Проблемы Эволюции
Проблемы Эволюции
Предыдущая глава Следующая глава Оглавление
Доказательства эволюции
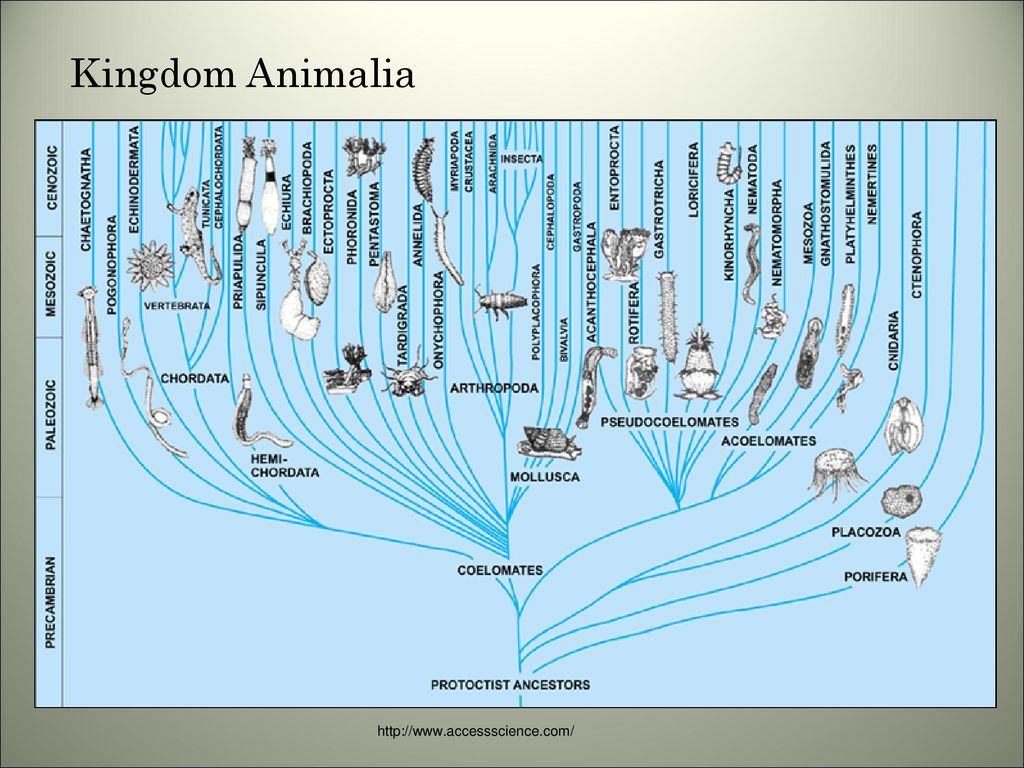
2. Эволюционное дерево
Филогенетическое дерево с указанием размера геномов.
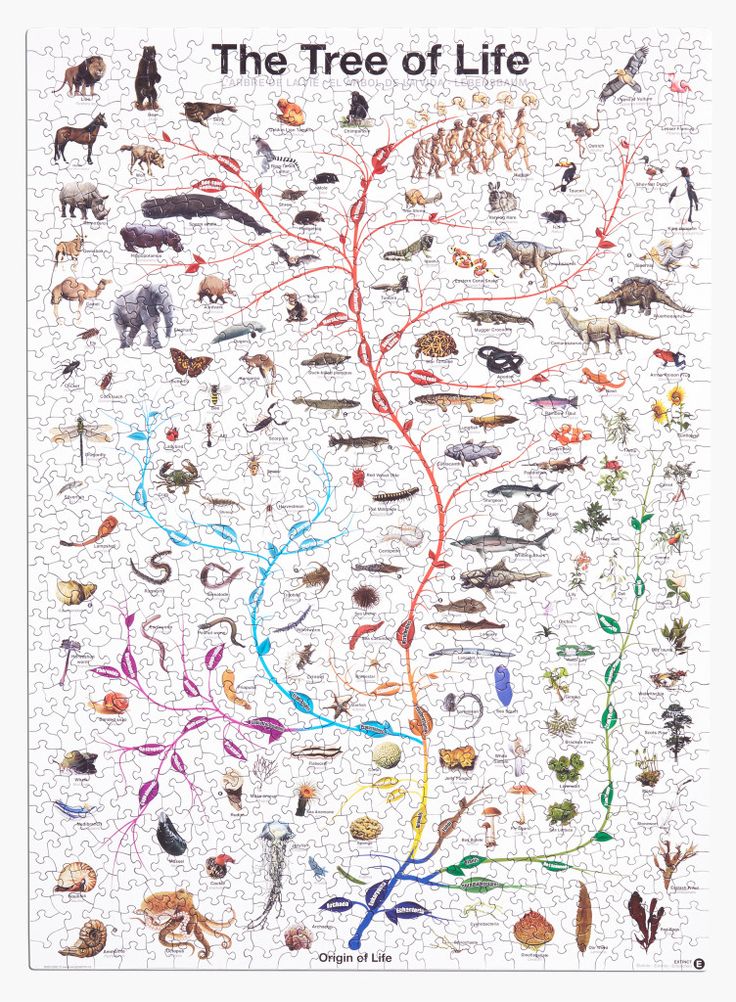
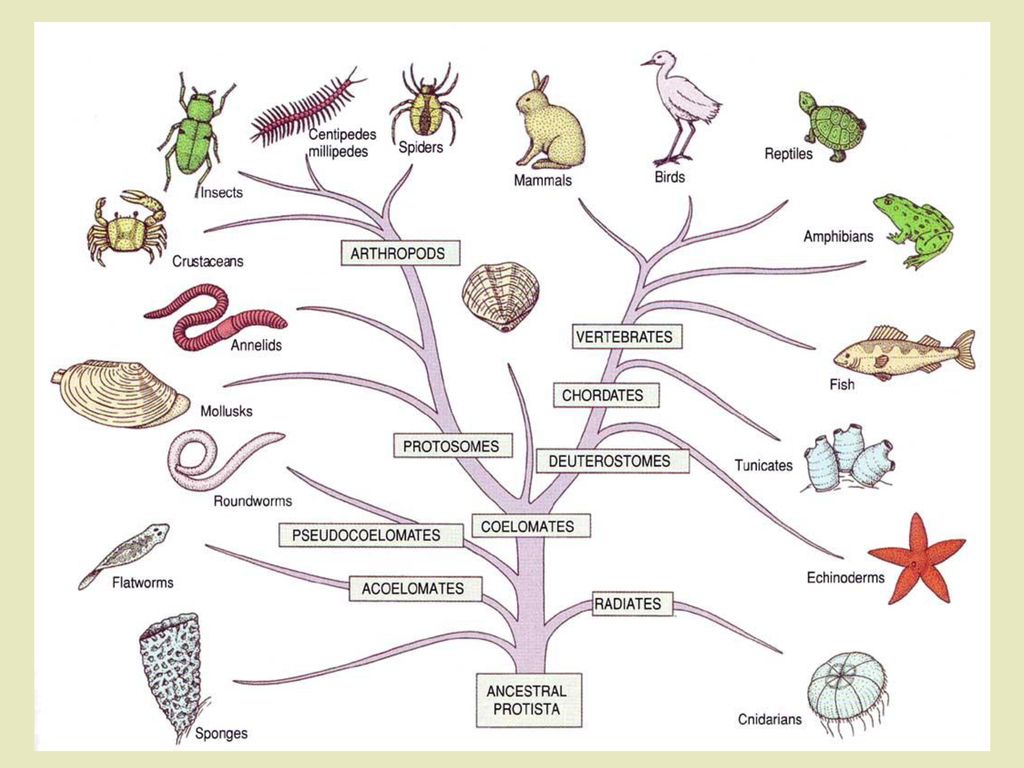
Классификация живых организмов представляет собой многоуровневую иерархическую структуру: организмы делятся на царства, царства делятся на типы, типы — на классы, классы — на отряды, и так далее. В результате такого ветвления получается филогенетическое дерево. Наличие единственной (естественной) классификации означает, что существует объективная закономерность в основе этой классификации.Именно такой результат можно ожидать при эволюционном происхождении животных от общего предка.Ветвление филогенетического дерева соответствует делению популяций в процессе видообразования.
Несмотря на многочисленные разногласия между биологами по поводу отнесения тех или иных видов к конкретным группам (таксонам), эти противоречия имеют частный характер. Практика показывает, что биологические классификации, построенные на основе разных признаков (морфологических, эмбриологических, биохимических или генетических) в тенденции стремятся к одной и той же древовидной иерархической схеме — естественной классификации, отражающей последовательность расхождения эволюционных линий. Чем больше признаков учитывается в ходе классификации, тем выше сходство получаемых деревьев. Наличие естественной классификации было очевидно биологам еще в додарвиновские времена, и это изначально трактовалось как свидетельство иерархической организации замысла Творца. Однако в разнообразии других природных объектов, которые, в отличие от живых организмов, не происходят от общего предка, отсутствует единая древовидная иерархическая структура. Классификация таких объектов либо получается принципиально различной при использовании разных наборов признаков (например, минералы), либо имеет принципиально не «древесный» вид (например, химические элементы, звезды).
