определение, признаки, границы и критерии выделения
Главная » Что мы делаем » Полнотекстовые статьи и лекции » Москалюк Т.А. » Курс лекций по биогеоценологии » Лекция 3. Фитоценоз как гла…
Москалюк Т.А.
Список литературы
Воронов А.Г. Геоботаника. Учеб. Пособие для ун-тов и пед. ин-тов. Изд. 2-е. М.: Высш. шк., 1973. 384 с.
Ипатов В.С., Кирикова Л.А. Фитоценология: Учебник. СПБ: Изд-во ЛГУ, 1997. 316 с.
Степановских А.С. Общая экология: Учебник для вузов. М.: ЮНИТИ, 2001. 510 с.
Сукачев В.Н. Основы лесной типологии и биогеоценологии. Избр. тр. Л.: Наука, 1972. Т. 3. 543 с.
Программа и методика биогеоценологических исследований / Изучение лесных биогеоценозов / М.: Наука, 1974. С. 281-317.
Цветков В.Ф. Лесной биогеоценоз. Архангельск, 2003. 2-е изд. 267 с.
Вопросы
1. Выделение границ биогеоценоза.
2. Биогеоценозообразующая роль фитоценоза в ландшафте.
3. Определение понятия «фитоценоз».
4. Основные признаки фитоценоза.
5. Минимальный размер площади выявления фитоценоза.
6. О границах фитоценоза. Понятие о континууме растительного покрова.
7. Различия понятий «фитоценоз», «ассоциация» и «растительное сообщество».
Чтобы управлять биогеоценотическими процессами, надо знать закономерности, которым они подчинены. Эти закономерности изучает ряд наук: метеорология, климатология, геология, почвоведение, гидрология, различные отделы ботаники и зоологии, микробиология и др. Биогеоценология же синтезирует результаты перечисленных наук под определенным углом зрения, обращая основное внимание на взаимодействия компонентов биогеоценозов между собой и вскрывая общие закономерности, управляющие этими взаимодействиями. Эта область знания изучает биогеоценоз в целом, исследует свойственные ему процессы.
1. Выделение границ биогеоценоза
Известно, что только при установлении точных границ биогеоценоза возможно проведение ее анализа. И чем конкретнее и точнее ограничивается биогеоценоз в пространстве, тем с большей объективностью возможна количественная характеристика протекающих в нем процессов и явлений.
Общеизвестна трудность точного установления этих границ, «… проведение границ между ними нередко носит условный, в известной степени субъективный характер…». В.Н. Сукачев писал по этому поводу: «…Различные биогеоценозы имеют, конечно, различную вертикальную мощность, например лесные, степные, пустынные и т. п. Однако, как правило, можно считать, что верхняя граница биогеоценоза располагается на несколько метров выше растительного покрова, нижняя часть лежит на несколько метров ниже поверхности почвы…» (Основы лесной биогеоценологии, 1964: 32.
Примерно так же понимает границы биогеоценозов Е. М. Лавренко (1962) при выделении части биосферы, названной им » фитосферой «.
Возникает вопрос, какие же критерии наиболее доступны при выделении биогеоценозов в природе?
1. Начинать надо с анализа рельефа. Хотя рельеф, как известно, и не входит в состав компонентов биогеоценоза, но он является очень важным фактором его существования. Для первой ориентировки в лесных биогеоценозах при их выделении и разграничения в природе, может играть очень большую роль. С него начинают описание биогеоценоза.
С него начинают описание биогеоценоза.
2. В пределах однородного рельефа наиболее показательным признаком однородности биогеоценоза является однородность почвы и растительного покрова. Из этих двух показателей для выделения биогеоценозов особенно пригодна однородность растительного покрова, благодаря ее наглядности. Поэтому при разграничении биогеоценозов в природе целесообразно пользоваться именно фитоценозом. Границы каждого биогеоценоза в отдельности определяются границами фитоценоза.
Наглядность – это очень важная деталь, но главное заключается в том, что среди компонентов биогеоценоза именно фитоценозу, принадлежит определяющая – биогеоценозообразующая, роль.
2. Биогеоценозообразующая роль фитоценоза в ландшафтеФитоценоз – главная составляющая, узловая во всех отношениях подсистема биогеоценоза, в которой протекают основные процессы образования и преобразования того, что является основой жизни на планете – органического вещества. Он определяет пространственные границы биогеоценоза, его структуру и облик, внутренний климат, состав, обилие и распределение животных, микроорганизмов, особенности и интенсивность материально-энергетического обмена всей системы биогеоценоза.
Он определяет пространственные границы биогеоценоза, его структуру и облик, внутренний климат, состав, обилие и распределение животных, микроорганизмов, особенности и интенсивность материально-энергетического обмена всей системы биогеоценоза.
Фитоценозы служат:
1) главными приемниками и трансформаторами солнечной энергии,
2) главными поставщиками продукции в биогеоценозе,
3) в их структуре объективно отражаются все процессы, протекающие в биогеоценозе,
4) при этом они легко доступны для изучения непосредственно в природе,
5) для них на протяжении нескольких десятков лет разработаны и разрабатываются эффективные полевые методы исследований и методы камеральной обработки фактических материалов.
Исходя из вышесказанного, подробное изучение фитоценоза является обязательной частью любого биогеоценологического исследования. С растительности всегда начинается изучение любой природной системы. В наших лекциях основное внимание будет уделено фитоценозу и методам его изучения. Тем более, что многие закономерности, свойственные фитоценозу, распространяются и на зооценоз и микроорганизмы.
Тем более, что многие закономерности, свойственные фитоценозу, распространяются и на зооценоз и микроорганизмы.
В общей форме изучение фитоценозов сводится к решению следующих важнейших вопросов:
- Определение роли фитоценоза в накоплении органических веществ и энергии и превращениях вещества и энергии в общей системе биогеоценоза (узловой вопрос!).
- Определение роли фитоценоза в динамике биогеоценоза.
- Определение характера и степени воздействия фитоценоза на остальные компоненты биогеоценоза.
- Соответственно – определение влияния других компонентов биогеоценоза на свойства, особенности и эффективность «работы» фитоценоза.
- Определение характера и степени воздействия фитоценоза на соседние биогеоценозы.
- Определение формы, способов и средств прямого и косвенного воздействия на фитоценоз со стороны хозяйственной деятельности человека с целью повышения биологической продуктивности биогеоценоза и усиления других его полезных свойств.
3.
 Определение понятия «фитоценоз»
Определение понятия «фитоценоз»Первое определение растительного сообщества было дано Г.Ф. Морозовым (1904) для леса, а затем распространено В.Н. Сукачевым (1908) на все растительные сообщества. Термин «фитоценоз» был применен в 1915 г. И.К. Пачоским для «чистых зарослей» (образованных одним видом растений), а для всех сообществ – В.Н. Сукачевым (1917) и Г. Гамсом (Gams, 1918).
Фитоценоз, как и биогеоценоз, следует понимать как географическое явление, а именно – каждый наземный фитоценоз занимает определенную территорию со свойственными ей гидрологическим режимом, микрорельефом, микроклиматом, почвами.
