Клетчатка. Что это, зачем нужна, почему и как есть больше
Всемирная Организация Здравоохранения рекомендует употреблять 400 грамм (5 порций) овощей и фруктов в день, что содержит 25–30 грамм клетчатки.
Согласно статистике европейцы употребляют только 50–70% дневной нормы клетчатки. Особенно на это влияет преобладание в рационе фастфуда, полуфабрикатов, насыщенных жиров, сахара и животного белка.
В 2019 году группа ученых провела мета-анализ влияния клетчатки на человеческий организм. Результаты исследования подтвердили, что суточная норма клетчатки в размере 25–30 грамм, которую рекомендует ВОЗ — оптимальна для здоровья.
В этой статье мы расскажем о том, что такое клетчатка, в каких продуктах она содержится и как влияет на организм.
Содержание:
1. Что такое клетчатка
2. Как клетчатка поддерживает здоровье микробиоты
3. Как увеличить долю клетчатки в рационе
4. Как оценить здоровье микробиоты
Что такое клетчатка
Клетчатка — сложный углевод, или пищевое волокно, которое содержится в продуктах растительного происхождения. Она не переваривается организмом, но бактерии кишечника используют ее для выполнения ряда функций.
Она не переваривается организмом, но бактерии кишечника используют ее для выполнения ряда функций.
Существует два вида клетчатки — растворимая и нерастворимая. Практически все растения содержат оба вида, но в разных пропорциях. Оба нужны нашему организму.
Растворимая клетчатка
Photo by Melissa Di Rocco / UnsplashРастворимая клетчатка при соединении с водой приобретает вязкую гелеобразую консистенцию, что помогает пище проходить через кишечник, а также положительно влияет на некоторые показатели в организме:
Уровень сахара в крови: растворимая клетчатка замедляет всасывание макронутриентов из пищи, особенно сахаров. Тем самым она помогает контролировать уровень глюкозы в крови, что важно для людей с устойчивостью к инсулину, диабетом 2 типа или предрасположенностью к диабету.
Уровень холестерина в крови: растворимая клетчатка блокирует и снижает общий уровень поступающего с пищей холестерина, в том числе липопротеинов низкой плотности. Это помогает при заболеваниях сердца и снижает риски их развития.
Это помогает при заболеваниях сердца и снижает риски их развития.
Желчные кислоты: тело использует холестерин для производства желчных кислот, которые помогают расщеплять жиры и собирать отходы, образующиеся в процессе метаболизма. Чтобы желчь не накапливалась в организме, растворимая клетчатка блокирует ее и выводит со стулом.
Аппетит и вес: исследования показывают, что растворимая клетчатка усиливает чувство насыщения и надолго утоляют голод. Результаты исследований указывают на снижение веса и улучшение индекса массы тела у пациентов с лишним весом и ожирением.
Микробиота кишечника: растворимая клетчатка считается пребиотиком — питанием для полезных бактерий кишечника. Они, в свою очередь, расщепляют ее и производят короткоцепочечные жирные кислоты.
Тест микробиоты Атлас расскажет о способности микробиоты к расщиплению клечатки. Сделав тест, вы получите рекомендации по питанию, чтобы употреблять достаточное количество клечатки для здоровья микробиоты.

Нерастворимая клетчатка
Photo by Tom Hermans / UnsplashНерастворимая клетчатка — устойчивые к пищеварительным ферментам волокна, которые проходят через организм практически в неизменном виде. Они помогают формировать каловые массы.
Нерастворимая клетчатка стимулирует стенки кишечника на выделение слизи (муцина) и жидкости. Волокна впитывают воду, формируя каловые массы, а дополнительная слизь помогает продвигать их через кишечник и выводить из организма.
Как клетчатка поддерживает здоровье микробиоты
Необходимое количество клетчатки в рационе улучшает сразу несколько показателей организма благодаря ее воздействию на полезные и пробиотические бактерии.
Влияние пищевых волокон на укрепление иммунитета, регулярность опорожнения кишечника и уменьшение аппетита доказано клинически. Например, волокно инулин способствует впитыванию минеральных веществ, но при этом вызывает повышенное газообразование.
Волокна способствуют регулировке уровня сахара и холестерина в крови. Это снижает риск диабета 2 типа и сердечно-сосудистых заболеваний. Кроме того, взаимодействие клетчатки и микробов помогает контролировать и уменьшать хронические воспаления, которые ассоциируются со многими распространенными заболеваниями.
Это снижает риск диабета 2 типа и сердечно-сосудистых заболеваний. Кроме того, взаимодействие клетчатки и микробов помогает контролировать и уменьшать хронические воспаления, которые ассоциируются со многими распространенными заболеваниями.
| Польза | Тип клетчатки | Продукты |
|---|---|---|
| Сытость, уменьшение аппетита | Декстрин, полидекстроза | Пшеница, картофель, рис |
| Улучшение уровня сахара в крови | Фруктоолигосахариды, резистентный крахмал, пиродекстрин | Бобовые, пшеница, рожь, лук, чеснок, термически обработанный крахмал, овощные бананы, охлажденный картофель и паста |
| Уменьшение воспалений и укрепление иммунитета | Арабиногалактан, 𝛃-глюкан, фруктоолигосахариды, галактоолигосахариды, ксило-олигосахариды | Редис, морковь, груши, томаты, отруби, цельнозерновые, грибы, бобовые, пшеница, рожь, лук, чеснок, ромашка и эхинацея (не более 10 гр в день) |
| Улучшение уровня холестерина в крови | 𝛃-глюкан, целлюлоза | Отруби, цельнозерновые, грибы, большинство съедобных растений |
| Регулярность стула | 𝛃-глюкан | Отруби, цельнозерновые, грибы |
| Всасывание кальция и магния | Инулин | Цикорий, топинамбур, лук, чеснок |
Употребление клетчатки на 15–30% снижает общую смертность и летальные случаи, вызванные проблемами с сердечно-сосудистой системой.
Масляная кислота
Масляная кислота или бутират — короткоцепочечная жирная кислота, которая защищает кишечник от воспалений и поддерживает целостность его стенок.
Кишечные бактерии производят бутират при ферментации пищевых волокон. Среди таких бактерий — Faecalibacterium prausnitzii, Roseburia, Eubacterium и некоторые другие. При недостатке клетчатки производство масляной кислоты нарушается, что приводит к ослаблению иммунитета и увеличивает риски воспалений.
Бактерии-производители бутирата питаются такими видами клетчатки, как арабиноксилан, инулин, пектин, 𝛃-глюкан, полидекстроза. Они содержатся в ячмене, овсе, цикории, топинамбуре, луке, чесноке, ржи, яблоках, цитрусовых, ягодах, цельнозерновых продуктах, отрубях, грибах.
Клетчатка как пребиотик
Пребиотики — виды волокон, которые при попадании в кишечник питают бактерии и стимулируют их рост.
К таким видам клетчатки относятся бета-глюканы, галакто-, фрукто-, ксило- и арабиноолигосахариды, изомальтоза, лактулоза, олигофруктоза, инулин, устойчивый крахмал. Все они содержатся в продуктах растительного происхождения.
Все они содержатся в продуктах растительного происхождения.
Клинические исследования свидетельствуют о том, что бактерии, как и люди, очень избирательны в еде. Одни предпочитают пищевые волокна из бобовых, а другим больше по вкусу клетчатка из брокколи или овсянки.
Если вы прошли Тест Микробиоты Атлас, то могли заметить большое количество непонятных терминов в отчете «Уровень потребления пищевых волокон». Все это — пребиотики, которые присутствуют в вашем рационе.
Благодаря тестированию микробиоты мы узнаём, какие бактерии живут и преобладают в вашем кишечнике, какие виды клетчатки они предпочитают, и насколько эффективно микробиота справляется с расщеплением волокон. Эта информация нужна, чтобы подобрать персональные рекомендации по питанию.