Если бы была верна концепция «бараминов», естественная классификация и филогенетические деревья, получаемые на основе различных признаков и статистических методов, должны были бы иметь весьма характерный вид: изолированные пучки коротких ветвей (барамины) должны были бы отделяться друг от друга очень длинными (точнее, имеющими абсолютно произвольную длину) ветвями. Этого не наблюдается. Длины ветвей филогенетического дерева (отражающие, в первом приближении, степень различий по используемым признакам) в среднем примерно одинаковы между видами одного рода, родами одного семейства, семействами одного отряда и т.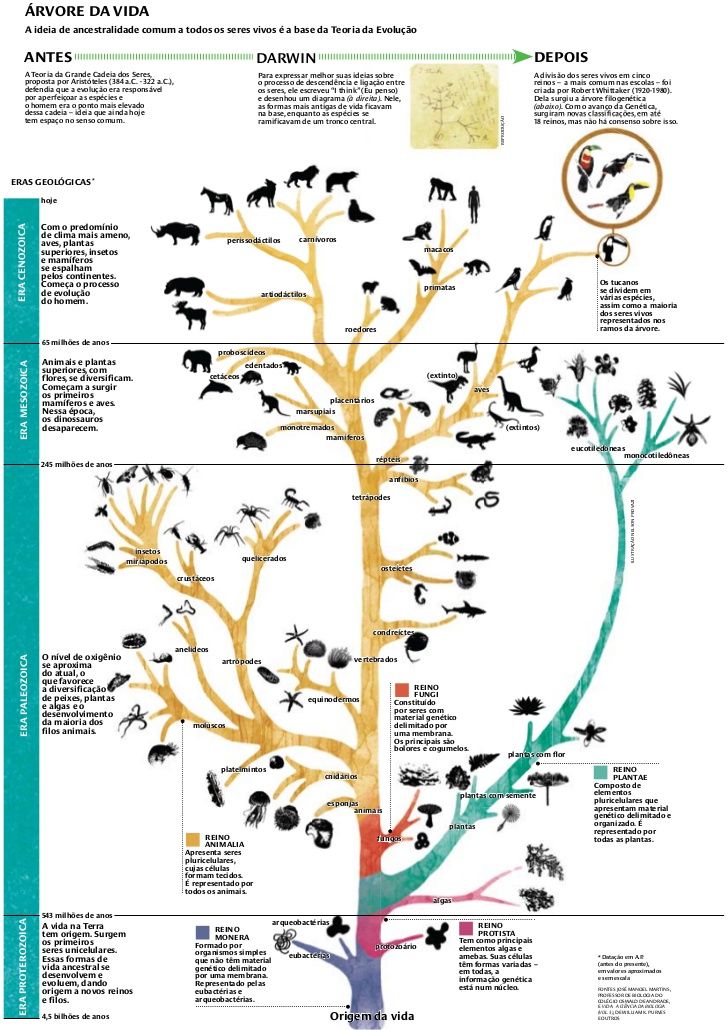
Кроме того, если бы в идее «бараминов» было хоть какое-то рациональное зерно, этого не могли бы не заметить специалисты по систематике животных и растений еще задолго до Дарвина. Они бы обязательно увидели, что один из надвидовых таксономических рангов (род, отряд или семейство — антиэволюционисты обычно утверждают, что их «барамины» соответствуют каким-то из этих рангов) радикально отличается от всех остальных. Например, они бы увидели, что животный мир четко делится на семейства, которые поэтому являются особым, «естественным» уровнем классификации (и соответствуют «богосотворенным родам»), и гораздо менее четко делится на роды, подсемейства, надсемейства, отряды, подклассы и т. д., которые никаким «богосотворенным родам» не соответствуют. В действительности все ранги биологической классификации имеют примерно одинаковую степень как «четкости», так и «расплывчатости». По-настоящему «естественной» единицей классификации безоговорочно признается лишь вид (по критерию полного или почти полного отсутствия скрещиваний с другими видами в природе), но барамин по определению — это не вид, а нечто большее. Однако никакого надвидового ранга, более «естественного», чем остальные, в природе не существует.