В. Н. Сукачев (1956) весьма удачно дает определение фитоценоза: «… Фитоценоз, или растительное сообщество, – совокупность растений, произрастающих совместно на однородной территории, характеризующаяся определенным составом, строением, сложением и взаимоотношениями растений как друг с другом, так и с условиями среды. Характер этих взаимоотношений определяется, с одной стороны, жизненными, иначе, экологическими свойствами растений, с другой стороны, свойствами местообитания, т.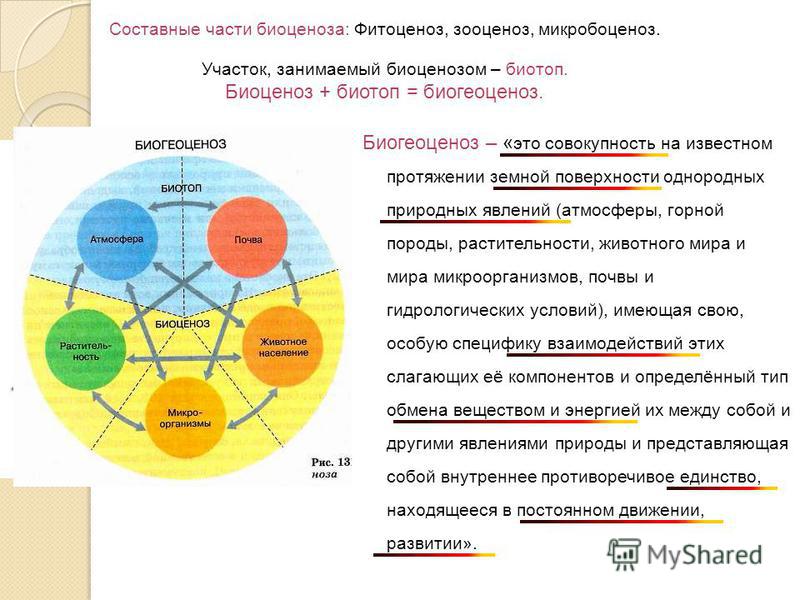 е. характером климата, почвы и влиянием человека и животных …».
е. характером климата, почвы и влиянием человека и животных …».
Строение и состояние фитоценоза хорошо отражает и конкуренцию и взаимопомощь растений.
ПРИМЕР . Широколиственно-липовый с лиановой растительностью лещиновый разнотравный фитоценоз в средней части склона сев.-сев.-вост. экспозиции (высотные отметки 250-300 м н.ур.м.; крутизна склона в среднем составляет 15-20).
Древостой состоит из 2-х ярусов. Для него характерны очень высокая сомкнутость крон – 0,97, Деревья смыкаются кронами и под их пологом царит полумрак. Первый ярус образован дубом монгольским, липами амурской и маньчжурской, орехом маньчжурским, единичными старыми деревьями берёзы чёрной и клёна мелколистного.
Большая часть лип представлена стройными полнодревесными деревьями с гладкими стволами. Средние таксационные показатели деревьев 1-ого яруса: Дср 18-20 см, Нср – 17-18 м. В хорошо выраженном втором ярусе преобладают липа – в основном более молодые, чем в верхнем ярусе, и отставшие в росте деревья, клён мелколистный.
Незначительна примесь ильма горного, маакии амурской, граба сердцелистного; единичны диморфант, клён ложнозибольдов и мелкоплодник. Кроме того, в него «проникают» трескун, бересклет Максимовича, основная масса особей которых сосредоточена в подлеске.
Дальнейшее существование любого леса обеспечивается возобновлением видов материнского древостоя. Подрост в количестве 8,6 тыс. экз.\га представлен всеми видами древостоя. В его видовом составе доминирует клен мелколистный, в самосеве единичны особи темнохвойной породы – пихты цельнолистной (Abies holophylla).. Распределение подроста по площади – равномерно-групповое (клен, липа, трескун) и одиночными особями (ильм, орех, маакия и др.).
Подлесок густой, в нем доминирует лещина маньчжурская, обычны чубушник, элеутерококк, реже встречается большими кустами смородина Максимовича, жимолость раннецветущая, единичны бересклеты большекрылый и малоцветковый, клён зеленокорый. Редко группами растёт калина буреинская и, как правило, вместе с нею – одиночные побеги барбариса амурского.
Из-за высокой сомкнутости древесно-кустарниковых ярусов травостой разрежен. Кроме лесного мака весеннего в нем доминируют василисник клубненосный, осоки: уссурийская, возвратившаяся, длинноносая, звездчатка Бунге, подмаренник даурский, папоротники. Как и в древостое, в травостое можно выделить может яруса. Верхний, высотой до 1 м, образуют растущие повсеместно виды крупнотравья: пион горный, клопогон даурский, дудник даурский и амурский, воронец заострённый, подлесник красноцветковый, лилия двурядная; единичны волжанка азиатская и стеблелист мощный. Иногда мелкие густопокровные микрогруппировки образуют травы средних размеров (разнотравье), высотой до 0,5 м – смилацина волосистая, хвощ зимующий, глухая крапива, ландыш, и мелкотравье, высотой не более 0,25 м: шлемник уссурийский, лютик Франше, майник двулистный, тригонотис укореняющийся, адокса мускусная, мерингия бокоцветная, фиалки Колина, копытень Зибольда, джефферсония сомнительная, разные виды хохлаток.
Кроме растений, образующих ярусы, в описываемом фитоценозе можно выделить и так называемые внеярусные растения, например лианы актинидии, лимонника, винограда.
Все виды трав можно разделить на группы по сезонному развитию (одни – весенние эфемероиды (ветреницы, хохлатки, адонис, ллойдия, и др.), проходят цикл развития в течение месяца и уже в июне находятся в состоянии покоя. У других (лилия двурядная, лихнис сверкающий, стеблелист мощный и др.) кульминация развития приходится на июль, третьи (плектрантус, десмодиум, соссюреи, акониты) цветут и остаются зелеными и в сентябре), по происхождению (таежные леса, мелколиственные, неморальные, луговые и пр.), по обилию (одни из них встречаются в столь значительном количестве, что образуют сплошной покров, другие редки, третьи единичны).
Таким образом, в этом лесу можно выделить шесть надземных ярусов: два древесных, один кустарниковый (с подростом) и три травяных.
Выкопав в таком лесу траншею, можно наблюдать и подземную ярусность (правда, менее резко выраженную, чем надземная): корни и корневища трав располагаются в менее глубоких горизонтах почвы, корни кустарников и деревьев – в более глубоких.
 Незначительна примесь ильма горного, маакии амурской, граба сердцелистного; единичны диморфант, клён ложнозибольдов и мелкоплодник. Кроме того, в него «проникают» трескун, бересклет Максимовича, основная масса особей которых сосредоточена в подлеске.
Незначительна примесь ильма горного, маакии амурской, граба сердцелистного; единичны диморфант, клён ложнозибольдов и мелкоплодник. Кроме того, в него «проникают» трескун, бересклет Максимовича, основная масса особей которых сосредоточена в подлеске.

 Благодаря подземной ярусности растения используют разные слои почвы для получения влаги и питательных веществ.
Благодаря подземной ярусности растения используют разные слои почвы для получения влаги и питательных веществ.Таким образом, фитоценоз характеризуется:
1. определенным видовым составом образующих его растений,
2. определенным их обилием,
3. определенной структурой и
4. приуроченностью к определенному местообитанию.
4. Основные признаки фитоценоза
Всякое ли сочетание растений нужно считать фитоценозом, и если не всякое, то какие критерии существуют для того, чтобы отличать фитоценоз от других сочетаний растений?
Существенные признаки фитоценоза – фитоценотические отношения (отношения между растениями) и наличие фитоценотической среды.
Создание фитосреды – первый по времени появления признак фитоценоза, т.к. воздействие растительных организмов на среду уже может быть там, где еще нет влияния растений друг на друга. ФИТОЦЕНОТИЧЕСКАЯ СРЕДА начинает формироваться еще в то время, когда отдельные растения, появившиеся на ранее лишенной или не имевшей связного растительного покрова территории, растут разобщено, не образуя сплошного покрова.