Питание для пробиотических бактерий
Микробиота кишечника — сообщество бактерий. Чем выше их разнообразие, тем лучше показатели здоровья. Средняя и низкая представленность бактерий говорит о дисбалансе микробиоты.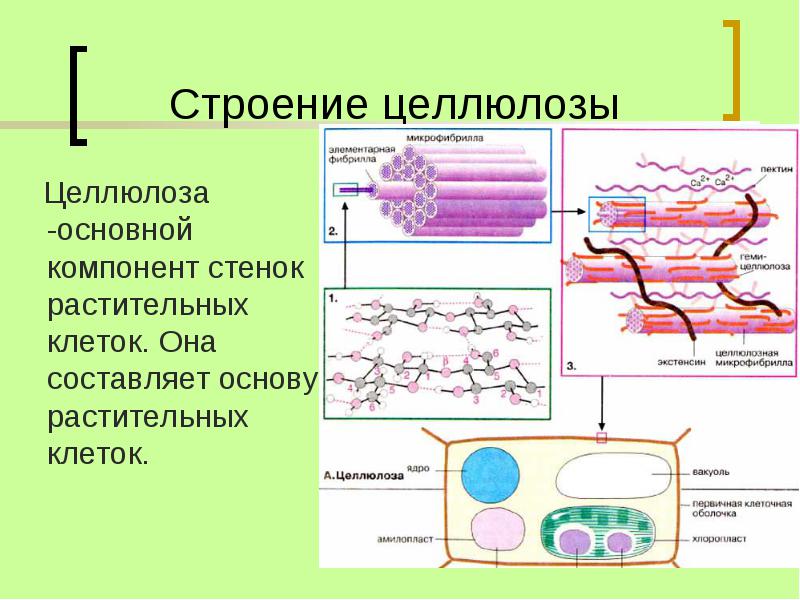 Из-за этого потенциал к защите организма от заболеваний и воспалений может быть снижен.
Из-за этого потенциал к защите организма от заболеваний и воспалений может быть снижен.
Чем больше пищевых волокон из разных источников в рационе, тем больше полезных бактерий в вашем кишечнике. Например, пробиотические бактерии Bifidobacterium производят ацетат и лактат. Их, в свою очередь, используют бактерии вида Firmicutes для производства бутирата.
Эти бактерии поддерживают кислотность кишечника, обеспечивают защиту от воспалений и патогенных организмов, способствуют укреплению иммунитета, и даже снижению уровня стресса.
Такие представители сообщества, как Akkermansia, обладают другими полезными качествами. Они присутствуют в кишечнике людей с нормальным весом, и помогают контролировать набор и поддержание здоровой массы тела.
| Эффект | Тип клетчатки | Продукты |
|---|---|---|
| Рост бифидобактерий | Арабинан, арабиноксилан, фруктоолигосахариды (FOS), галактоолигосахариды (GOS), галактоманнаны, маннанолигосахариды | Свекла, рожь, ячмень, овес, молочные продукты, цикорий, топинамбур, лук, чеснок, грибы майтаке, пекарские дрожжи |
| Рост молочнокислых бактерий | Фруктоолигосахариды, инулин, галактан, галактоманнан, пуллулан, пиродекстрин | Термически обработанный крахмал, рожь, пшеница, лук, чеснок, бобовые, молочные продукты, цикорий, топинамбур, красные водоросли, грибы майтаке |
| Рост Akkermansia | Арабиноксилан | Рожь, ячмень, овес |
Как увеличить долю клетчатки в рационе
Казалось бы, все что требуется — употреблять не менее 30 грамм клетчатки из натуральных продуктов. Однако, добавлять клетчатку следует постепенно. При резком увеличении волокон в рационе часто проявляются побочные эффекты, например, вздутие, повышенное газообразование и боли в животе.
Однако, добавлять клетчатку следует постепенно. При резком увеличении волокон в рационе часто проявляются побочные эффекты, например, вздутие, повышенное газообразование и боли в животе.
Британская ассоциация питания, образа жизни и медицины, рекомендует использовать принцип радуги: ежедневно съедать пять порций овощей и два фрукта, каждый из которых должен соответствовать одному из цветов радуги.
Отслеживать результат можно с помощью Дневника питания, который встроен в мобильное приложение Атлас.
Полуфабрикаты и фастфуд небогаты клетчаткой, и содержат много соли, сахара и жиров.
Конструктор рациона
🍛 Гарнир
Цельнозерновые: пшеница, рожь, ячмень, овес, киноа, гречка (это могут быть как зерна, так и мука).
Бобовые: фасоль, чечевица, нут.
Крахмал: картофель (приготовленный и охлажденный), зеленые бананы, коричневый рис.
🥬 Дополнительные источники клетчатки
Овощи: сырые, жареные, вареные, тушеные, на пару, сушеные.
Фрукты: предпочтительно свежие, иногда сушеные. Лучше всего на завтрак или десерт.
Зелень: свежая, сушеная.
Грибы: не забудьте и про них.
🥑 Жиры
Оливковое масло: соблюдайте меру.
Авокадо: вкусное и богатое клетчаткой, тоже важна мера.
Греческий йогурт: пробиотик, отлично подходит для завтрака или в качестве заправки.
🍋 Приправы и заправки
Семечки и орехи: отдавайте предпочтение сырым, они придадут хрустящую текстуру блюду.
Цитрусовые: их сок хорошо подойдет в качестве заправки
Травы: придают насыщенный вкус.
Специи: в зависимости от настроения.
Перец: черный, красный, смесь, сушеный или свежий
Соль: главное, не пересолить.
Пищевые дрожжи: добавляют сырный привкус и аромат. Лучше подходят для уже готовых блюд.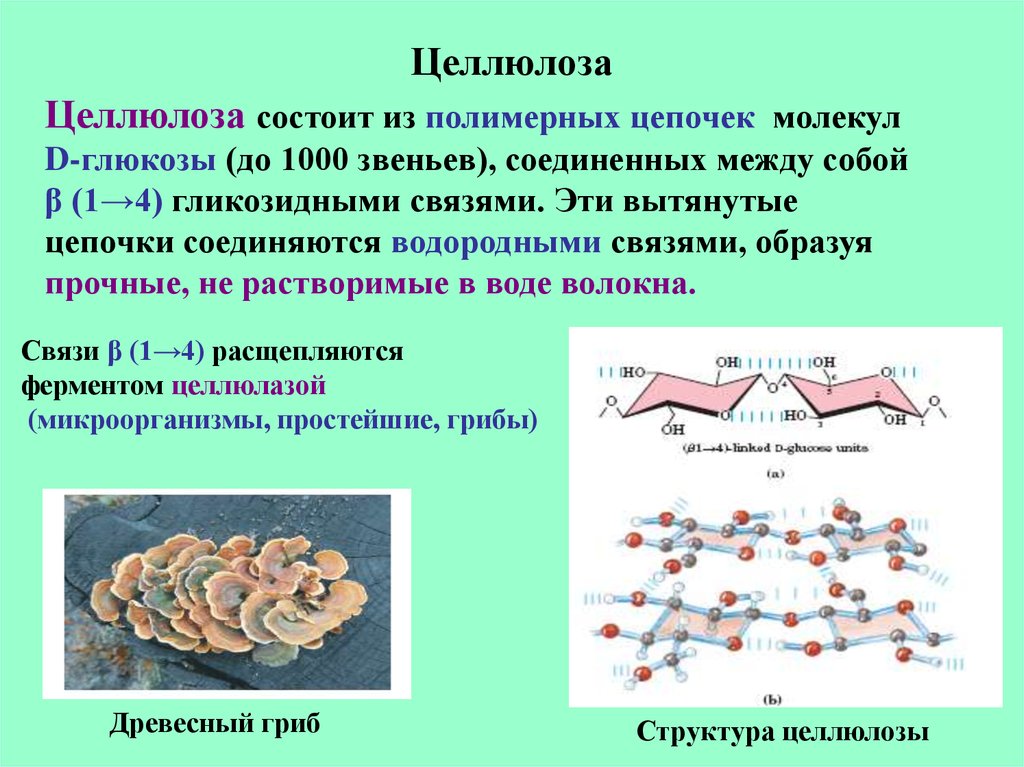
🥚 Белки
Красное мясо, морепродукты и жирная рыба: каждый не более раза в неделю.
Белое мясо и яйца: в меру.
Растительный белок: бобовые, тофу.
Стоит ли принимать клетчатку в виде БАД
Употреблять рекомендуемое количество клетчатки при обычном питании бывает нелегко. Теория о том, что в большом количестве она способствует похудению, а также доступность в виде пищевых добавок, делает выбор в пользу БАДов более привлекательным.
Результаты исследований, направленных на изучение того, как клетчатка в виде добавок влияет на снижение индекса массы тела — противоречивы. С одной стороны, некоторые БАДы на долгое время утоляют голод. Это уменьшает аппетит и способствует употреблению меньшего числа калорий.
С другой стороны, добавки содержат клетчатку в чистом виде: в отличие от овощей и фруктов они лишены витаминов и минералов. Кроме того, в процессе производства клетчатка часто теряет способность к гелеобразованию — она не создает вязкую массу, которая дает чувство сытости.
Клетчатка в виде биологически активных добавок имеет свои плюсы: она помогает дополнить количество волокон в рационе, а также используется для лечения некоторых желудочно-кишечных расстройств. Но прибегать к такому методу следует под наблюдением врача.
Как оценить здоровье микробиоты
В Тесте микробиоты мы изучаем ДНК бактерий из образца стула. Это позволяет узнать уровень разнообразия микробиоты, как бактерии справляются с синтезом масляной кислоты и витаминов, а также каких видов клетчатки вам не хватает.
