампа и сжатие временных последовательностей. Гиппокамп 6, 149–172 [PubMed] [Google Scholar] 39. Драгой Г и другие. (2006) Временное кодирование последовательностей мест сборками клеток гиппокампа. Нейрон 50, 145–157 [PubMed] [Google Scholar] 40. Диба К. и Бузаки Г. (2008) Динамика сети гиппокампа ограничивает временную задержку между пирамидными клетками в модифицированных средах.J. Neurosci. 28, 13448–13456 [Бесплатная статья PMC] [PubMed] [Google Scholar] 43. Лисман Дж. Э. и Идарт М. А. П. (1995) Хранение 7 +/- 2 кратковременных воспоминаний в колебательных подциклах. Наука 267, 1512–1515 [PubMed] [Google Scholar] 44. Харрис К.Д. и другие. (2003) Организация клеточных сборок в гиппокампе. Природа 424, 552–556 [PubMed] [Google Scholar] 45. Дженсен О. и Лисман Дж. Э. (2005) Кодирование последовательности гиппокампа, управляемое кортикальным многопозиционным буфером рабочей памяти. Trends Neurosci. 28, 67–72 [PubMed] [Google Scholar] 46.O’Keefe J и Recce ML (1993) Фазовая взаимосвязь между единицами места в гиппокампе и тета-ритмом ЭЭГ. Гиппокамп 3, 317–330 [PubMed] [Google Scholar] 47. О’Киф Дж. И Берджесс Н. (1996) Геометрические детерминанты пространственных полей нейронов гиппокампа. Природа 381, 425–428 [PubMed] [Google Scholar] 48. Готард КМ и другие. (1996) Динамика коррекции несоответствия в коде ансамбля гиппокампа для пространства: взаимодействие между интеграцией путей и сигналами окружающей среды. J. Neurosci. 16, 8027–8040 [Бесплатная статья PMC] [PubMed] [Google Scholar] 49.Джонс М.В. и Уилсон М. а. (2005) Прецессия фазы медиальной префронтальной корковой активности относительно тета-ритма гиппокампа. Гиппокамп 15, 867–873 [PubMed] [Google Scholar] 50. Hafting T и другие. (2008) Независимая от гиппокампа фазовая прецессия в клетках энторинальной сетки. Природа 453, 1248–1252 [PubMed] [Google Scholar] 51. van der Meer M. a a и Redish a D (2011) Прецессия тета-фазы в вентральном полосатом теле крысы связывает информацию о месте и вознаграждении. J. Neurosci. 31, 2843–2854 [Бесплатная статья PMC] [PubMed] [Google Scholar] 53.Макнотон Б.Л. и другие. (1996) Расшифровка полиглота гиппокампа: гиппокамп как система интеграции путей. J. Exp. Биол. 199, 173–185 [PubMed] [Google Scholar] 55. Foster TC и другие. (1989) Пространственная селективность нейронов гиппокампа крыс: зависимость от готовности к движению. Наука 244, 1580–1582 [PubMed] [Google Scholar] 56. Czurkó A и другие. (1999) Устойчивая активация пирамидных клеток гиппокампа за счет «сжатия пространства» в беговом колесе. Евро. J. Neurosci. 11, 344–352 [PubMed] [Google Scholar] 57.Taube JS и другие. (1990) Клетки направления головы, записанные из заднебукулярной кости у свободно движущихся крыс. I. Описание и количественный анализ. J. Neurosci. 10, 420–35 [Бесплатная статья PMC] [PubMed] [Google Scholar] 58. Гейслер С и другие. (2007) Клетки места гиппокампа представляют собой осцилляторы с регулируемой скоростью. Proc. Natl. Акад. Sci. USA 104, 8149–8154 [Бесплатная статья PMC] [PubMed] [Google Scholar] 59. Юнг МВт и другие. (1994) Сравнение пространственных характеристик возбуждения единиц в дорсальном и вентральном гиппокампе крысы.J. Neurosci. 14, 7347–7356 [Бесплатная статья PMC] [PubMed] [Google Scholar] 60. Kjelstrup KB и другие. (2008) Конечная шкала пространственного представления в гиппкампе. Наука 321, 140–143 [PubMed] [Google Scholar] 61. Bose A и Recce M (2001) Фазовая прецессия и фазовая синхронизация пирамидных клеток гиппокампа. Гиппокамп 11, 204–215 [PubMed] [Google Scholar] 64. О’Киф Дж. И Берджесс Н. (2005) Двойное фазовое и скоростное кодирование в клетках места гиппокампа: теоретическое значение и связь с клетками энторинальной решетки.