Уже на первых стадиях развития растительности изменяются условия микроклимата, с мертвыми растениями в почву или в грунт вносятся одни химические вещества, а живыми растениями извлекаются другие, изменяется характер микрорельефа (например, образуются шлейфы из пылеватых и песчаных частиц у стеблей растений), словом, происходит преобразование среды растениями. В дальнейшем, через взаимодействия растений, фитоценоз все более изменяет среду и создает свою фитосреду. При этом условия среды в разных частях фитоценоза (на поверхности почвы, на стволах и в кронах деревьев, на разной высоте над поверхностью почвы и т. д.) неодинаковы.
Наличие фитоценотических отношений – наиболее существенная особенность фитоценоза, но ВЗАИМОДЕЙСТВИЕ МЕЖДУ РАСТЕНИЯМИ начинается несколько позже, чем воздействие растений на их местообитание. Оно может иметь место только при определенной густоте растительного покрова. Однако подметить этот момент, когда начинается взаимодействие между растениями, очень трудно, поскольку оно не всегда предполагает прямой контакт между организмами.
Поэтому к фитоценозам следует относить различные стадии развития растительного покрова, кроме самых первых моментов поселения растений на территории, лишенной растительности.
Другой вопрос – степень выраженности фитоценотической среды и степень выраженности отношений между растениями в фитоценозах. В некоторых пустынях, в полярных тундрах растительный покров настолько разрежен, что его нельзя считать образованным из фитоценозов. При такой разреженности покрова очень трудно определить степень воздействия друг на друга корневых систем растений и почти невозможно при существующих методах исследования выяснить степень влияния на среду и на другие растительные организмы микроскопических растений – водорослей и бактерий, которая может быть весьма значительной. В таких условиях среды, вероятно, каждый участок длительно существующего и, следовательно, приспособившегося к среде растительного покрова следует расчленять на фитоценозы.
Поскольку фитоценоз есть не всякое собрание растительных видов, а лишь сочетание вполне закономерное, сложившееся в результате длительного исторического процесса и находящееся в связи с внешними условиями существования, то В. В. Алехин и другие представители московской фитоценологической школы полагают, что в качестве одного из обязательных признаков сообщества выдвигается «способность к восстановлению» или «способность к относительному восстановлению» (Прозоровский. 1956).
В. Алехин и другие представители московской фитоценологической школы полагают, что в качестве одного из обязательных признаков сообщества выдвигается «способность к восстановлению» или «способность к относительному восстановлению» (Прозоровский. 1956).
С этой точки зрения нельзя считать фитоценозами культурную растительность, группировки растений, поселяющиеся на лишенных растительности участках, а также все те естественные сочетания растений, которые не восстанавливаются после их нарушения или уничтожения.
Так, нельзя было бы считать фитоценозами не только вторичные березовые или осиновые леса, появляющиеся на месте первичных после их вырубания, но и первичные северные леса умеренного пояса, обитающие на участках с неглубоко залегающими грунтовыми водами, так как эти леса после вырубки или выгорания не возобновляются, а площади лесосек и гарей заболачиваются. Нельзя считать и кустарниковые сообщества (леспедечники, облепишники), сменившие высокоствольные фитоценозы (дубняки, кедрово-широколиственные).
Вряд ли такая точка зрения может быть признана правильной. Ведь и в культурных сообществах, и в первичных и вторичных лесах, и в пионерных группировках растений (может быть, за исключением самых начальных стадий их существования) есть те признаки, которые составляют существенные особенности фитоценоза: создание фитосреды и наличие фитоценотических отношений.
Между растениями в сложившемся фитоценозе существуют два типа взаимоотношений:
1) конкуренция друг с другом из-за средств жизни или борьба за существование в том широком смысле, как ее понимал Ч. Дарвин. Это с одной стороны, ослабляет растения, но с другой – составляет основу естественного отбора – важнейшего фактора видообразования и, следовательно, процесса эволюции.
ПРИМЕР. Дифференциация и самоизреживания древостоя с возрастом или ухудшением условий произрастания – выживают сильнейшие. От десятков тысяч проростков и самосев к возрасту спелости остается менее 1%.

В процессе естественного отбора в состав фитоценоза вошли такие виды, которые взаимосвязаны друг с другом или зависят друг от друга. В него входят не только цветковые, голосеменные, папоротникообразные растения, плауны, хвощи, мхи, но и низшие растительные организмы: грибы, водоросли, бактерии, лишайники.
Сказанное о роли естественного отбора в формировании сообществ верно лишь для ненарушенных, сложившихся естественных фитоценозов. В фитоценозах, развивающихся на ранее лишенных растительного покрова участках, на первых стадиях развития отсутствует прямое влияние отдельных растений друг на друга и поэтому взаимосвязи между отдельными видами еще не выражены.
2) взаимопомощь – растения в фитоценозе оказывают друг на друга благоприятное воздействие: под пологом деревьев обитают тенелюбивые травы, которые не могут расти или плохо растут на открытых местах; стволы деревьев и ветви кустарников служат механической опорой лианам, на которых в свою очередь поселяются не связанные с почвой эпифиты.
5. Минимальный размер площади выявления фитоценоза
Как же определить минимальный размер площади, на которой можно выделить фитоценоз?
Очевидно, наименьшая территория выявления фитоценоза должна иметь такие размеры, чтобы могли проявиться все признаки самого фитоценоза (видовой состав, структура и проч.), а также все основные особенности почвы, микроклимата, микрорельефа поверхности, словом, особенности фитосреды.
Само собой разумеется, что для разных фитоценозов размеры этой наименьшей территории неодинаковы: чем проще строение фитоценоза, чем меньше его влияние на среду обитания, тем меньше размеры территории.
В умеренном поясе они меньше для лугов по сравнению с лесами. Для лесов умеренного пояса они меньше по сравнению с тропическими лесами.
6. О границах фитоценоза. Понятие о континууме растительного покрова
Между фитоценозами могут быть резкие границы, но чаще же переходы постепенны, незаметны. Этим вызваны трудности выделения фитоценозов. Постепенность переходов от одного типа фитоценоза к другому – следствие постепенного изменения экологических условий местообитания. Если изменяющиеся значения какого-либо фактора (например, условий увлажнения, засоления и т. д.) нанести на график, то им будут соответствовать и постепенно сменяющие друг друга сочетания видов растений. Исходя из этого, Л.Г. Раменским было разработано учение о непрерывности растительного покрова (московская школа геоботаники), или, как его часто называют, учение о континууме . Как указывает Л.Г. Раменский (1910, 1925, 1937, 1938), «…способность растений образовывать разнообразные сочетания прямо-таки неисчерпаема…», т.е. число ассоциаций неограниченно.
Постепенность переходов от одного типа фитоценоза к другому – следствие постепенного изменения экологических условий местообитания. Если изменяющиеся значения какого-либо фактора (например, условий увлажнения, засоления и т. д.) нанести на график, то им будут соответствовать и постепенно сменяющие друг друга сочетания видов растений. Исходя из этого, Л.Г. Раменским было разработано учение о непрерывности растительного покрова (московская школа геоботаники), или, как его часто называют, учение о континууме . Как указывает Л.Г. Раменский (1910, 1925, 1937, 1938), «…способность растений образовывать разнообразные сочетания прямо-таки неисчерпаема…», т.е. число ассоциаций неограниченно.
Многие ученые, следуя этому учению, не признают реальность существования фитоценозов. Вопрос о выделении (на графике и на местности) границ того или иного фитоценоза оказывается, по Л.Г. Раменскому, до некоторой степени условным и зависит не только от родства этого фитоценоза с соседними, но и от «целевой установки, работы». Иными словами, границы между двумя фитоценозами могут отсутствовать и в разных случаях или разными исследователями могут быть проведены неодинаково.
Иными словами, границы между двумя фитоценозами могут отсутствовать и в разных случаях или разными исследователями могут быть проведены неодинаково.