д., которые никаким «богосотворенным родам» не соответствуют. В действительности все ранги биологической классификации имеют примерно одинаковую степень как «четкости», так и «расплывчатости». По-настоящему «естественной» единицей классификации безоговорочно признается лишь вид (по критерию полного или почти полного отсутствия скрещиваний с другими видами в природе), но барамин по определению — это не вид, а нечто большее. Однако никакого надвидового ранга, более «естественного», чем остальные, в природе не существует.
Существуют различные статистические методы для точной оценки того, насколько объекты с разными свойствами укладываются в ту или иную иерархию. Эти методы измеряют так называемый «филогенетический сигнал», позволяя отличить мнимые закономерности от объективных. Например, у любого «генеалогического древа» автомобилей будет низкий уровень филогенетического сигнала. У эволюционного дерева и у различных его частей, напротив, стабильно четкий филогенетический сигнал.
Есть несколько источников данных, на основе которых можно делать выводы о степени родства между видами. Если существует единое эволюционное дерево, объединяющее все виды в объективную генеалогию, то все данные должны подтверждать эту генеалогию. Филогенетические деревья, построенные независимо, должны соответствовать друг другу. Наиболее простой, хотя и не очень точный способ построить такое дерево — сравнение строения организмов животных: чем меньше различий между видами, тем меньше поколений отделяют их от общего предка. Палеонтологическая летопись, наряду с другими подходами, позволяет установить родство между классами животных. Например, найденные останки пернатых динозавров свидетельствуют о родстве между рептилиями и птицами. Биогеография и эмбриология также дают информацию об эволюционной близости видов. Наиболее точный источник данных, недоступный во времена Дарвина — сравнительный анализ геномов различных организмов. Эволюционное дерево можно построить по каждому отдельно взятому гену, также исследователи рассматривают всевозможные некодирующие последовательности.
Все эти источники информации дают одинаковую картину с точностью до погрешности используемых методов. Тот факт, что эволюционные деревья, построенные по разным данным, соответствуют друг другу, элементарно объясняется эволюционной теорией. Другие объяснения отсутствуют: например, нет никаких причин, почему организмы, сходные по строению, должны иметь сходные некодирующие последовательности в геноме (интроны в одних и тех же местах генов, остатки встроившихся ретровирусов в одних и тех же местах генома и т.д.), если они не произошли от одного предка.
Примеры биологических исследований, связанных с построением эволюционных деревьев:
1) Новые данные позволили уточнить родословную животного царства
2) Невероятное разнообразие жуков получило эволюционное объяснение
3) Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных
4) Найдено «недостающее звено» между членистоногими и загадочными кембрийскими чудовищами аномалокарисами
5) Доказан скачкообразный характер эволюционного процесса
Горизонтальный перенос генов, межвидовая гибридизация и симбиогенез порождают горизонтальные перемычки между некоторыми ветвями эволюционного дерева, особенно в его «нижней» части — у прокариот. Это, однако, не разрушает его общую древовидную (иерархическую) структуру, потому что вертикальная передача генов (от родителей к потомкам) у всех живых организмов резко преобладает над горизонтальной.
Это, однако, не разрушает его общую древовидную (иерархическую) структуру, потому что вертикальная передача генов (от родителей к потомкам) у всех живых организмов резко преобладает над горизонтальной.
Лингвистическая эволюция как аналог биологической
Эволюция языков отчасти похожа на эволюцию биологических видов, хотя это сходство не следует преувеличивать. Лингвистическая эволюция в основном нейтральна, т.е. не имеет приспособительного характера (нельзя утверждать, например, что грамматика чукотского языка более приспособлена к холодному климату, чем грамматика африканских языков). В биологической эволюции, впрочем, тоже велик элемент нейтральности. Построение эволюционных деревьев в биологии осуществляется чаще всего на основе именно нейтральных признаков (потому что приспособительные признаки — менее надежное свидетельство родства; они часто могут возникать у неродственных форм в сходных условиях в результате одинаковой направленности отбора). Специалисты по исторической лингвистике активно и весьма успешно используют математические методы построения эволюционных деревьев, разработанные биологами (подробнее см. в заметке Лингвистическая эволюция сходна с биологической).
Специалисты по исторической лингвистике активно и весьма успешно используют математические методы построения эволюционных деревьев, разработанные биологами (подробнее см. в заметке Лингвистическая эволюция сходна с биологической).