Гиппокамп 15, 853–866 [Бесплатная статья PMC] [PubMed] [Google Scholar] 65. Ван И и другие. (2015) Тета-последовательности необходимы для внутренних полей возбуждения гиппокампа. Nat. Neurosci. 18, 282–288 [PubMed] [Google Scholar] 66. Уинсон Дж. (1978) Потеря тета-ритма в гиппокампе приводит к дефициту пространственной памяти у крыс. Наука 201, 160–163 [PubMed] [Google Scholar] 67. Mizumori SJY и другие. (1990) Обратимая инактивация медиальной перегородки по-разному влияет на две формы обучения у крыс.Brain Res. 528, 12–20 [PubMed] [Google Scholar] 68. Као Хи и другие. (2017) Фенциклидин дискоординирует сетевую активность гиппокампа, но не определяет поля. J. Neurosci. 37, 12031–12049 [Бесплатная статья PMC] [PubMed] [Google Scholar] 69. Ньюман Э.Л. и другие. (2017) Точная динамика временных спайков активности клеток гиппокампа, чувствительных к холинергическим нарушениям. Гиппокамп 27, 1069–1082 [Бесплатная статья PMC] [PubMed] [Google Scholar] 70. Робби Д и другие. (2006) Каннабиноиды показывают важность координации времени спайков в функции гиппокампа.Nat. Neurosci. 9, 1526–1533 [PubMed] [Google Scholar] 73. Buzsáki G (2015) Резкая волновая рябь в гиппокампе: когнитивный биомаркер эпизодической памяти и планирования. Гиппокамп 25, 1073–1188 [Бесплатная статья PMC] [PubMed] [Google Scholar] 75. Fujisawa S и другие. (2008) Поведенчески-зависимая краткосрочная динамика сборки в медиальной префронтальной коре. Nat. Neurosci. 11, 823–833 [Бесплатная статья PMC] [PubMed] [Google Scholar] 76. Харви CD и другие. (2012) Последовательности, специфичные для выбора, в теменной коре во время решения задачи виртуальной навигации.Природа 484, 62–68 [Бесплатная статья PMC] [PubMed] [Google Scholar] 77. Говард Л.Р. и другие. (2014) Гиппокамп и энторинальная кора кодируют путь и евклидовы расстояния до целей во время навигации. Curr. Биол. 24, 1331–1340 [Бесплатная статья PMC] [PubMed] [Google Scholar] 78. Graybiel AM (1998) Базальные ганглии и фрагменты репертуара действий. Neurobiol. Учиться. Mem. 70, 119–136 [PubMed] [Google Scholar] 79. Джин Икс и другие. (2014) Подсхемы базальных ганглиев отчетливо кодируют синтаксический анализ и конкатенацию последовательностей действий.Nat. Neurosci. 17, 423–430 [Бесплатная статья PMC] [PubMed] [Google Scholar] 81. Nitz D. a (2006) Отслеживание продвижения пути в задней теменной коре. Нейрон 49, 747–56 [PubMed] [Google Scholar] 83. Сарел А и другие. (2017) Векторное изображение пространственных целей в гиппокампе летучих мышей. Наука 355, 176–180 [PubMed] [Google Scholar] 85. Юстон DR и другие. (2007) Быстрое воспроизведение недавних последовательностей памяти в префронтальной коре во время сна. Наука 318, 1147–1150 [PubMed] [Google Scholar] 87.Леон М.И. и Шадлен М.Н. (2003) Представление времени нейронами задней теменной коры макака. Нейрон 38, 317–327 [PubMed] [Google Scholar] 88. Янссен П. и Шадлен М.Н. (2005) Представление степени опасности прошедшего времени в области LIP макак. Nat. Neurosci. 8, 234–241 [PubMed] [Google Scholar] 89. Лебедев М.А. и другие. (2008) Расшифровка временных интервалов по активности коркового ансамбля Михаил. J. Neurophysiol. 99, 166–186 [PubMed] [Google Scholar] 91. Макнотон Б.Л. и другие. (1994) Кортикальное представление движения во время неограниченной пространственной навигации у крысы.Цереб. Кора 4, 27–39 [PubMed] [Google Scholar] 92. Олсон К.Р. и Геттнер С.