В настоящее время учение о непрерывности растительного покрова получило широкое распространение, особенно в работах американских ученых: Х. Глизона (Glеаsоп, 1939), Д. Куртиса (Curtis, 1955, 1958), Р. Уиттекера (Whittaker, 1953, 1956, 1960), Ф. Иглера (Egler, 1951, 1954), польского исследователя В. Матушкевича (Matuszkievicz, 1948). Основная идея этого учения – невозможность уложить все разнообразие сочетаний растений с множеством различных переходных группировок в ограниченное число ассоциаций. Сторонники этой концепции, как правило, считают фитоценозы (ассоциации, растительные сообщества) условными, абстрактными категориями, не существующими в природе, хотя, как полагают некоторые из них, необходимыми с практической или – для теоретической точки зрения.
В связи с этим вопрос о существовании резких или постепенных границ между фитоценозами приобретает существенное значение. Л.Г. Раменский, Т.А. Работнов (1967) и другие сторонники теории континуума полагают, что правилом являются постепенные, неясные границы, а исключением – резкие.
Л.Г. Раменский, Т.А. Работнов (1967) и другие сторонники теории континуума полагают, что правилом являются постепенные, неясные границы, а исключением – резкие.
Сторонники другого учения (ленинградская школа) – о дискретности растительного покрова, например В.Н. Сукачев, Г. Дюpьe, в противоположность взгляду Л.Г. Раменского, считают, что, как правило, границы между ассоциациями резкие, но иногда возможны и плавные переходы.
На самом деле право на существование имеют обе теории. Характер границ между фитоценозами отражают влияние на среду растений-эдификаторов. Плавные переходы от одного ценоза к другому чаще наблюдаются там, где влияние эдификаторов не столь уж сильно изменяет среду, например на лугах, а резкие – там, где один мощный эдификатор сменяется другим (например, на границах участков леса, образованных различными древесными породами).
Как указывал Б.А. Быков (1957), эдификаторы своим влиянием в значительной степени определяют резкость границ даже при постепенной смене условий местообитания. Поэтому «…границы растительных ассоциаций бесспорно намечены в природе резче, нежели границы местообитаний…» (Ниценко, 1948).
Поэтому «…границы растительных ассоциаций бесспорно намечены в природе резче, нежели границы местообитаний…» (Ниценко, 1948).
7. Различия понятий «фитоценоз», «ассоциация» и «растительное сообщество»
Между терминами «фитоценоз» и «растительное сообщество» многие ученые долгое время ставили знак равенства, употребляя их как синонимы. В.Н. Сукачев неоднократно указывал, что термин «фитоценоз» (растительное сообщество) можно применять и к конкретным участкам растительного покрова, и для обозначения таксономических единиц различных рангов: ассоциации, формации, типа растительности и т. д.
В последнее время (Воронов, 1973) термин «фитоценоз» применяется только к конкретным участкам растительного покрова, соответствующим приведенному выше определению В.Н Сукачева..
Ассоциацией называют типологическую единицу фитоценозов. Тот же смысл вкладывает в этот термин и С.М. Разумовский (1981). В его трактовке ассоциация – элементарная единица растительного покрова, объединяет участки с одними и теми же видами каждого яруса и одной и той же сукцессионной тенденцией. Проводя аналогию, можно сказать, что фитоценоз и ассоциация так относятся друг к другу, как конкретное растение и вид этого растения.
Проводя аналогию, можно сказать, что фитоценоз и ассоциация так относятся друг к другу, как конкретное растение и вид этого растения.
ПРИМЕР. Проезжая по трассе Владивосток-Уссурийск, можно сказать, что почти вся лесная растительность на этом отрезке представлена вторичными дубовыми лесами. Участки этих лесов очень похожи один на другой. Древостой – дубовый с единичной примесью березы даурской. Подлесок фрагментарный, представлен лещиной и леспедецей. В напочвенном покрове – разнотравье и осоки. Тем не менее, несмотря на очень высокое сходство, нельзя найти даже двух участков с совершенно одинаковыми фитоценозами.
Иными словами, все эти похожие фитоценозы – однотипны, и в совокупности представляет одну ассоциацию, или один тип леса – дубняк осоково-разнотравный.
Ассоциация – первая ступень в системе таксономических единиц растительного покрова различного ранга. Последующие ступени составляют: группа ассоциаций, формация, группа формаций, класс формаций, тип растительности; имеются и промежуточные категории.
Такое обилие иерархических ступеней и вызвало потребность в особом термине, который можно было бы применять к таксономической категории любого ранга, Г. Дюрье (Du-Rietz, 1936) в качестве такого термина предложил «растительное сообщество». Это понятие более широкое, чем «фитоценоз». Оно не имеет определенного объема. Так, В.В. Алехин (1950) пишет, что «… растительное сообщество … это растительные объединения как более крупного, так и более мелкого объема, как лес вообще, так и лес хвойный, и подразделения последнего, а также данный конкретный его участок…».
Таким образом, растительным сообществом обозначается любой растительный «коллектив», где растения находятся в определенных взаимоотношениях, без указания на его размерность и характер организации. В этом случае растительное сообщество может рассматриваться как явление совместной общественной жизни растений. В таком широком смысле и использован этот термин В.В. Алехиным (1935, 1950, 1951)
Фитоценозы, относящиеся к одной и тои же ассоциации, располагаются разбросанно. Обычно они граничат с фитоценозами, принадлежащими к другим ассоциациям. На местности обычно встречаются сочетания фитоценозов, нередко они довольно заметно отличаются друг от друга. Это связано с различиями других природных компонентов, а в итоге образуются разные биогеоценозы, или так называемые природно-территориальные комплексы. Именно с такими комплексами обычно и приходится иметь дело при использовании растительных ресурсов.
Обычно они граничат с фитоценозами, принадлежащими к другим ассоциациям. На местности обычно встречаются сочетания фитоценозов, нередко они довольно заметно отличаются друг от друга. Это связано с различиями других природных компонентов, а в итоге образуются разные биогеоценозы, или так называемые природно-территориальные комплексы. Именно с такими комплексами обычно и приходится иметь дело при использовании растительных ресурсов.
Фитоценоз (растительное сообщество) | Ботаника. Реферат, доклад, сообщение, кратко, презентация, лекция, шпаргалка, конспект, ГДЗ, тест
Растительное сообщество — это исторически сложившаяся под влиянием условий среды совокупность различных видов растений, произрастающих совместно на относительно однородной территории, характеризующаяся определённым составом, строением и взаимоотношениями растений как друг с другом, так и с условиями среды.
В природе растения очень редко встречаются изолированно, отдельными особями. Это имеет место лишь при заселении какой-либо свободной от растений территории, например песчаных отмелей рек, карьеров, выработанных торфяников и других мест. Чаще всего растения образуют сообщества, которые называют фитоценозами.
Чаще всего растения образуют сообщества, которые называют фитоценозами.
Совокупность растительных сообществ на какой-то конкретной территории образует её растительный покров (растительность). Растительные сообщества тесно связаны со всеми организмами, обитающими в нём. Растения, животные, грибы, бактерии, совместно населяющие определённую территорию, образуют
Видовой состав
Видовой состав является важнейшим признаком фитоценоза, он характеризует разнообразие в нём видов и соотношение их численности или массы. Чем выше видовое разнообразие, тем большей устойчивостью обладает фитоценоз.
Чем выше видовое разнообразие, тем большей устойчивостью обладает фитоценоз.
Видовой состав растительного сообщества зависит от условий среды, длительности его существования, видового богатства флоры
, в состав которой входит фитоценоз, и история его формирования.Различают бедные и богатые видами сообщества.
- Там, где условия абиотической среды наиболее благоприятны для жизни растений, формируются богатые видами фитоценозы. Примерами их являются влажные тропические леса, хвойно-широколиственные леса и пойменные луга в лесной зоне.