Аналогия с эволюцией языков полезна нам прежде всего потому, что она помогает лучше понять два ключевых свойства биологической эволюции: ее постепенность и относительную дискретность видов.
Постепенность. Языки, как и биологические виды, эволюционируют путем накопления мелких изменений. Один язык (например, латынь) превращается в другой (например, итальянский) не сразу, а постепенно. Каждое следующее поколение говорит лишь немного иначе, чем предыдущее. Не бывает так, чтобы родители, говорящие, к примеру, на латыни, родили детей, которые, научившись говорить, вдруг заговорили по-итальянски. Так же и в эволюции видов: родители одного вида не могут родить детенышей, относящихся уже к другому виду.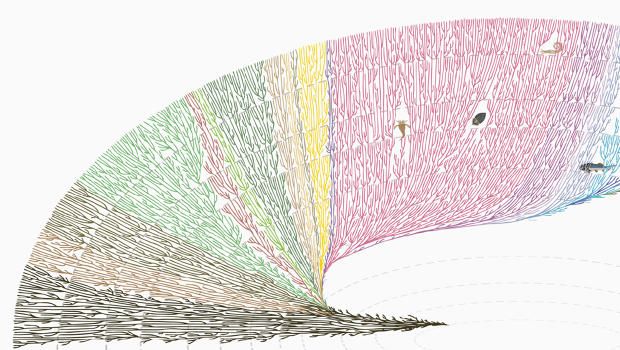
Дискретность видов. Несмотря на множество наблюдаемых в природе случаев плавных переходов между видами, разнообразных гибридных зон, кольцевых видов и т.п. (см. в разделе «Наблюдаемая эволюция»), большинство существующих видов все-таки достаточно дискретны, т.е. имеют довольно четкие границы. Обычно мы можем однозначно определить видовую принадлежность животного или растения. Между большинством видов есть заметные «разрывы» (хиатусы) в пространстве признаков. Антиэволюционисты иногда ставят это в упрек эволюционному учению, утверждая, что если бы эволюция действительно происходила, мы не должны были бы наблюдать никакой дискретности в видовом разнообразии, а только сплошные плавные переходы.
Наблюдаемая дискретность видов во многом определяется дискретностью экологических условий и пониженной приспособленностью промежуточных форм. Например, в тайге условия одни, среди арктических льдов — другие, к первым условиям лучше приспособлен бурый медведь, ко вторым — белый. Промежуточные формы в обоих биотопах будут проигрывать в конкурентной борьбе живущим там специалистам. О таком механизме формирования дискретности, основанном на пониженной конкурентоспособности промежуточных форм, писал еще Дарвин. Дополнительное объяснение дискретности можно получить из аналогии с языками.
Например, в тайге условия одни, среди арктических льдов — другие, к первым условиям лучше приспособлен бурый медведь, ко вторым — белый. Промежуточные формы в обоих биотопах будут проигрывать в конкурентной борьбе живущим там специалистам. О таком механизме формирования дискретности, основанном на пониженной конкурентоспособности промежуточных форм, писал еще Дарвин. Дополнительное объяснение дискретности можно получить из аналогии с языками.
Языки тоже в основном дискретны. Конечно, в зонах смешения разноязычных народов иногда формируются гибридные диалекты (аналог гибридных зон в биологии), но все-таки это не типично. Большинство людей говорит на вполне определенном языке, а не на каких-то смешанных диалектах. Главная причина дискретности в биологии и лингвистике, по-видимому, одна и та же. Человеку выгодно уметь свободно и полноценно обмениваться информацией с достаточно большой популяцией себе подобных. Говорить на смеси русского и немецкого очень неудобно: плохо будут понимать и русские, и немцы.