В личный кабинет в раздел Питание мы загрузим отчеты по четырем показателям:
| Уровень пробиотических и других полезных бактерий | Отчет расскажет о содержании полезных бактерий в микробиоте и о том, какие продукты необходимы для поддержания их численности. |
| Уровень разнообразия микробиоты | Разнообразие микробиоты положительно влияет на здоровье пищеварения, защиту от патогенов и укрепление иммунитета. Отчет помогает понять примерное число видов бактерий, обитающих в кишечнике. |
| Уровень потребления пищевых волокон | Отчет расскажет о том, насколько микробиота справляется с расщеплением клетчатки и синтезом необходимых веществ. |
| Уровень масляной кислоты | Отчет о потенциале микробиоты синтезировать масляную кислоту — основной источник питания клеток эпителия кишечника. |
Результаты теста также включают рекомендации по питанию, основанные на вашем составе кишечных бактерий.
Другие статьи о полезных нутриентах в блоге Атлас:
- Все, что нужно знать о жирах
- Гид по углеводам
- 13 главных витаминов
- ED Jesch & TP Carr, Food Ingredients That Inhibit Cholesterol Absorption, 2017
- SV Thompson et al., Effects of isolated soluble fiber supplementation on body weight, glycemia, and insulinemia in adults with overweight and obesity: a systematic review and meta-analysis of randomized controlled trials, 2017
- J Slavin & H Green, Dietary fibre and satiety, 2007
- D Dhingra et al.
 , Dietary fibre in foods: a review, 2012
, Dietary fibre in foods: a review, 2012 - World Gastroenterology Organisation, Diet and the gut guidelines, 2018
- Andrew Reynolds et al, Carbohydrate quality and human health: a series of systematic reviews and meta-analyses,2019
- McRorie, Johnson W. Jr PhD, FACG, AGAF, FACN, Evidence-Based Approach to Fiber Supplements and Clinically Meaningful Health Benefits, Part 2 What to Look for and How to Recommend an Effective Fiber Therapy, 2015
- Max H Pittler, Edzard Ernst, Dietary supplements for body-weight reduction: a systematic review, 2004
 , Dietary fibre in foods: a review, 2012
, Dietary fibre in foods: a review, 2012Wsn180211 – DW – 01.09.2011
Фото: picture-alliance/dpa
1 сентября 2011 г.
Проблема биотоплива была бы давно решена, если бы дрожжи могли расщеплять целлюлозу. Поскольку это не так, ученые вынуждены искать альтернативные технологии. Большие надежды они возлагают на синтетическую биологию.
https://p.dw.com/p/10KDV
Реклама
У всех высших растений на нашей планете оболочки клеток состоят главным образом из целлюлозы. В химическом отношении целлюлоза, она же клетчатка, представляет собой биополимер, высокомолекулярный углевод, полисахарид, молекулы которого образованы длинными, соединенными друг с другом множеством водородных связей линейными цепочками из сотен или даже тысяч остатков глюкозы. Целлюлоза буквально напрашивается в качестве сырья для биотоплива, поскольку сельскохозяйственное производство дает из года в год сотни тысяч, а то и миллионы тонн растительных отходов, которые состоят практически из чистой клетчатки. Проблема лишь в том, что целлюлоза плохо поддается расщеплению.
В химическом отношении целлюлоза, она же клетчатка, представляет собой биополимер, высокомолекулярный углевод, полисахарид, молекулы которого образованы длинными, соединенными друг с другом множеством водородных связей линейными цепочками из сотен или даже тысяч остатков глюкозы. Целлюлоза буквально напрашивается в качестве сырья для биотоплива, поскольку сельскохозяйственное производство дает из года в год сотни тысяч, а то и миллионы тонн растительных отходов, которые состоят практически из чистой клетчатки. Проблема лишь в том, что целлюлоза плохо поддается расщеплению.
Вильфрид Вебер (Wilfried Weber), профессор Центра по изучению биологических сигнальных процессов (BIOSS) при Фрайбургском университете, говорит: «Если вы собираетесь производить биотопливо не из продовольственного сырья вроде кукурузы, пальмового масла и так далее, то оптимальной альтернативой представляются трава и солома как отход сельского хозяйства, а также древесина. Но с этими видами сырья связана одна проблема: главный энергоноситель в них — целлюлоза.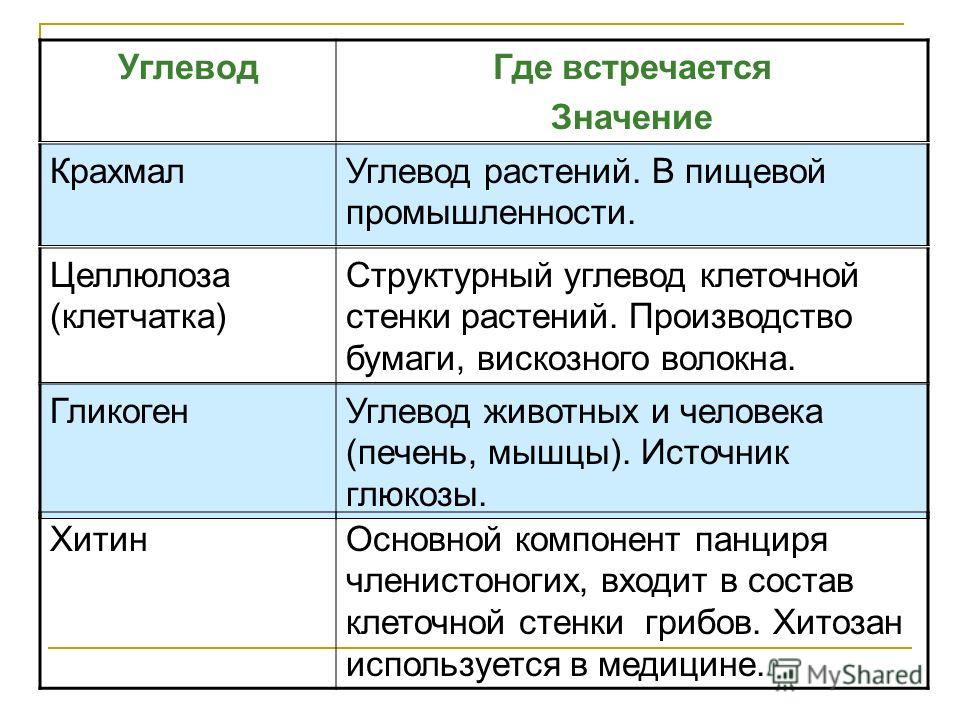 А переработать целлюлозу в жидкое топливо, пригодное, скажем, для заправки автомобиля, очень непросто».
А переработать целлюлозу в жидкое топливо, пригодное, скажем, для заправки автомобиля, очень непросто».
Термиты как альтернатива дрожжам
Во всяком случае, дрожжи, легко осуществляющие спиртовое брожение простых сахаров, с целлюлозой не справляются. Поэтому ученые разных стран активно ищут альтернативные процессы и технологии. Особенно активно работают в этом направлении американцы и немцы. Американский микробиолог, профессор Гари Строубел (Gary Strobel) из университета штата Монтана в Боузмене — изучает обнаруженный в Чили симбиотический гриб Gliocladium roseum, способный расщеплять целлюлозу с образованием этана и других летучих углеводородов; другой — профессор Джаред Ледбеттер (Jared R. Leadbetter) из Калифорнийского технологического института в Пасадене — исследует кишечную микрофлору обитающих в Коста-Рике термитов рода Nasutitermes, способных за считанные месяцы полностью переварить деревянный дом вместе с мебелью.
А специалисты Объединенного института геномных исследований в Уолнат-Крике, штат Калифорния, обратились к крупному рогатому скоту. Ведь хотя жвачные животные — так же, как и люди — не обладают собственными ферментами, способными расщеплять целлюлозу, они тем не менее прекрасно обходятся в качестве корма травой и сеном, то есть практически чистой клетчаткой, и умудряются усваивать ее так, что она вполне удовлетворяет их энергетические потребности.
Ведь хотя жвачные животные — так же, как и люди — не обладают собственными ферментами, способными расщеплять целлюлозу, они тем не менее прекрасно обходятся в качестве корма травой и сеном, то есть практически чистой клетчаткой, и умудряются усваивать ее так, что она вполне удовлетворяет их энергетические потребности.