- Где условия среды не благоприятны для большинства видов растений, растительные сообщества имеют бедный видовой состав, как, например, в северной тундре, арктических и глинистых пустынях.
Молодые, формирующиеся фитоценозы имеют обычно меньшее число видов, чем давно сложившиеся.
Доминантные виды
Количественное участие видов растений в сложении фитоценоза неодинаковое. Это зависит от размеров их особей, а также биологических и экологических особенностей. Виды, преобладающие по численности, называют доминантными. Они занимают господствующее положение в сообществе и по ним называют фитоценозы, например:
Это зависит от размеров их особей, а также биологических и экологических особенностей. Виды, преобладающие по численности, называют доминантными. Они занимают господствующее положение в сообществе и по ним называют фитоценозы, например:
Эдификаторы
Не все доминирующие виды оказывают одинаковое влияние на микроклимат сообщества. Среди них есть виды, которые своей жизнедеятельностью в наибольшей степени создают среду для всего сообщества. Такие виды называют эдификаторами. Например, в еловом лесу эдификатором является ель, в сосновом — сосна, а в степях — дерновинные злаки.
Загрузка…
Редкие виды
Кроме относительно небольшого числа видов-доминантов в состав фитоценозов входит множество малочисленных и редких видов. Они также очень важны для жизни сообщества, так как создают его видовое богатство, увеличивают разнообразие биоценотических связей и служат резервом для замещения доминантов при изменении условий среды.
Пространственная структура фитоценоза
Пространственная структура фитоценоза отражает закономерности распределения видов растений в пределах занимаемого ими участка, который называют биотопом. В наземных фитоценозах она определяется распределением надземной и подземной массы растений.
Ярусность
При совместном произрастании разных по высоте растений фитоценоз имеет ярусное сложение: ассимилирующие надземные органы растений и подземные их части располагаются в несколько слоёв, по-разному используя и изменяя среду. Особенно чётко выражена ярусность в лесных сообществах. В хвойных лесах обычно выделяются три яруса: древесный, подлесок и травяно—кустарничковый, но в некоторых лесах могут быть только два яруса, как, например, в сосняках-беломошниках — древесный и лишайниковый. Подземные части растений тоже располагаются ярусно. Корни деревьев, как правило, проникают на большую глубину, чем у кустарников, а корневые системы у большинства травянистых растений располагаются ближе к поверхности почвы.
Мозаичное (групповое) распределение
В горизонтальном направлении фитоценозам свойственны мозаичное или групповое распределение растений, что обусловлено неоднородностью почв, микрорельефа и биологическими особенностями видов, например, их вегетативным размножением, опадением семян вблизи материнского растения и др.
Взаимоотношения между растениями
см. Взаимоотношения между растениями
Взаимоотношения между растениями в фитоценозе играют очень важную роль в его функционировании и развитии.
Динамика фитоценозов
см. Динамика фитоценозов
Динамика фитоценозов
В связи с действием различных внешних и внутренних факторов, фитоценозы постоянно подвергаются различным изменениям. Но им присущ и процесс восстановления, который называется устойчивостью.
На этой странице материал по темам:
Эдификаторы в растительных сообществ примерах
Взаимоотношения растений в фитоценозе презентация
Фитоценозы тест
Искусственное растительное сообщество доклад реферат
Снижение конкуренции между растениями фитоценоза
Вопросы по этому материалу:
Дайте определение понятия «растительное сообщество». Приведите примеры естественных и искусственных растительных сообществ. Чем они отличаются?
Какие фитоценозы могут быть по видовому богатству?
Какие факторы влияют на видовой состав фитоценозов? Приведите примеры.
Охарактеризуйте понятия «виды-доминанты» и «эдификаторы». Приведите примеры таких видов в растительных сообществах.
Что такое ярусность фитоценоза?
Какое значение имеет ярусное сложение сообщества?

SCIRP Открытый доступ
Издательство научных исследований
Журналы от A до Z
Журналы по темам
- Биомедицинские и биологические науки.
- Бизнес и экономика
- Химия и материаловедение.
- Информатика. и общ.
- Науки о Земле и окружающей среде.
- Машиностроение
- Медицина и здравоохранение
- Физика и математика
- Социальные науки. и гуманитарные науки
Журналы по тематике
- Биомедицина и науки о жизни
- Бизнес и экономика
- Химия и материаловедение
- Информатика и связь
- Науки о Земле и окружающей среде
- Машиностроение
- Медицина и здравоохранение
- Физика и математика
- Социальные и гуманитарные науки
Публикация у нас
- Представление статьи
- Информация для авторов
- Ресурсы для экспертной оценки
- Открытые специальные выпуски
- Заявление об открытом доступе
- Часто задаваемые вопросы
Публикуйте у нас
- Представление статьи
- Информация для авторов
- Открытые специальные выпуски
- Заявление об открытом доступе
- Часто задаваемые вопросы
Подпишитесь на SCIRP
Свяжитесь с нами
клиент@scirp. org org | |
| +86 18163351462 (WhatsApp) | |
| 1655362766 | |
| Публикация бумаги WeChat |
| Недавно опубликованные статьи |
| Недавно опубликованные статьи |
Подпишитесь на SCIRP
Свяжитесь с нами
клиент@scirp. org org | |
| +86 18163351462 (WhatsApp) | |
| 1655362766 | |
| Публикация бумаги WeChat |
Бесплатные информационные бюллетени SCIRP
Copyright © 2006-2023 Scientific Research Publishing Inc. Все права защищены.
ВершинаВозрастные изменения газонных фитоценозов различных видов
БИО Web of Conferences 24 , 00036 (2020)
Возрастные изменения газонных фитоценозов различных видов
Хусаинова Ирина *
РГП «Институт ботаники и фитоинтродукции», интродукционный отдел, 050040 Алматы, Казахстан
* Автор, ответственный за переписку: fhusi@yandex. ru
ru
несколько составных многокомпонентных и монокомпонентных газонных ценозов на основе анализа их густоты и декоративности. Для обеспечения наилучшей устойчивости, проективного покрытия и декоративности важно при выборе компонентов и составлении травосмесей учитывать их межвидовую динамику развития, темп развития и долговечность. Оптимальные ценозы получаются при объединении видов из корневищно-кустарниковой группы, медленно растущих, но более долгоживущих, таких как Festuca rubra L., Poa pratensis L. Декоративность таких ценозов около 7,5 баллов, плотность около 20000 шт/м 2 . С возрастом в сложных ценозах густота побегов увеличивается, в монотипных — снижается.
© The Authors, опубликовано EDP Sciences, 2020
Это статья с открытым доступом, распространяемая в соответствии с лицензией Creative Commons Attribution License 4.0, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии, что оригинал работа цитируется правильно.
1 Введение
На первых этапах научного обоснования при ассортиментном отборе газонных ценозов лучшими считались монокультурные ценозы. Постепенно исследователи пришли к выводу, что использование травосмесей включает в себя преимущество, так как увеличивает спектр генетического разнообразия и повышает адаптационный потенциал культур фитоценозов. В травосмесях увеличивают проективное покрытие, уменьшают количество сорняков и снижают материальные затраты на их содержание. [1,2]. Потребление травяных смесей также призвано компенсировать разницу климатических условий по годам. Наличие в травостое злаковых видов с различными экологическими характеристиками должно повышать приспособленность травостоя к меняющимся влажностным и термическим условиям, которые сильно варьируют по годам. Доказано также преимущество смесей по сравнению с одновидовыми культурами в снижении вероятности возникновения болезней и их вредоносности [3]. Для определения процентного соотношения отдельных групп газонных трав в травосмесях, доминирования видов в Большое значение имеют определенные почвенно-климатические условия, скорость их развития и продолжительность жизни, ритмичность развития в течение вегетационного периода, межвидовая динамика компонентов фитоценоза во времени [4]. Эти особенности широко используются при моделировании травосмесей для газонов. Один из самых распространенных вариантов – это сочетание видов, отличающихся скоростью роста и продолжительностью жизни. Большинство исследователей считают, что кущение трав считается одним из существенных показателей жизнеспособности того или иного вида, произрастающего в ценозе с другими видами. Максимальное кущение происходит при температурах, значительно более низких, чем оптимальные температуры роста побегов [5], а оптимальная температура для трав «холодного климата» составляет от 10 до 18 °С [6, 7].