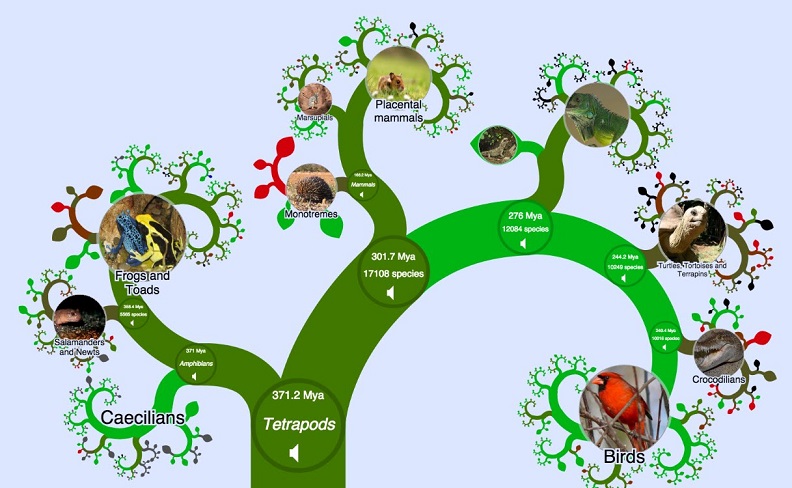 Живым организмам точно так же выгодно уметь свободно и полноценно обмениваться генами (смешивая их в потомстве) с достаточно многочисленной группой особей. В пределах каждого генофонда гены под действием отбора становятся «притертыми», приспособленными друг к другу. Иметь в своем геноме смесь генов, приспособленных к разным генофондам, в общем случае невыгодно, потому что потомство такого организма от скрещивания с любой из «чистых» форм, скорее всего, будет иметь пониженную жизнеспособность. Необходимость (выгодность) свободного обмена информацией (словесной или генетической) в пределах достаточно больших популяций в тенденции приводит к формрованию и поддержанию наблюдаемой дискретности. В природных условиях довольно часто возникают межвидовые гибриды, но число их, как правило, остается небольшим, а их потомство в будущем может постепенно распасться на практически «чистых» представителей двух исходных видов. Попадая в генофонд вида А, смешанный набор генов гибридной особи будет под действием отбора постепенно очищен от «посторонних примесей», так что в итоге в генофонде останутся только гены, хорошо «приспособленные» именно к этому генофонду.
Живым организмам точно так же выгодно уметь свободно и полноценно обмениваться генами (смешивая их в потомстве) с достаточно многочисленной группой особей. В пределах каждого генофонда гены под действием отбора становятся «притертыми», приспособленными друг к другу. Иметь в своем геноме смесь генов, приспособленных к разным генофондам, в общем случае невыгодно, потому что потомство такого организма от скрещивания с любой из «чистых» форм, скорее всего, будет иметь пониженную жизнеспособность. Необходимость (выгодность) свободного обмена информацией (словесной или генетической) в пределах достаточно больших популяций в тенденции приводит к формрованию и поддержанию наблюдаемой дискретности. В природных условиях довольно часто возникают межвидовые гибриды, но число их, как правило, остается небольшим, а их потомство в будущем может постепенно распасться на практически «чистых» представителей двух исходных видов. Попадая в генофонд вида А, смешанный набор генов гибридной особи будет под действием отбора постепенно очищен от «посторонних примесей», так что в итоге в генофонде останутся только гены, хорошо «приспособленные» именно к этому генофонду.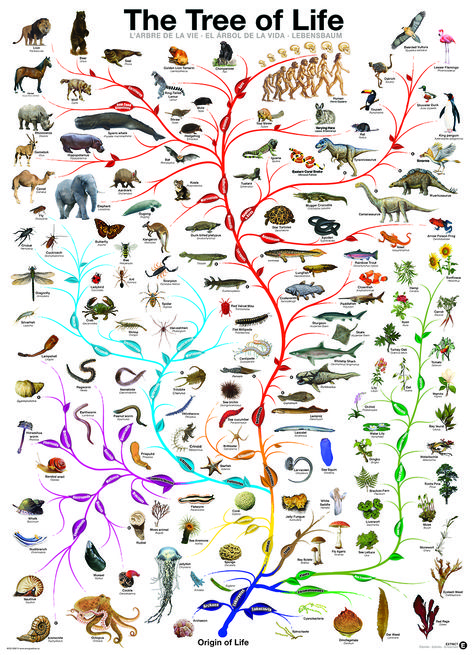 То же самое произойдет и с теми генами гибрида, которые будут подвергаться отбору в пределах генофонда Б. Именно поэтому межвидовая гибридизация, как правило, не приводит к слиянию двух разошедшихся видов в один.
То же самое произойдет и с теми генами гибрида, которые будут подвергаться отбору в пределах генофонда Б. Именно поэтому межвидовая гибридизация, как правило, не приводит к слиянию двух разошедшихся видов в один.