Метагеномный анализ коровьего желудка
Очевидно, все дело тут в микрофлоре пищеварительного тракта. Именно это и побудило калифорнийских ученых произвести так называемый метагеномный анализ микроорганизмов, населяющих коровий желудок, то есть секвенировать все имеющиеся там нуклеиновые кислоты в надежде обнаружить гены, причастные к расщеплению клетчатки. В общей сложности исследователи идентифицировали в коровьем желудке почти 30 тысяч биомолекул. Теперь ученые надеются, что процессы, происходящие в желудке коровы, удастся воспроизвести в искусственных условиях — сначала в лаборатории, а потом и в крупных промышленных установках по производству биоэтанола.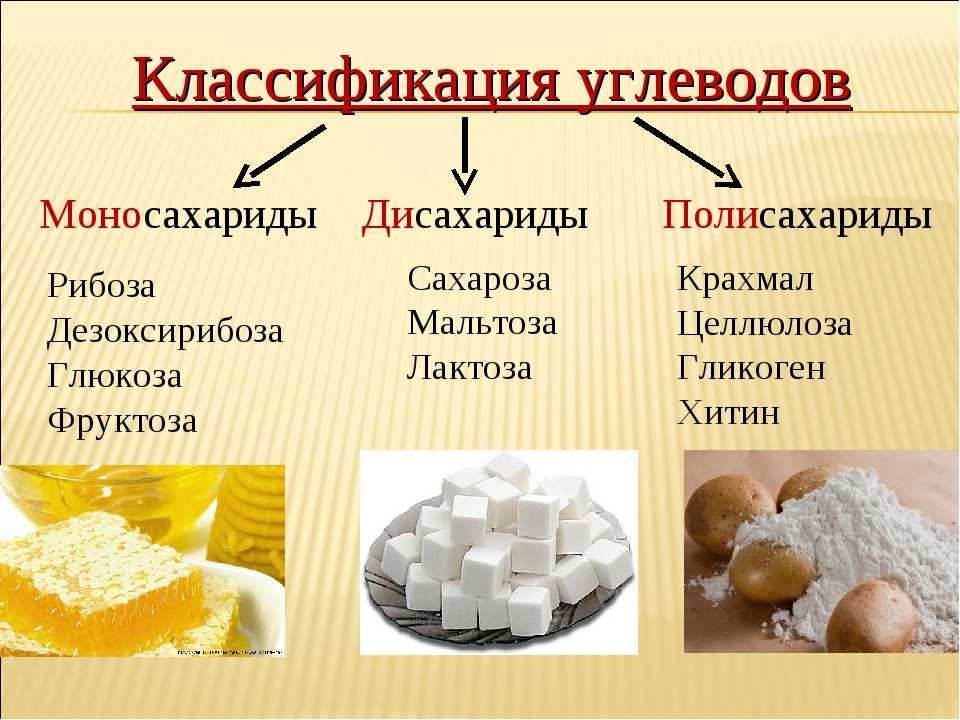 Во всяком случае, это отвечало бы представлениям так называемой синтетической биологии, которая видит свою задачу в том, чтобы по-новому скомбинировать отдельные природные элементы.
Во всяком случае, это отвечало бы представлениям так называемой синтетической биологии, которая видит свою задачу в том, чтобы по-новому скомбинировать отдельные природные элементы.
Журналисты любят сравнивать такой подход со сборкой самых разных изделий и сооружений из стандартных кирпичиков конструктора Lego. Но на самом деле не все так просто, — говорит Вильфрид Вебер: «До такой легоподобной биологии нам пока, к сожалению, еще очень далеко. Это связано с тем, что биологические компоненты гораздо хуже поддаются стандартизации или переносу из одной среды в другую, чем кирпичики лего или, скажем, электронные компоненты».
Легоподобные биокомпоненты
Ключевую роль в современной синтетической биологии играет база данных в Массачусетском технологическом институте в Кеймбридже близ Бостона. Эта база содержит информацию обо всех имеющихся в наличии биокомпонентах. Их можно заказать, как книги или одежду по каталогу посылторга. Впрочем, и тут есть проблемы. Вильфрид Вебер: «На сегодняшний день этот каталог MIT содержит более 3,5 тысяч биологических компонентов — в основном генов. Это очень ценный ресурс, позволяющий получить все необходимые фрагменты ДНК. К сожалению, описание этих фрагментов часто оставляет желать лучшего. Нам уже не раз доводилось испытывать разочарование, когда мы заказывали там определенные компоненты и получали совсем не то, что указывалось в описании. То есть система контроля качества там пока не на высоте». Но это, конечно, ни в коей мере не ставит под сомнение перспективы синтетической биологии.
Это очень ценный ресурс, позволяющий получить все необходимые фрагменты ДНК. К сожалению, описание этих фрагментов часто оставляет желать лучшего. Нам уже не раз доводилось испытывать разочарование, когда мы заказывали там определенные компоненты и получали совсем не то, что указывалось в описании. То есть система контроля качества там пока не на высоте». Но это, конечно, ни в коей мере не ставит под сомнение перспективы синтетической биологии.
Автор: Владимир Фрадкин
Редактор: Ефим Шуман
Реклама
Пропустить раздел Еще по темеЕще по теме
Показать еще
Пропустить раздел Топ-тема1 стр. из 3
Пропустить раздел Другие публикации DWНа главную страницу
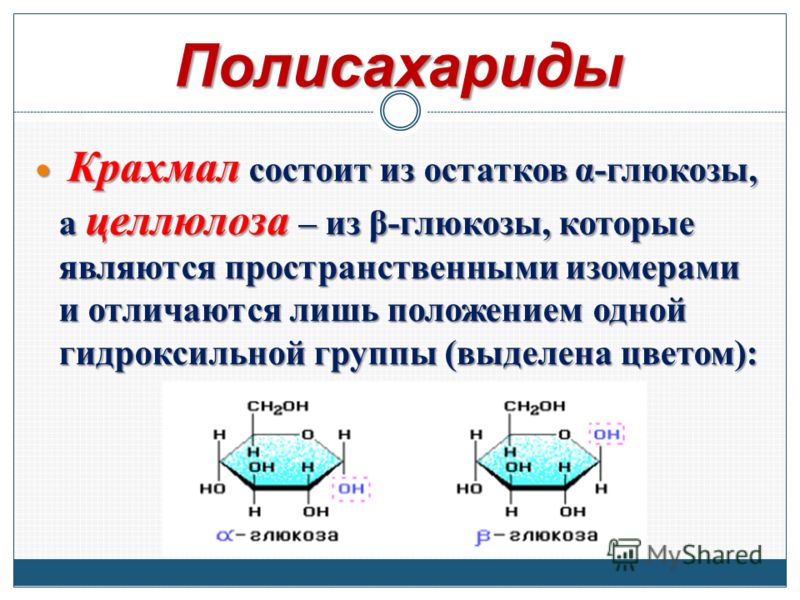
Структура и функция углеводов
Результаты обучения
- Различать моносахариды, дисахариды и полисахариды
- Определите несколько основных функций углеводов
Большинство людей знакомы с углеводами, одним из типов макромолекул, особенно когда речь идет о том, что мы едим.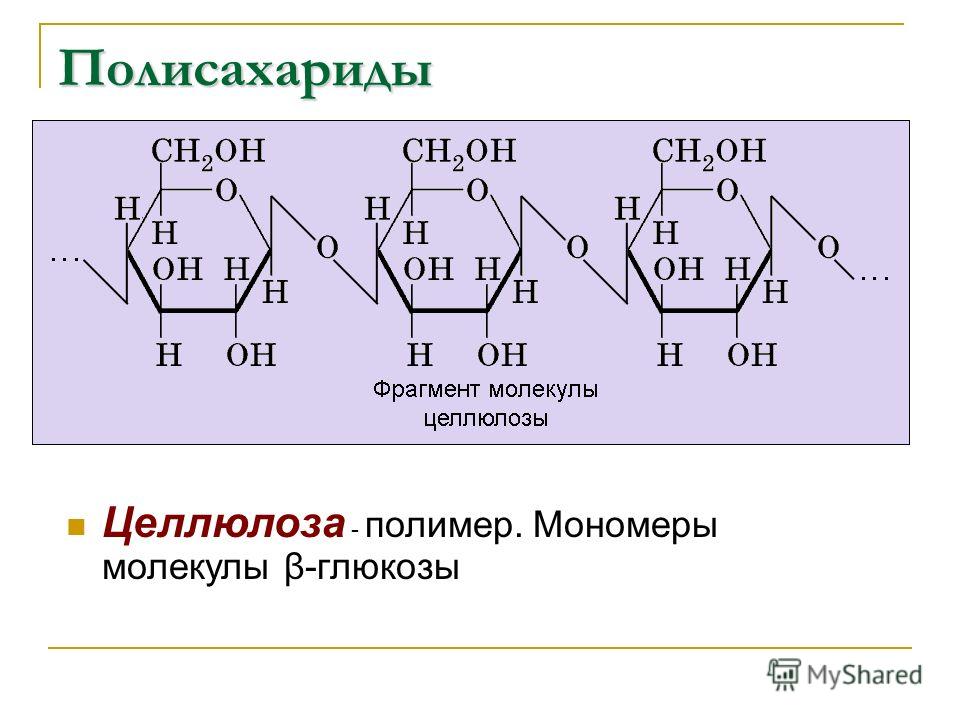 Чтобы похудеть, некоторые люди придерживаются «низкоуглеводной» диеты. Спортсмены, напротив, часто «загружаются углеводами» перед важными соревнованиями, чтобы убедиться, что у них достаточно энергии для соревнований на высоком уровне. Углеводы, по сути, являются неотъемлемой частью нашего рациона; злаки, фрукты и овощи являются естественными источниками углеводов. Углеводы обеспечивают организм энергией, в частности, за счет глюкозы, простого сахара, который является компонентом крахмала и ингредиентом многих основных продуктов питания. Углеводы также выполняют другие важные функции в организме человека, животных и растений.