Эти особенности широко используются при моделировании травосмесей для газонов. Один из самых распространенных вариантов – это сочетание видов, отличающихся скоростью роста и продолжительностью жизни. Большинство исследователей считают, что кущение трав считается одним из существенных показателей жизнеспособности того или иного вида, произрастающего в ценозе с другими видами. Максимальное кущение происходит при температурах, значительно более низких, чем оптимальные температуры роста побегов [5], а оптимальная температура для трав «холодного климата» составляет от 10 до 18 °С [6, 7].
Цель исследования — изучить связь различных видов трав, произрастающих в многокомпонентных газонных ценозах различного состава, и провести сравнение с однородными монокультурными травостоями на основе оценки их кустистости и декоративности.
2 Материалы и методы
2.1 Объект исследования
При создании искусственных газонных фитоценозов заранее намечают компоненты травосмесей с программированием доминантов. Нами составлено 4 варианта составных смесей и 3 варианта монокомпонентных: 1. Типа «Овсяница жевательная» – Festuca rubra ‘Boreal’ 40%, Lolium perenne ‘Score’ 30%, Poa pratensis ‘Balin’ 20%, F. rubra ‘Frida’ 10%; 2. Тип ‘Райграс’ – L. perenne ‘Score’ 30%, L. perenne ‘Raygaubek’ 30%, F. rubra ‘Boreal’ 20%, P. pratensis ‘Balin’ 20%; 3. Тип ‘Мятлик луговой’ – P. pratensis ‘Sobra’ 60 %, F. rubra ‘Boreal’ 20 %, F. rubra ‘Frida’ 10 %, L. perenne ‘Score’ 10 %; 4. F. rubra ‘Boreal’ 100%; 5. L. perenne «Счет»; 6. P. pratensis ‘Балин’ 100%. Порядок компонентов в составе травосмеси представлен с учетом процентного содержания их в травосмеси от большего к меньшему. В качестве быстрорастущего компонента использовали L. perenne. Не только обеспечивает высокое проективное покрытие в первый год посева, но и создает микроусловия для прорастания других трав. Учетные участки подвергались регулярным стрижкам (20-22 раза за сезон).
Нами составлено 4 варианта составных смесей и 3 варианта монокомпонентных: 1. Типа «Овсяница жевательная» – Festuca rubra ‘Boreal’ 40%, Lolium perenne ‘Score’ 30%, Poa pratensis ‘Balin’ 20%, F. rubra ‘Frida’ 10%; 2. Тип ‘Райграс’ – L. perenne ‘Score’ 30%, L. perenne ‘Raygaubek’ 30%, F. rubra ‘Boreal’ 20%, P. pratensis ‘Balin’ 20%; 3. Тип ‘Мятлик луговой’ – P. pratensis ‘Sobra’ 60 %, F. rubra ‘Boreal’ 20 %, F. rubra ‘Frida’ 10 %, L. perenne ‘Score’ 10 %; 4. F. rubra ‘Boreal’ 100%; 5. L. perenne «Счет»; 6. P. pratensis ‘Балин’ 100%. Порядок компонентов в составе травосмеси представлен с учетом процентного содержания их в травосмеси от большего к меньшему. В качестве быстрорастущего компонента использовали L. perenne. Не только обеспечивает высокое проективное покрытие в первый год посева, но и создает микроусловия для прорастания других трав. Учетные участки подвергались регулярным стрижкам (20-22 раза за сезон).
2.2 Методы исследования
Плотность побегов является одним из основных аспектов качества [8]. Доказано, что уровни визуального качества коррелируют с плотностью побегов [9]. Для определения количества побегов на единицу площади поверхности почвы, как одного из объективных показателей декоративно-механических качеств дернового покрытия, использовали каркас размером (0,1 х 0,1) м [10]. Учеты проводили дважды за сезон – в мае (в период активного кущения) и в сентябре (в период активизации осеннего кущения) в четырехкратной повторности. В рамках вышеизложенного подсчитывали общее количество побегов независимо от типа кущения, отдельно количество побегов каждого вида, входящего в состав травосмеси, количество зеленых и желтых листьев дважды за сезон в течение пяти лет По предлагаемому способу Л. П. Мыцик, видовое озеленение газона определяли с расстояния 5-7 метров и угла 15 градусов один раз в год посева [11], проективное же покрытие ежегодно два раза за сезон. Оценка качества дерна проводилась по шкале от 1 до 9. : 1 – плохой, 2 – удовлетворительный, 9 – идеальный, а также цвет, густота, восприимчивость к болезням, однородность, зимнее повреждение, жизнеспособность растений [12].
: 1 – плохой, 2 – удовлетворительный, 9 – идеальный, а также цвет, густота, восприимчивость к болезням, однородность, зимнее повреждение, жизнеспособность растений [12].
2.3 Статистические методы
Статистический анализ результатов наблюдений с учетом средних значений и коэффициента вариации проводили с использованием программы Microsoft Excel 2010; LSD (самое низкое значимое различие) использовалось для определения значимости различий между наборами данных [13].
3 результатов
Испытания L. perenne. в условиях засушливой степной зоны юго-востока Казахстана показал хорошую энергию прорастания и всхожесть в полевых условиях, быстрое развитие, отличную кустистость в первый год жизни. Господствующее положение L. perenne займет 13 лет, а затем постепенно начнут сменяться медленно развивающимися, но более долгоживущими видами (F. rubra, P. pratensis). F. rubra и P. pratensis по типу кущения относятся к корневищно-кустарниковой группе. Они образуют систему розеточных побегов кустовой природы, соединенных корневищами разной длины [14]. Образцы F. rubra различаются степенью образования вегетативных побегов и типом побегов, что учитывается при оценке декоративности биотипов. Отмечены значительные различия в густоте побегов между видами и между видами внутри вида [15]. Наиболее совершенны для многолетних дерновых покрытий корневищно-рыхлые кустарниковые злаки, такие как F. rubra, P. pratensis, . Со временем эти виды станут содоминантами. То есть их участие будет зависеть от климатических и почвенных условий закладываемого места эксперимента [16]. Субдоминантные виды составляют более 10 % числа побегов в травосмеси. Виды, составляющие менее 10%, классифицируются как комплементарные [4].
Они образуют систему розеточных побегов кустовой природы, соединенных корневищами разной длины [14]. Образцы F. rubra различаются степенью образования вегетативных побегов и типом побегов, что учитывается при оценке декоративности биотипов. Отмечены значительные различия в густоте побегов между видами и между видами внутри вида [15]. Наиболее совершенны для многолетних дерновых покрытий корневищно-рыхлые кустарниковые злаки, такие как F. rubra, P. pratensis, . Со временем эти виды станут содоминантами. То есть их участие будет зависеть от климатических и почвенных условий закладываемого места эксперимента [16]. Субдоминантные виды составляют более 10 % числа побегов в травосмеси. Виды, составляющие менее 10%, классифицируются как комплементарные [4].