Предыдущая глава Следующая глава Оглавление
Как реконструируют ход эволюции? / Хабр
Ход эволюции изображают в виде деревьев
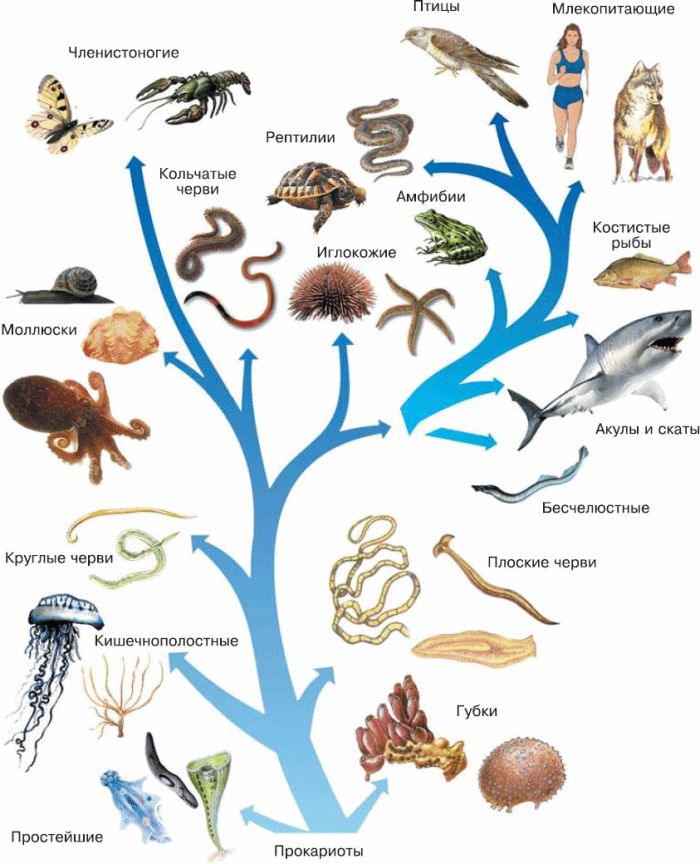
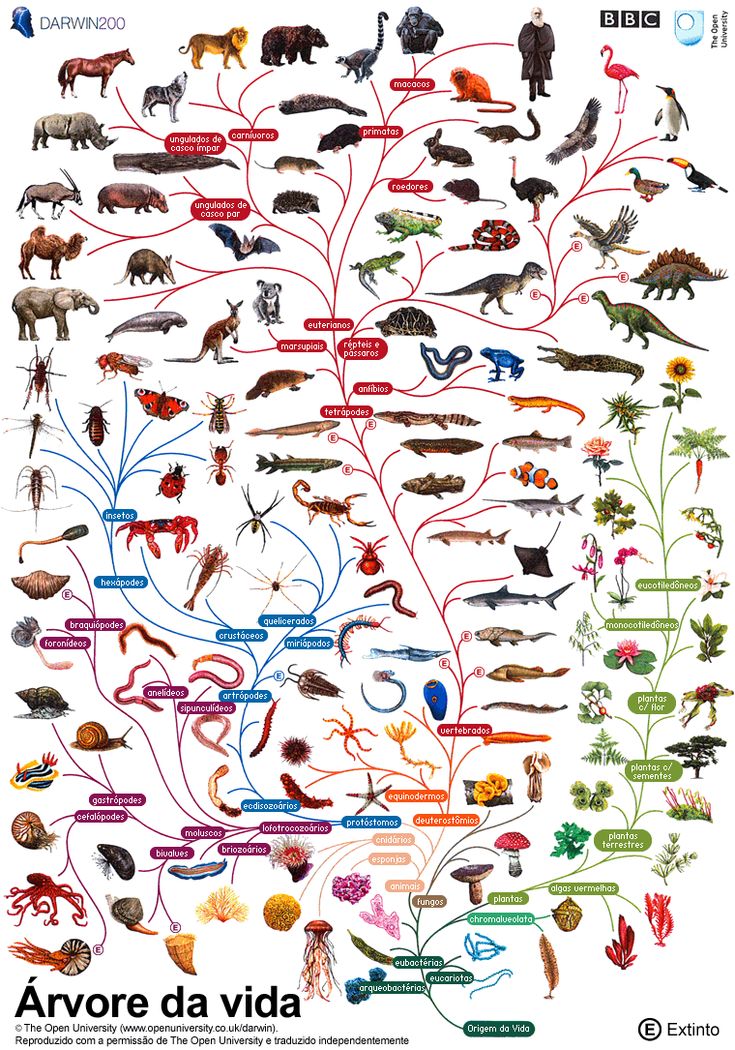
Важнейшей задачей биологии является реконструкция хода эволюции. Данных палеонтологии катастрофически не хватает, а все, что у нас есть кроме нее, это ныне живые организмы. Пути эволюции изображают в виде деревьев жизни, или филогентических деревьев, показывающих, в каком порядке разошлись эволюционные пути различных организмов. Филогенетическими они называются потому, что процесс эволюционной истории организма называется филогенезом.