Чтобы похудеть, некоторые люди придерживаются «низкоуглеводной» диеты. Спортсмены, напротив, часто «загружаются углеводами» перед важными соревнованиями, чтобы убедиться, что у них достаточно энергии для соревнований на высоком уровне. Углеводы, по сути, являются неотъемлемой частью нашего рациона; злаки, фрукты и овощи являются естественными источниками углеводов. Углеводы обеспечивают организм энергией, в частности, за счет глюкозы, простого сахара, который является компонентом крахмала и ингредиентом многих основных продуктов питания. Углеводы также выполняют другие важные функции в организме человека, животных и растений.
Молекулярные структуры
Углеводы могут быть представлены формулой (CH 2 O) n , где n — число атомов углерода в молекуле. Другими словами, отношение углерода к водороду и кислороду составляет 1:2:1 в молекулах углеводов. Эта формула также объясняет происхождение термина «углевод»: компонентами являются углерод («карбо») и компоненты воды (отсюда «гидрат»). Углеводы представляют собой полимеры, построенные из сахаров, которые являются их мономерами. Углеводы делятся на три подтипа: моносахариды, дисахариды и полисахариды.
Углеводы представляют собой полимеры, построенные из сахаров, которые являются их мономерами. Углеводы делятся на три подтипа: моносахариды, дисахариды и полисахариды.
Моносахариды
Моносахариды ( моно – = «один»; сахар – = «сладкий») представляют собой простые сахара, наиболее распространенным из которых является глюкоза. В моносахаридах число атомов углерода обычно колеблется от трех до семи. Большинство названий моносахаридов заканчиваются суффиксом — ose . Если в сахаре есть альдегидная группа (функциональная группа со структурой R-CHO), он известен как альдоза, а если он имеет кетоновую группу (функциональная группа со структурой RC(=O)R′), он известен как кетоза. В зависимости от количества атомов углерода в сахаре они также могут быть известны как триозы (три атома углерода), пентозы (пять атомов углерода) и/или гексозы (шесть атомов углерода). См. рисунок 1 для иллюстрации моносахаридов.
Рисунок 1. Моносахариды классифицируются на основе положения их карбонильной группы и количества атомов углерода в основной цепи. Альдозы имеют карбонильную группу (обозначена зеленым цветом) на конце углеродной цепи, а кетозы имеют карбонильную группу в середине углеродной цепи. Триозы, пентозы и гексозы имеют соответственно три, пять и шесть углеродных цепей.
Альдозы имеют карбонильную группу (обозначена зеленым цветом) на конце углеродной цепи, а кетозы имеют карбонильную группу в середине углеродной цепи. Триозы, пентозы и гексозы имеют соответственно три, пять и шесть углеродных цепей.
Химическая формула глюкозы C 6 H 12 O 6 . Для человека глюкоза является важным источником энергии. Во время клеточного дыхания энергия высвобождается из глюкозы, и эта энергия используется для производства аденозинтрифосфата (АТФ). Растения синтезируют глюкозу, используя углекислый газ и воду, а глюкоза, в свою очередь, используется для удовлетворения энергетических потребностей растения. Избыток глюкозы часто откладывается в виде крахмала, который катаболизируется (расщепление более крупных молекул клетками) людьми и другими животными, питающимися растениями.
Галактоза и фруктоза являются другими распространенными моносахаридами — галактоза содержится в молочных сахарах, а фруктоза — во фруктовых сахарах. Хотя глюкоза, галактоза и фруктоза имеют одинаковую химическую формулу (C 6 H 12 O 6 ), они различаются структурно и химически (и известны как изомеры) из-за различного расположения функциональных групп вокруг асимметричный углерод; все эти моносахариды имеют более одного асимметрического углерода.
Хотя глюкоза, галактоза и фруктоза имеют одинаковую химическую формулу (C 6 H 12 O 6 ), они различаются структурно и химически (и известны как изомеры) из-за различного расположения функциональных групп вокруг асимметричный углерод; все эти моносахариды имеют более одного асимметрического углерода.
Моносахариды могут существовать в виде линейных цепочек или кольцеобразных молекул; в водных растворах они обычно находятся в кольцевых формах.
Дисахариды
Дисахариды ( di – = «два») образуются, когда два моносахарида подвергаются реакции дегидратации (также известной как реакция конденсации или синтез дегидратации). В ходе этого процесса гидроксильная группа одного моносахарида соединяется с водородом другого моносахарида, высвобождая молекулу воды и образуя ковалентную связь (рис. 2).
Рисунок 2. Сахароза получается в результате химической реакции между двумя простыми сахарами, называемыми глюкозой и фруктозой.
Общие дисахариды включают лактозу, мальтозу и сахарозу. Лактоза – это дисахарид, состоящий из мономеров глюкозы и галактозы. В природе содержится в молоке. Мальтоза, или солодовый сахар, представляет собой дисахарид, образующийся в результате реакции дегидратации между двумя молекулами глюкозы. Наиболее распространенным дисахаридом является сахароза или столовый сахар, который состоит из мономеров глюкозы и фруктозы.
Полисахариды
Длинная цепь моносахаридов, связанных ковалентными связями, известна как полисахарид ( поли – = «много»). Цепь может быть разветвленной или неразветвленной, и она может содержать различные типы моносахаридов. Полисахариды могут быть очень большими молекулами. Крахмал, гликоген, целлюлоза и хитин являются примерами полисахаридов.
Крахмал представляет собой запасенную форму сахаров в растениях и состоит из амилозы и амилопектина (оба полимеры глюкозы). Растения способны синтезировать глюкозу, а избыток глюкозы откладывается в виде крахмала в различных частях растения, включая корни и семена. Крахмал, потребляемый животными, расщепляется на более мелкие молекулы, такие как глюкоза. Затем клетки могут поглощать глюкозу.
Крахмал, потребляемый животными, расщепляется на более мелкие молекулы, такие как глюкоза. Затем клетки могут поглощать глюкозу.
Рисунок 3. Амилоза и амилопектин представляют собой две разные формы крахмала. Гликоген представляет собой запасную форму глюкозы у человека и других позвоночных и состоит из мономеров глюкозы.
Гликоген представляет собой запасную форму глюкозы у людей и других позвоночных и состоит из мономеров глюкозы. Гликоген является животным эквивалентом крахмала и представляет собой сильно разветвленную молекулу, обычно хранящуюся в клетках печени и мышц. Всякий раз, когда уровень глюкозы снижается, гликоген расщепляется с выделением глюкозы.
Целлюлоза является одним из самых распространенных природных биополимеров. Клеточные стенки растений в основном состоят из целлюлозы, которая обеспечивает структурную поддержку клетки. Древесина и бумага в основном целлюлозные по своей природе. Целлюлоза состоит из мономеров глюкозы, которые связаны связями между определенными атомами углерода в молекуле глюкозы.
Все остальные мономеры глюкозы в целлюлозе перевернуты и плотно упакованы в виде вытянутых длинных цепей (см. рис. 4). Это придает целлюлозе жесткость и высокую прочность на растяжение, что так важно для растительных клеток. Целлюлоза, проходящая через нашу пищеварительную систему, называется пищевыми волокнами. В то время как глюкозо-глюкозные связи в целлюлозе не могут быть разрушены пищеварительными ферментами человека, травоядные животные, такие как коровы, буйволы и лошади, способны переваривать траву, богатую целлюлозой, и использовать ее в качестве источника пищи. У этих животных некоторые виды бактерий обитают в рубце (часть пищеварительной системы травоядных) и выделяют фермент целлюлазу. Аппендикс также содержит бактерии, расщепляющие целлюлозу, что придает ей важную роль в пищеварительной системе жвачных животных. Целлюлазы могут расщеплять целлюлозу на мономеры глюкозы, которые могут использоваться животным в качестве источника энергии.