Тип овсяницы жевательной. Основная доминанта смеси F. rubra представлена в этой травосмеси двумя сортами из subsp. rubra и подвид. коммутат. Эти два подвида отличаются не только особенностями кущения, образования вегетативных и генеративных побегов, семенной продуктивностью, габитусом, но и внешним видом [17]. Перспективное озеленение наступает на 30-й день с момента массовых всходов. Проективное покрытие – 83%. В табл. 1 представлена пятилетняя динамика развития травостоя в сложных травосмесях типа «Овсяница жевательная», «Райграс» и др. Тип «мятлик луговой». Изменения соотношения компонентов происходят постепенно в сторону уменьшения присутствия типчака и увеличения содержания мятлика и райграса. Изменения соотношения компонентов носят волнообразный характер с положительной тенденцией увеличения общей густоты травы на 20 % к пятому году жизни. Максимальное количество всходов овсяницы наблюдается на четвертом году жизни газона. Увеличение процента poa встречается только на пятом курсе. Наблюдается значительный разброс по содержанию всходов овсяницы (5350-18200 на м 2 ) и райграса (800-14050 на м 2 ). Представитель подвида rubra сорта ‘Boreal’ характеризуется наличием как плагиотропных, так и ортотропных побегов. Подвид commutata, , к которому относится сорт ‘Фрида’, не образует плагиотропных побегов и наращивает вегетативную массу исключительно за счет ортотропных побегов и листьев на них.
Перспективное озеленение наступает на 30-й день с момента массовых всходов. Проективное покрытие – 83%. В табл. 1 представлена пятилетняя динамика развития травостоя в сложных травосмесях типа «Овсяница жевательная», «Райграс» и др. Тип «мятлик луговой». Изменения соотношения компонентов происходят постепенно в сторону уменьшения присутствия типчака и увеличения содержания мятлика и райграса. Изменения соотношения компонентов носят волнообразный характер с положительной тенденцией увеличения общей густоты травы на 20 % к пятому году жизни. Максимальное количество всходов овсяницы наблюдается на четвертом году жизни газона. Увеличение процента poa встречается только на пятом курсе. Наблюдается значительный разброс по содержанию всходов овсяницы (5350-18200 на м 2 ) и райграса (800-14050 на м 2 ). Представитель подвида rubra сорта ‘Boreal’ характеризуется наличием как плагиотропных, так и ортотропных побегов. Подвид commutata, , к которому относится сорт ‘Фрида’, не образует плагиотропных побегов и наращивает вегетативную массу исключительно за счет ортотропных побегов и листьев на них. Сочетание в ценозе растений с разным характером позволяет создать газон с лучшим проективным покрытием и декоративными качествами. P. pratensis действует как содоминант. В качестве субдоминанта – L. perenne. Несмотря на незначительное присутствие райграса в первый год посева, уже со второго года наблюдается увеличение содержания его побегов в траве с максимумом к третьему году.
Сочетание в ценозе растений с разным характером позволяет создать газон с лучшим проективным покрытием и декоративными качествами. P. pratensis действует как содоминант. В качестве субдоминанта – L. perenne. Несмотря на незначительное присутствие райграса в первый год посева, уже со второго года наблюдается увеличение содержания его побегов в траве с максимумом к третьему году.
Тип «райграс». Некоторые исследователи сообщают, что при посеве райграса в смеси с P. pratensis и Festuca arundinacea L, даже в небольшом процентном соотношении (около 10%), в конечном итоге, L. perenne доминировал, несмотря на эксперименты по уходу [3,18,19]. В других источниках отмечено уменьшение числа побегов райграса со второго года жизни как в чистых посевах, так и в смеси с другими видами [20]. По мнению А.А. Lapteva, L. perenne действует как временный доминант в первые 1-3 года жизни травостоя [4]. По нашему опыту, этот быстроразвивающийся вид представлен двумя сортами «Скор» и «Райгаубек». Перспективное озеленение – на 22 день, проективное покрытие – 77%. Наблюдается увеличение общей густоты травы на 28 % к пятому году жизни за счет увеличения содержания мятлик и овсяница . Максимальное количество всходов райграса наблюдается на второй год жизни газона, после чего его содержание постепенно снижается.
Перспективное озеленение – на 22 день, проективное покрытие – 77%. Наблюдается увеличение общей густоты травы на 28 % к пятому году жизни за счет увеличения содержания мятлик и овсяница . Максимальное количество всходов райграса наблюдается на второй год жизни газона, после чего его содержание постепенно снижается.
Тип «Мятлик луговой». P. pratensis будет доминировать, две разновидности F. rubra выступают здесь как субдоминанты, а L. perenne присутствует как комплементарный вид для более быстрого получения зеленого фона. Перспективное озеленение – на 30-е сутки, проективное покрытие – 80%. Изменения соотношения компонентов происходят постепенно в сторону уменьшения присутствия райграса и овсяницы и плавного увеличения Poa содержание с тенденцией к увеличению общей плотности на 19% к пятому году жизни. Максимальное количество побегов мятлика наблюдается на четвертый год жизни газона, после чего его содержание постепенно снижается.
Монокультура F. rubra ‘Boreal’. Медленно развивающийся травостой. Перспективное озеленение – на 35-е сутки, проективное покрытие 76%. В табл. 2 представлена пятилетняя динамика развития монокультурных культур «Бореал», «Счет», «Балин». В варианте 5 максимальная кустистость отмечена на второй год испытания 22000 побегов/м 2 . Общая тенденция – снижение плотности травяного покрова на 15% к пятому году жизни.
Монокультура L. perenne «Счет». Быстрорастущий травостой. Перспективное озеленение – на 26 день, проективное покрытие – 74%. Изменения количества побегов райграса носят волнообразный характер независимо от времени года от 14400 до 28100 побегов/м 2 . Максимальное кущение отмечено в первый год испытания 28100 побегов/м 2 . Общая тенденция полноты травостоя – снижение на 53 % к пятому году жизни.
Монокультура P. pratensis ‘Balin’ Медленнорастущий травостой. Перспективное озеленение – на 35 день, проективное покрытие 80%.
В этом варианте максимальная кустистость отмечена на четвертый год – 14450 побегов/м 2 . Снижение густоты травостоя к пятому году на 2%. Так, при создании чистых газонов из медленнорастущих трав P. pratensis, F. rubra, травостои развиваются медленно, в том числе и с низким проективным покрытием в первый год, поэтому в этот период активно заселяются сорняками, в результате что их содержание удорожается и теряется декоративность. Сравнивая данные между многокомпонентными травосмесями и монокультурой, можно увидеть разницу в начале озеленения. У сортов овсяница жевательная и мятлик луговой это на пять дней раньше, чем у чистых посевов, у типа райграс — на 4 дня раньше.
Таблица 1
Изменения состава травостоя в течение пяти лет жизни и основные параметры образцов
Таблица 2
Изменения густоты монотипных травостоев в течение пяти лет жизни и их сравнение с аналогичными многокомпонентными
4 Обсуждение
Из таблиц видно, что динамика изменения содержания медленнорастущих компонентов типа F. rubra и P. pratensis происходит очень плавно. Так, в травостое овсяницы жевательной преобладают F. rubra с первого года жизни, пик его присутствия наблюдается только на четвертом году жизни. На четвертый год также наблюдается максимальное присутствие P. pratensis в травостое типа ‘Мятлик луговой’. Количество побегов Festuca, Poa, Lolium сильно колеблется от весны к осени, от года к году во всех испытанных травосмесях. Каждый искусственно созданный фитоценоз представляет собой пример сложной, подвижной во времени и пространстве, но достаточно гибкой системы. При совместном произрастании видов из разных родов, с различными жизненными формами и различной скоростью развития в онтогенезе в режиме газонного ценоза в условиях конкуренции и симбиоза на декоративность влияют густота посадки, условия агротехнического ухода, состав травы. смесь. Степень поражения листьев может свидетельствовать о состоянии того или иного ценоза. Сравнение тенденций побегообразования сложных и монотипных ценозов выявляет устойчивое увеличение густоты побегов к пятому году жизни у овсяницы жевательной на 20 %, у райграсного на 27 %, у мятлика лугового.