Н. (1995) Объектно-ориентированная направленная селективность в дополнительном поле глаза макака. Наука 269, 985–989 [PubMed] [Google Scholar] 93. Чафи ΜV и другие. (2007) Представление пространственных отношений в задней теменной коре: позиция с привязкой к объекту кода одиночных нейронов. Цереб. Кора 17, 2914–2932 [PubMed] [Google Scholar] 94. Nitz D (2009) Теменная кора, навигация и построение произвольных систем отсчета для пространственной информации.Neurobiol. Учиться. Mem. 91, 179–185 [PubMed] [Google Scholar] 95. Stachenfeld KL и другие. (2017) Гиппокамп как прогностическая карта. Nat. Neurosci. 20, 1643–1653 [PubMed] [Google Scholar] 96. Bunsey M и Eichenbaum HB (1996) Сохранение функции памяти гиппокампа у крыс и людей. Природа 379, 255–7 [PubMed] [Google Scholar] 98. Тейлер TJ и DiScenna P (1986) Теория индексации памяти гиппокампа. Behav. Neurosci. 100, 147–154 [PubMed] [Google Scholar] 99. Buzsáki G (2006) Ритмы мозга, Oxford University Press.[Google Scholar] 100. Надел л. и Moscovitch M (1997) Реконсолидация памяти, ретроградная амнезия и гиппокампальный комплекс. Карр. Opin. Neurobiol. 7, 217–227 [PubMed] [Google Scholar] 101. Сквайр LR и другие. (1992) Память и гиппокамп: синтез результатов, полученных на крысах, обезьянах и людях. Psychol. Ред. 99, 195–231 [PubMed] [Google Scholar] 102. Марр Д. (1970) Простая память: теория Archicortex Филос. Пер. R. Soc. Лондон 262. [PubMed] [Google Scholar] 103. Maguire EA и другие. (2006) Лондонские водители такси и водители автобусов: структурная МРТ и нейропсихологический анализ.Гиппокамп 17, 801–812 [Google Scholar] 106. Кайл CT и другие. (2015) Успешное извлечение конкурирующих пространственных сред у людей связано с механизмами разделения паттернов гиппокампа. Элиф 4, 1–19 [Бесплатная статья PMC] [PubMed] [Google Scholar] 107. Васс Л.К. и Эпштейн Р.А. (2013) Абстрактные представления местоположения и направления взгляда в человеческом мозге. J. Neurosci. 33, 6133–6142 [Бесплатная статья PMC] [PubMed] [Google Scholar] 108. Вольберс Т и другие. (2007) Дифференциальное задействование гиппокампа, медиальной префронтальной коры и комплекса движений человека во время интеграции путей у людей.J. Neurosci. 27, 9408–9416 [Бесплатная статья PMC] [PubMed] [Google Scholar] 109. Ховард М.В. и Кахана М.Дж. (2002) Распределенное представление временного контекста. J. Math. Psychol. 46, 269–299 [Google Scholar] 110. Ezzyat Y и Davachi L (2014) Сходство порождает близость: сходство паттернов внутри и между контекстами связано с более поздними мнемоническими суждениями о временной близости. Нейрон 81, 1179–1189 [Бесплатная статья PMC] [PubMed] [Google Scholar] 112. Экстром А.Д. и Ранганат С. (2017) Пространство, время и эпизодическая память: гиппокамп находится на всей когнитивной карте.Гиппокамп [PubMed] [Google Scholar] 113. Dede AJO и другие. (2016) Изучение и запоминание реальных событий после повреждения медиальной височной доли. Proc. Natl. Акад. Sci. 113, 13480–13485 [Бесплатная статья PMC] [PubMed] [Google Scholar] 115. Nielson DM и другие. (2015) Человеческий гиппокамп представляет пространство и время во время восстановления воспоминаний реального мира. Proc. Natl. Акад. Sci. 112, 11078–11083 [Бесплатная статья PMC] [PubMed] [Google Scholar] 116. Davachi L и DuBrow S (2015) Как гиппокамп сохраняет порядок: роль предсказания и контекста.Trends Cogn. Sci. 19, 92–99 [Бесплатная статья PMC] [PubMed] [Google Scholar] 117. DuBrow S и Davachi L (2013) Влияние границ контекста на память для последовательного порядка событий. J. Exp. Psychol. Gen. 142, 1277–1286 [Бесплатная статья PMC] [PubMed] [Google Scholar] 119. Swanson LW и Cowan WM (1979) Соединения перегородки у крысы. J. Comp. Neurol. 