Рисунок 4. В целлюлозе мономеры глюкозы связаны в неразветвленные цепи β 1-4 гликозидными связями. Из-за способа соединения субъединиц глюкозы каждый мономер глюкозы переворачивается по отношению к следующему, что приводит к линейной волокнистой структуре.
Из-за способа соединения субъединиц глюкозы каждый мономер глюкозы переворачивается по отношению к следующему, что приводит к линейной волокнистой структуре.
Рисунок 5. Насекомые имеют твердый внешний скелет, состоящий из хитина, разновидности полисахарида.
Углеводы выполняют другие функции у разных животных. Членистоногие, такие как насекомые, пауки и крабы, имеют внешний скелет, называемый экзоскелетом, который защищает их внутренние части тела. Этот экзоскелет состоит из биологической макромолекулы хитина, азотсодержащего углевода. Он состоит из повторяющихся звеньев модифицированного сахара, содержащего азот.
Зарегистрированный диетолог
Рис. 6. Зарегистрированный диетолог-диетолог (RDN) Шеф-повар Бренда Томпсон работает с персоналом общественного питания над составлением своего рецепта буррито на завтрак во время тестирования вкуса, разработанного шеф-поваром в школе в Айдахо. Благодаря гранту Team Nutrition Министерства сельского хозяйства США (USDA) шеф-повар RDN Бренда Томпсон разработала рецепты для кулинарной книги Chef Designed School Lunch.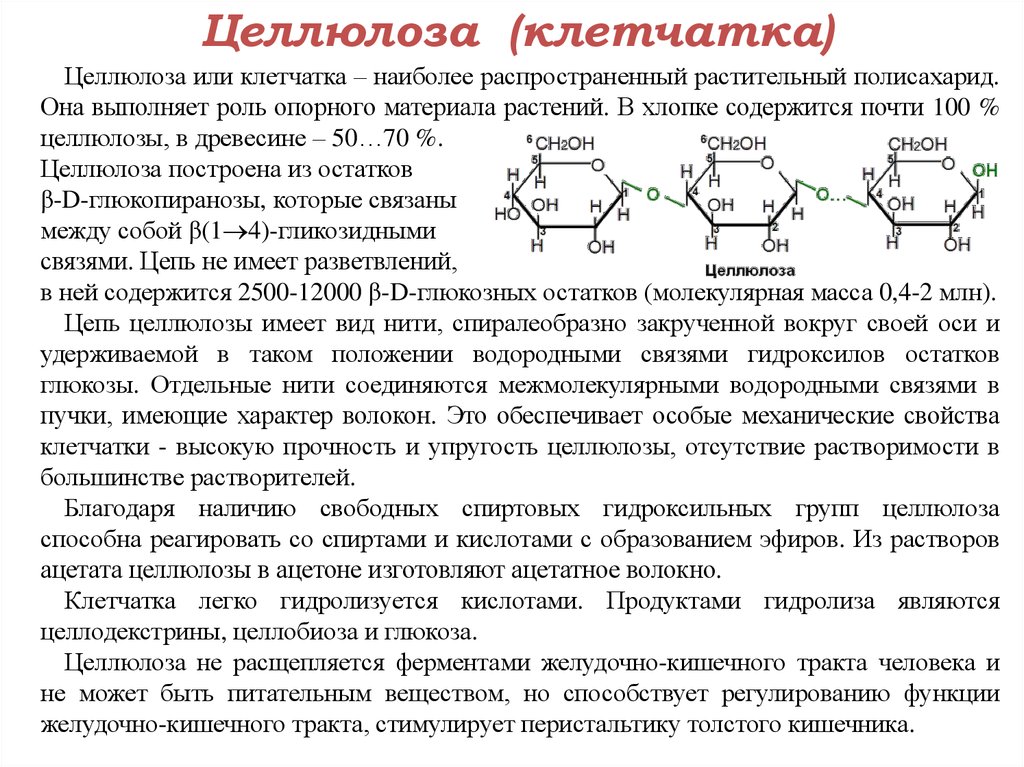
Ожирение является проблемой здравоохранения во всем мире, и многие заболевания, такие как диабет и болезни сердца, становятся все более распространенными из-за ожирения. Это одна из причин, почему все чаще обращаются за советом к зарегистрированным диетологам. Зарегистрированные диетологи помогают планировать программы питания и питания для людей в различных условиях. Они часто работают с пациентами в медицинских учреждениях, разрабатывая планы питания для профилактики и лечения заболеваний. Например, диетологи могут научить пациента с диабетом, как управлять уровнем сахара в крови, употребляя углеводы правильного типа и количества. Диетологи также могут работать в домах престарелых, школах и частных практиках.
Чтобы стать зарегистрированным диетологом, необходимо получить как минимум степень бакалавра в области диетологии, питания, пищевых технологий или в смежных областях. Кроме того, зарегистрированные диетологи должны пройти стажировку под наблюдением и сдать национальный экзамен. Те, кто занимается диетологией, проходят курсы по питанию, химии, биохимии, биологии, микробиологии и физиологии человека. Диетологи должны стать экспертами в области химии и функций пищи (белков, углеводов и жиров).
Те, кто занимается диетологией, проходят курсы по питанию, химии, биохимии, биологии, микробиологии и физиологии человека. Диетологи должны стать экспертами в области химии и функций пищи (белков, углеводов и жиров).
Резюме: структура и функция углеводов
Углеводы представляют собой группу макромолекул, которые являются жизненно важным источником энергии для клетки и обеспечивают структурную поддержку клеток растений, грибов и всех членистоногих, включая омаров, крабов, креветок, насекомые и пауки. Углеводы классифицируются как моносахариды, дисахариды и полисахариды в зависимости от количества мономеров в молекуле. Моносахариды связаны гликозидными связями, которые образуются в результате реакций дегидратации, образуя дисахариды и полисахариды с отщеплением молекулы воды на каждую образовавшуюся связь. Глюкоза, галактоза и фруктоза являются распространенными моносахаридами, тогда как распространенные дисахариды включают лактозу, мальтозу и сахарозу. Крахмал и гликоген, примеры полисахаридов, являются запасными формами глюкозы в растениях и животных соответственно. Длинные полисахаридные цепи могут быть разветвленными или неразветвленными. Целлюлоза является примером неразветвленного полисахарида, тогда как амилопектин, входящий в состав крахмала, представляет собой сильно разветвленную молекулу. Хранение глюкозы в виде полимеров, подобных крахмалу гликогена, делает ее несколько менее доступной для метаболизма; однако это предотвращает его утечку из клетки или создание высокого осмотического давления, которое может вызвать чрезмерное поглощение воды клеткой.
Длинные полисахаридные цепи могут быть разветвленными или неразветвленными. Целлюлоза является примером неразветвленного полисахарида, тогда как амилопектин, входящий в состав крахмала, представляет собой сильно разветвленную молекулу. Хранение глюкозы в виде полимеров, подобных крахмалу гликогена, делает ее несколько менее доступной для метаболизма; однако это предотвращает его утечку из клетки или создание высокого осмотического давления, которое может вызвать чрезмерное поглощение воды клеткой.
Попробуйте
Внесите свой вклад!
У вас есть идеи по улучшению этого контента? Мы будем признательны за ваш вклад.
Улучшить эту страницуПодробнее
Brittle Culm1, COBRA-подобный белок, участвует в сборке целлюлозы посредством связывания микрофибрилл целлюлозы
1. Дарли К.П., Форрестер А.М., МакКуин-Мейсон С.Дж. (2001) Молекулярная основа растяжения клеточной стенки растений. Растение Мол Биол 47: 179–195. [PubMed] [Google Scholar]
2. Ариоли Т., Пэн Л., Бетцнер А.С., Берн Дж., Виттке В. и соавт. (1998) Молекулярный анализ биосинтеза целлюлозы у арабидопсиса. Наука
279: 717–720. [PubMed] [Google Scholar]
Ариоли Т., Пэн Л., Бетцнер А.С., Берн Дж., Виттке В. и соавт. (1998) Молекулярный анализ биосинтеза целлюлозы у арабидопсиса. Наука
279: 717–720. [PubMed] [Google Scholar]
3. Schindelman G, Morikami A, Jung J, Baskin TI, Carpita NC, et al. (2001) COBRA кодирует предполагаемый GPI-заякоренный белок, который локализован полярно и необходим для направленной экспансии клеток у арабидопсиса. Гены и Дев 15: 1115–1127. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Desprez T, Juraniec M, Crowell EF, Jouy H, Pochylova Z, et al. (2007) Организация комплексов синтазы целлюлозы, участвующих в синтезе первичной клеточной стенки у Arabidopsis thaliana . Proc Natl Acad Sci USA 104: 15572–15577. [Бесплатная статья PMC] [PubMed] [Google Scholar]
6. Taylor NG, Laurie S, Turner SR (2000) Множественные каталитические субъединицы синтазы целлюлозы необходимы для синтеза целлюлозы у арабидопсиса. Растительная клетка 12: 2529–2540. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Taylor NG, Howells RM, Huttly AK, Vickers K, Turner SR (2003)Взаимодействия между тремя различными белками CesA, необходимыми для синтеза целлюлозы. Proc Natl Acad Sci USA
100: 1450–1455. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Taylor NG, Howells RM, Huttly AK, Vickers K, Turner SR (2003)Взаимодействия между тремя различными белками CesA, необходимыми для синтеза целлюлозы. Proc Natl Acad Sci USA
100: 1450–1455. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Li Y, Qian Q, Zhou Y, Yan M, Sun L и др. (2003) BRITTLE CULM1 , который кодирует белок типа COBRA, влияет на механические свойства растений риса. Растительная клетка 15: 2020–2031. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Чжун Р., Пена М.Дж., Чжоу Г.К., Нэрн С.Дж., Вуд-Джонс А. и др. (2005) Arabidopsis Fragile Fiber8 , которое кодирует предполагаемую глюкуронилтрансферазу, необходимо для нормального синтеза вторичной стенки. Растительная клетка 17: 3390–3408. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10.