rubra и P. pratensis происходит очень плавно. Так, в травостое овсяницы жевательной преобладают F. rubra с первого года жизни, пик его присутствия наблюдается только на четвертом году жизни. На четвертый год также наблюдается максимальное присутствие P. pratensis в травостое типа ‘Мятлик луговой’. Количество побегов Festuca, Poa, Lolium сильно колеблется от весны к осени, от года к году во всех испытанных травосмесях. Каждый искусственно созданный фитоценоз представляет собой пример сложной, подвижной во времени и пространстве, но достаточно гибкой системы. При совместном произрастании видов из разных родов, с различными жизненными формами и различной скоростью развития в онтогенезе в режиме газонного ценоза в условиях конкуренции и симбиоза на декоративность влияют густота посадки, условия агротехнического ухода, состав травы. смесь. Степень поражения листьев может свидетельствовать о состоянии того или иного ценоза. Сравнение тенденций побегообразования сложных и монотипных ценозов выявляет устойчивое увеличение густоты побегов к пятому году жизни у овсяницы жевательной на 20 %, у райграсного на 27 %, у мятлика лугового. на 21%. В то время как в монотипных злаковых насаждениях наблюдается постепенное снижение плотности: у мятлика лугового (2%), овсяницы жевательной, овсяницы (15%) и в большей степени райграса (53%). Максимальная густота побегов, проективное покрытие и высокая декоративность у сортов овсяницы жевательной и мятлика лугового (7,3 и 6,9, соответственно) происходят из-за присутствия двух подвидов F. rubra. Таким образом, сложные культурные фитоценозы, состоящие из наиболее совместимых компонентов, формируют более плотную надземную массу по сравнению с чистыми культурами, что согласуется с выводами других ученых. Последние нежелательно использовать для создания экстенсивных газонных покрытий из-за их неустойчивости к неблагоприятным климатическим условиям, болезням и низкой декоративности.
на 21%. В то время как в монотипных злаковых насаждениях наблюдается постепенное снижение плотности: у мятлика лугового (2%), овсяницы жевательной, овсяницы (15%) и в большей степени райграса (53%). Максимальная густота побегов, проективное покрытие и высокая декоративность у сортов овсяницы жевательной и мятлика лугового (7,3 и 6,9, соответственно) происходят из-за присутствия двух подвидов F. rubra. Таким образом, сложные культурные фитоценозы, состоящие из наиболее совместимых компонентов, формируют более плотную надземную массу по сравнению с чистыми культурами, что согласуется с выводами других ученых. Последние нежелательно использовать для создания экстенсивных газонных покрытий из-за их неустойчивости к неблагоприятным климатическим условиям, болезням и низкой декоративности.
Каталожные номера
- В.А. Тюльдюков, И.В. Кобозев, Н.В. Парахин, Газонокосилки и благоустройство заселенных территорий. М., 2002. [Google Scholar]
- СМ. Дональд, зав. Агрон. 15, 1-118 (1963) [Перекрестная ссылка]
[Google Scholar]
- Дж. Х. Данн, Э.Х. Эрвин, Б.С. Фрезенбург, Хорт. Наука. 37, 214-217 (2002) [Перекрестная ссылка] [Google Scholar]
- А.А. Лаптев, Дерновые травы (Киев, 1983) [Google Scholar]
- Дж. Б. Берд, Наука и культура газонов. Prentice Hall, Engelwood Cliffs, NJ, 658 (1973)
[Google Scholar]
- Да Коста, Б. Хуанг, Физиология теплового стресса и управление им. Газонные травы: биология, использование и управление (Agron. Monogr. ASA, CSSA и SSSA. Мэдисон, Висконсин, 2013 г.) [Google Scholar]
- X. Лю, Б. Хуанг, Environ. Эксп. Бот. 53, 233-245 (2005) [Google Scholar]
- В. Боллер, Великобритания Посселт, Кормовые культуры и декоративные травы. Справочник по селекции растений. 5, 552 (2010)
[Google Scholar]
- А.Х.Брюно, А.Дж. Ньюэлл и F.M.E. Кроссли, J. Turfgrass Sci. 76, 63-69(2000) [Google Scholar]
- Смелов С.П. Установление объективных показателей качества газона на аэродромах. 5, 65 (Москва, 1943) [Google Scholar]
- Мыцык Л. П. Эколого-биологические основы создания газонных насаждений в степной зоне (кандидатская диссертация по биологическим наукам, Днепропетровский государственный университет, 1992)
[Google Scholar]
- Национальная программа оценки дерновых трав, Национальное испытание пастбищного райграса 1994. (NTEP Prog. Rep. USDA-ARS, Beltsville, MD, 1998) [Google Scholar]
- Л.Г. Рязанова, А.В. Проворченко, И.В. Горбунов, Основы статистического анализа результатов исследований в садоводстве (Кубанский ГАУ, Краснодар, 2013) [Google Scholar]
- Т. И. Серебрякова, Морфогенез побегов и эволюция жизненных форм злаков (Наука, М., 1971).
[Google Scholar]
- М.Дж. Гудинг, А.Дж. Ньюэлл, Дж. Спортс Терф Рез. Инст. 66, 141-148 (1990) [Google Scholar]
- Т.А. Работнов, Фитозенология (М., 1978). [Google Scholar]
- А. Квальбейн, Т.С. Амлид, Путеводитель по травам (Landvik, исследовательская группа NIBIO по газонным травам, 2015 г.)
[Google Scholar]
- К.Л. Хант, Дж. Х. Данн, Агрон Дж. 85, 211–215 (1993) [Google Scholar]
- T. Hsiang, K. Carey, B. He, J.E. Eggens, Intl. Газон Соц. рез. J. 8, 671-679 (1997) [Google Scholar]
- А.
 Дональд, зав. Агрон. 15, 1-118 (1963) [Перекрестная ссылка]
[Google Scholar]
Дональд, зав. Агрон. 15, 1-118 (1963) [Перекрестная ссылка]
[Google Scholar] Б. Берд, Наука и культура газонов. Prentice Hall, Engelwood Cliffs, NJ, 658 (1973)
[Google Scholar]
Б. Берд, Наука и культура газонов. Prentice Hall, Engelwood Cliffs, NJ, 658 (1973)
[Google Scholar] Боллер, Великобритания Посселт, Кормовые культуры и декоративные травы. Справочник по селекции растений. 5, 552 (2010)
[Google Scholar]
Боллер, Великобритания Посселт, Кормовые культуры и декоративные травы. Справочник по селекции растений. 5, 552 (2010)
[Google Scholar] П. Эколого-биологические основы создания газонных насаждений в степной зоне (кандидатская диссертация по биологическим наукам, Днепропетровский государственный университет, 1992)
[Google Scholar]
П. Эколого-биологические основы создания газонных насаждений в степной зоне (кандидатская диссертация по биологическим наукам, Днепропетровский государственный университет, 1992)
[Google Scholar] И. Серебрякова, Морфогенез побегов и эволюция жизненных форм злаков (Наука, М., 1971).
[Google Scholar]
И. Серебрякова, Морфогенез побегов и эволюция жизненных форм злаков (Наука, М., 1971).
[Google Scholar] Квальбейн, Т.С. Амлид, Путеводитель по травам (Landvik, исследовательская группа NIBIO по газонным травам, 2015 г.)
[Google Scholar]
Квальбейн, Т.С. Амлид, Путеводитель по травам (Landvik, исследовательская группа NIBIO по газонным травам, 2015 г.)
[Google Scholar]