186, 621–656 [PubMed] [Google Scholar] 120. Вонг LC и другие. (2016) Эффективная модуляция мужской агрессии через боковую перегородку к медиальной проекции гипоталамуса.Curr. Биол. 26, 593–604 [Бесплатная статья PMC] [PubMed] [Google Scholar] 122. Нисидзё Х и другие. (1997) Нейронные реакции перегородки, связанные с пространственным представлением у обезьян. Гиппокамп 464, 460–464 [PubMed] [Google Scholar] 123. Чжоу Т.Л. и другие. (1999) Сравнение активности нейронов медиальной и латеральной перегородки при выполнении пространственных задач у крыс. Гиппокамп 9, 220–234 [PubMed] [Google Scholar] 124. Leutgeb S и Mizumori SJY (2002) Контекстно-зависимые пространственные представления латеральными перегородочными клетками. Неврология 112, 655–663 [PubMed] [Google Scholar] 125.Такамура Y и другие. (2006) Свойства пространственного возбуждения нейронов латеральной перегородки. Гиппокамп 16, 635–644 [PubMed] [Google Scholar] 126. Mau W и другие. (2018) Одна и та же популяция гиппокампа CA1 одновременно кодирует временную информацию в нескольких временных масштабах. Curr. Биол. 28, 1–10 [Бесплатная статья PMC] [PubMed] [Google Scholar] 128. McKenzie S и Buzsáki G (2016) Механизмы гиппокампа для сегментации пространства по целям и границам в микро-, мезо- и макродинамике мозга (Buzsáki G and Christen Y, eds), стр.1–21, Springer [Google Scholar] 129. Nádasdy Z и другие. (1999) Воспроизведение и временная компрессия повторяющихся последовательностей спайков в гиппокампе. J. Neurosci. 19, 9497–9507 [Бесплатная статья PMC] [PubMed] [Google Scholar] 130. Skaggs WE и McNaughton BL (1996) Воспроизведение последовательностей возбуждения нейронов в гиппокампе крыс во время сна после пространственного опыта. Наука 271, 1870–1873 [PubMed] [Google Scholar] 131. Ли А.К. и Уилсон М.А. (2002) Память о последовательном опыте в гиппокампе во время медленного сна.Нейрон 36, 1183–1194 [PubMed] [Google Scholar]Проектирование оборудования для работы с дронами
Сводка
Наша команда предоставила комплексные услуги по проектированию новых средств дистанционного пилотирования (ДПВС) MQ-9 и вспомогательных элементов спального места элемента управления полетом (MCE) на авиабазе Шоу. Два объекта спроектированы для прикроватного комплекса MQ-9 RPA, который включает частично двухэтажный штаб и двухэтажный оперативный комплекс. Соответствующие работы на объекте включают механическое оборудование объекта, генераторное оборудование, загон для мусора, 250 парковочных мест и навес для двух мобильных наземных станций управления (MGCS).В комплексе есть внешний двор, который способствует взаимодействию персонала, обеспечивая при этом общую точку доступа с парковок. Конструкции зданий используют линии крыши для создания визуально интегрированного комплекса, обеспечивая при этом сильное присутствие военно-воздушных сил.
Этот оперативный объект, который, по прогнозам, будет укомплектован 432 сотрудниками, напрямую поддерживает истребитель в зоне ответственности (AOR), позволяя беспилотным летательным аппаратам совершать операции из местоположений в пределах Соединенных Штатов.Этот проект обеспечивает критически важное пространство для планирования миссии и будет присоединен к стационарным объектам наземной станции управления, которые используются для управления системой беспилотного воздушного вооружения в ЗО с базовой станции. Операционный центр поддерживает планирование миссий, выполнение полетов, краткие / краткие описания миссий и устройства для обучения подразделений и должен иметь резервные системы связи, питания и коммунальных услуг для обеспечения устойчивой круглосуточной работы.
Штаб-квартира спроектирована для размещения командного пункта, аудитории на 250 человек и отряда, обслуживающего административные помещения.