Шиманска-Чаргот М., Цибульска Дж., Здунек А. (2011)Определение структурных различий в целлюлозе яблока и материалах бактериальных клеточных стенок с помощью рамановской и ИК-спектроскопии. Датчики (Базель)
11: 5543–5560. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Датчики (Базель)
11: 5543–5560. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Saxena IM, Brown RM Jr (2005)Биосинтез целлюлозы: современные взгляды и развивающиеся концепции. Энн Бот 96: 9–21. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Томас Л.Х., Форсайт В.Т., Стуркова А., Кеннеди С.Дж., Мэй Р.П. и др. (2013) Структура микрофибрилл целлюлозы в первичных клеточных стенках колленхимы. Завод Физиол 161: 465–476. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Фернандес А.Н., Томас Л.Х., Альтанер К.М., Кэллоу П., Форсайт В.Т. и соавт. (2011)Наноструктура микрофибрилл целлюлозы в древесине ели. Proc Natl Acad Sci USA 108: 1195–1203. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Джарвис М. (2003) Химия: целлюлоза складывается. Природа 426: 611–612. [PubMed] [Google Scholar]
15.
Дональдсон Л. (2007) Агрегаты микрофибрилл целлюлозы и изменение их размера в зависимости от типа клеточной стенки. Научные технологии по дереву
41: 443–460. [Академия Google]
[Академия Google]
16. Андерсон К.Т., Кэрролл А., Ахметова Л., Сомервилль С. (2010)Визуализация переориентации целлюлозы в реальном времени во время расширения клеточной стенки в корнях арабидопсиса. Завод Физиол 152: 787–796. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Somerville C (2006)Синтез целлюлозы в высших растениях. Annu Rev Cell Dev Биол 22: 53–78. [PubMed] [Google Scholar]
18. Нишияма Ю., Ланган П., Чанзи Х. (2002) Кристаллическая структура и система водородных связей в целлюлозе I β из синхротронного рентгеновского излучения и нейтронной волоконной дифракции. J Am Chem Soc 124: 9074–9082. [PubMed] [Google Scholar]
19. Нишияма Ю., Сугияма Дж., Чанзи Х., Ланган П. (2003) Кристаллическая структура и система водородных связей в целлюлозе I α по данным синхротронного рентгеновского излучения и дифракции нейтронного волокна. J Am Chem Soc 125: 14300–14306. [PubMed] [Google Scholar]
20.
Харрис Д.М., Корбин К. , Ван Т., Гутьеррес Р., Бертоло А.Л. и др. (2012) Кристалличность микрофибрилл целлюлозы снижается за счет мутации остатков С-концевой трансмембранной области CESA1 A903V и CESA3 T942I синтазы целлюлозы. Proc Natl Acad Sci USA
109: 4098–4103. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Ван Т., Гутьеррес Р., Бертоло А.Л. и др. (2012) Кристалличность микрофибрилл целлюлозы снижается за счет мутации остатков С-концевой трансмембранной области CESA1 A903V и CESA3 T942I синтазы целлюлозы. Proc Natl Acad Sci USA
109: 4098–4103. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Vietor RJ, Newman RH, Ha MA, Apperley DC, Jarvis MC (2002)Конформационные особенности целлюлозы на поверхности кристаллов высших растений. Завод J 30: 721–731. [PubMed] [Google Scholar]
22. Николь Ф., Хис И., Жоно А., Вернхеттес С., Канут Х. и др. (1998) Связанная с плазматической мембраной предполагаемая эндо-1,4-β-D-глюканаза необходима для нормальной сборки стенок и удлинения клеток у арабидопсиса. Эмбо Дж 17: 5563–5576. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23.
Takahashi J, Rudsander UJ, Hedenstrom M, Banasiak A, Harholt J, et al. (2009) KORRIGAN1 и его гомолог осины PttCel9A1 снижают кристалличность целлюлозы в стеблях арабидопсиса. Физиол растительной клетки
50: 1099–1115. [PubMed] [Google Scholar]
Физиол растительной клетки
50: 1099–1115. [PubMed] [Google Scholar]
24. Санчес-Родригес С., Бауэр С., Хемати К., Сакс Ф., Ибанез А.Б. и соавт. (2012) Chitinase-like1/POM-POM1 и его гомолог CTL2 представляют собой взаимодействующие с глюканом белки, важные для биосинтеза целлюлозы у арабидопсиса. Растительная клетка 24: 589–607. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Haigler CH, Brown RM Jr, Benziman M (1980) Calcofluor white ST изменяет сборку микрофибрилл целлюлозы in vivo. Наука 210: 903–906. [PubMed] [Google Scholar]
26. Fujita M, Himmelspach R, Hocart CH, Williamson RE, Mansfield SD, et al. (2011) Кортикальные микротрубочки оптимизируют кристалличность клеточной стенки, чтобы стимулировать однонаправленный рост арабидопсиса. Завод J 66: 915–928. [PubMed] [Google Scholar]
27.
Уитни СЕК, Бригам Дж. Э., Дарк А. Х., Рейд Дж. С. Г., Гидли М. Дж. (1995) Сборка сетей целлюлоза/ксилоглюкан in vitro: ультраструктурные и молекулярные аспекты. Завод J
8: 491–504. [Google Академия]
Завод J
8: 491–504. [Google Академия]
28. Лай-Ки-Хим Дж., Чанзи Х., Мюллер М., Путо Дж.Л., Имаи Т. и др. (2002) Микрофибриллы целлюлозы in vitro и in vivo из синтаз первичной стенки растений: структурные различия. J Биол Хим 277: 36931–36939. [PubMed] [Google Scholar]
29. Рудье Ф., Фернандес А.Г., Фуджита М., Химмельспах Р., Борнер Г.Х. и др. (2005) COBRA, внеклеточный гликозил-фосфатидил-инозитол-заякоренный белок арабидопсиса, специфически контролирует сильно анизотропное расширение за счет своего участия в ориентации микрофибрилл целлюлозы. Растительная клетка 17: 1749–1763. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Roudier F, Schindelman G, DeSalle R, Benfey PN (2002)Семейство COBRA предполагаемых GPI-заякоренных белков у арабидопсиса. Новое товарищество в расширении. Завод Физиол 130: 538–548. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31.
Брэди С.М., Сонг С., Дугга К.С., Рафальский Дж.А., Бенфей П.Н. (2007) Сочетание экспрессии и сравнительного эволюционного анализа. Семейство генов COBRA . Завод Физиол
143: 172–187. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Семейство генов COBRA . Завод Физиол
143: 172–187. [Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Браун Д.М., Зиф Л.А., Эллис Дж., Гудакр Р., Тернер С.Р. (2005)Идентификация новых генов у арабидопсиса, участвующих в формировании вторичной клеточной стенки, с использованием профиля экспрессии и обратной генетики. Растительная клетка 17: 2281–2295. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Сато К., Судзуки Р., Нисикубо Н., Такэноути С., Ито С. и др. (2010)Выделение нового мутанта архитектуры клеточной стенки риса с дефектным ортологом BC1 Arabidopsis COBL4, необходимым для регулируемого отложения компонентов вторичной клеточной стенки. Планта 232: 257–270. [PubMed] [Академия Google]
34.
Sindhu A, Langewisch T, Olek A, Multani DS, McCann MC, et al. (2007) Кукуруза Brittle stalk2 кодирует COBRA-подобный белок, экспрессируемый в раннем развитии органов, но необходимый для гибкости тканей в зрелом возрасте. Завод Физиол
145: 1444–1459. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Jones MA, Raymond MJ, Smirnoff N (2006) Анализ транскриптома морфогенеза корневых волосков выявил молекулярную идентичность шести генов, играющих роль в развитии корневых волосков у Arabidopsis. Завод J 45: 83–100. [PubMed] [Академия Google]
36. Hochholdinger F, Wen TJ, Zimmermann R, Chimot-Marolle P, da Costa e Silva O, et al. (2008) Кукуруза ( Zea mays L.) без корневых волосков 3 ген кодирует предполагаемый GPI-заякоренный, специфичный для однодольных, COBRA-подобный белок, который значительно влияет на урожайность зерна. Завод J 54: 888–898. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Dai X, You C, Chen G, Li X, Zhang Q и др. (2011) OsBC1L4 кодирует COBRA-подобный белок, который влияет на синтез целлюлозы в рисе. Растение Мол Биол 75: 333–345. [PubMed] [Академия Google]
38.
Dai X, You C, Wang L, Chen G, Zhang Q и др. (2009)Молекулярная характеристика, характер экспрессии и функциональный анализ семейства OsBC1L в рисе. Растение Мол Биол
71: 469–481. [PubMed] [Google Scholar]
Растение Мол Биол
71: 469–481. [PubMed] [Google Scholar]
39. Xiong G, Li R, Qian Q, Song X, Liu X и др. (2010) Рисовый динамин-родственный белок DRP2B опосредует перенос мембран и, таким образом, играет критическую роль во вторичном биосинтезе целлюлозы клеточной стенки. Завод J 64: 56–70. [PubMed] [Google Scholar]
40. Уденфренд С., Кодукула К. (1995) Как образуются гликозил-фосфатидилинозитол-заякоренные мембранные белки. Анну Рев Биохим 64: 563–591. [PubMed] [Google Scholar]
41. Ning J, Zhang B, Wang N, Zhou Y, Xiong L (2011) Увеличенный угол листа1, Raf-подобный MAPKKK, который взаимодействует с семейством ядерных белков, регулирует образование механической ткани в суставе пластинки риса. Растительная клетка 23: 4334–4347. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42.
Маклин Б.В., Брей М.Р., Борастон А.Б., Гилкс Н.Р., Хейнс К.А. и соавт. (2000) Анализ связывания углеводсвязывающего модуля семейства 2а из Cellulomonas fimi Ксиланаза 10А к целлюлозе: специфичность и идентификация функционально важных аминокислотных остатков. Белок англ
13: 801–809. [PubMed] [Google Scholar]
Белок англ
13: 801–809. [PubMed] [Google Scholar]
43. Чжан Б., Дэн Л., Цянь К., Сюн Г., Цзэн Д. и др. (2009) Миссенс-мутация в трансмембранном домене CESA4 влияет на содержание белка в плазматической мембране и приводит к аномальному биосинтезу клеточной стенки у риса. Растение Мол Биол 71: 509–524. [PubMed] [Google Scholar]
44. Танака К., Мурата К., Ямадзаки М., Оносато К., Мияо А. и др. (2003)Три различных гена каталитической субъединицы синтазы целлюлозы риса необходимы для синтеза целлюлозы во вторичной стенке. Завод Физиол 133: 73–83. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Тигесен А., Оддершеде Дж., Лилхольт Х., Томсен А., Сталь К. (2005) Об определении кристалличности и содержания целлюлозы в растительных волокнах. Целлюлоза 12: 563–576. [Google Scholar]
47.
Сигал Л., Крили Дж., Мартин А., Конрад С. (1959) Эмпирический метод оценки степени кристалличности нативной целлюлозы с использованием рентгеновского дифрактометра. Разрешение текста J
29: 786–794. [Google Академия]
[Google Академия]
48. Паттерсон А. (1939) Формула Шеррера для определения размера частиц с помощью рентгеновских лучей. Физическая версия 56: 978–982. [Google Академия]
49. Hoch HC, Galvani CD, Szarowski DH, Turner JN (2005) Два новых флуоресцентных красителя, применимых для визуализации клеточных стенок грибов. Микология 97: 580–588. [PubMed] [Google Scholar]
50. Борастон А.Б., Болам Д.Н., Гилберт Х.Дж., Дэвис Г.Дж. (2004)Модули связывания углеводов: точная настройка распознавания полисахаридов. Биохим Дж 382: 769–781. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Georgelis N, Yennawar NH, Cosgrove DJ (2012)Структурная основа для энтропийно-управляемого связывания целлюлозы модулем связывания целлюлозы типа A (CBM) и бактериальным экспансином. Proc Natl Acad Sci USA 109: 14830–14835. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52.
Сато К., Ито С., Фуджи Т., Судзуки Р., Такенучи С. и др. (2010) Последовательность, подобная углеводному связывающему модулю (CBM), имеет решающее значение для функции CWA1/BC1 риса в правильной сборке материалов вторичной клеточной стенки. Завод Sig & Behav
5: 1433–1436. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Завод Sig & Behav
5: 1433–1436. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Лейхтер Р., Вольф К., Циммерманн М. (1998) Выделение кДНК арабидопсиса, дополняющей Schizosaccharomyces pombe мутант с дефицитом синтеза фитохелатинов. Завод Физиол 117: 1526. [Google Scholar]
54. Najmudin S, Guerreiro CIPD, Carvalho AL, Prates JAM, Correia MAS, et al. (2006) Ксилоглюкан распознается модулями связывания углеводов, которые взаимодействуют с цепями β-глюкана. J Биол Хим 281: 8815–8828. [PubMed] [Google Scholar]
55. Бензиман М., Хейглер К.Х., Браун Р.М., Уайт А.Р., Купер К.М. (1980)Биогенез целлюлозы: полимеризация и кристаллизация — связанные процессы в Ацетобактерии хилинум . Proc Natl Acad Sci USA 77: 6678–6682. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Somerville C, Bauer S, Brininstool G, Facette M, Hamann T, et al. (2004) На пути к системному подходу к пониманию клеточных стенок растений. Наука 306: 2206–2211. [PubMed] [Google Scholar]
57. Эндлер А., Перссон С. (2011)Синтазы и синтез целлюлозы у арабидопсиса. Мол Завод
4: 199–211. [PubMed] [Google Scholar]
Эндлер А., Перссон С. (2011)Синтазы и синтез целлюлозы у арабидопсиса. Мол Завод
4: 199–211. [PubMed] [Google Scholar]
58. Харрис Д., ДеБолт С. (2008)Относительная кристалличность растительной биомассы: исследования сборки, адаптации и акклиматизации. PLoS один 3: е2897. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Zhou Y, Li S, Qian Q, Zeng D, Zhang M, et al. (2008) BC10, DUF266-содержащий и локализованный в Golgi мембранный белок типа II, необходим для биосинтеза клеточной стенки риса ( Oryza sativa L.). Завод J 57: 446–462. [PubMed] [Google Scholar]
60. Petersen TN, Brunak S, von Heijne G, Nielsen H (2011)SignalP 4.0: различение сигнальных пептидов из трансмембранных областей. Нат Методы 8: 785–786. [PubMed] [Академия Google]
61. Eisenhaber B, Bork P, Eisenhaber F (1999)Предсказание потенциальных сайтов модификации GPI в последовательностях пропротеинов. Джей Мол Биол 292: 741–758. [PubMed] [Google Scholar]
62.
Летуник И. , Доеркс Т., Борк П. (2012) SMART 7: последние обновления ресурса аннотаций белковых доменов. Нуклеиновые Кислоты Res
40: 302–305. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Доеркс Т., Борк П. (2012) SMART 7: последние обновления ресурса аннотаций белковых доменов. Нуклеиновые Кислоты Res
40: 302–305. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Пунта М., Коггилл П.С., Эберхардт Р.Ю., Мистри Дж., Тейт Дж. и др. (2012) База данных семейств белков Pfam. Нуклеиновые Кислоты Res 40:290–301. [Бесплатная статья PMC] [PubMed] [Google Scholar]
64. Корпет Ф. (1988) Множественное выравнивание последовательностей с иерархической кластеризацией. Нуклеиновые Кислоты Res 16: 10881–10890. [Бесплатная статья PMC] [PubMed] [Google Scholar]
65. Mutwil M, Obro J, Willats WG, Persson S (2008) GeneCAT — новые веб-инструменты, сочетающие анализ BLAST и коэкспрессии. Нуклеиновые Кислоты Res 36: 320–326. [Бесплатная статья PMC] [PubMed] [Google Scholar]
66.
Feiz L, Irshad M, Pont-Lezica RF, Canut H, Jamet E (2006)Оценка препаратов клеточных стенок для протеомики: новая процедура очистки клеточных стенок от гипокотилей арабидопсиса. Растительные методы
2: 10.