Сейчас деревья имеют множество важных применений.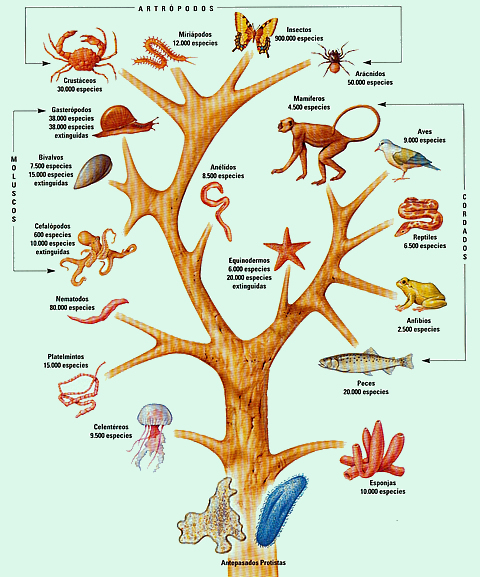 Как чисто фундаментального характера, например, чтобы узнать особенности строения организмов, живших миллиарды лет назад. Так и более прикладных. Они используются при сборках геномов, поиске генов, важных для патогенности возбудителей болезней, и многого другого.
Как чисто фундаментального характера, например, чтобы узнать особенности строения организмов, живших миллиарды лет назад. Так и более прикладных. Они используются при сборках геномов, поиске генов, важных для патогенности возбудителей болезней, и многого другого.
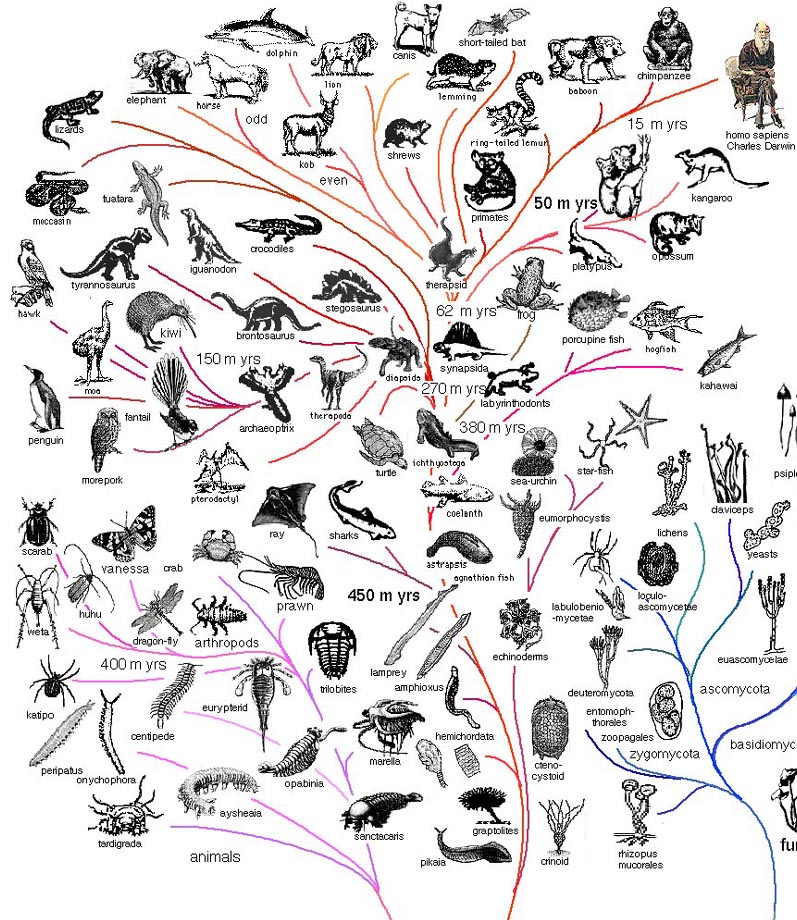
Как и многое в эволюционной биологии, первое филогенетическое дерево построил Дарвин, когда путешествовал на корабле Бигль (на картинке именно оно), однако, тогда это была лишь концепция, иллюстрировавшая идеи о том, что в ходе эволюции один вид разделялся на несколько.
Как понять, какое дерево нам подходит?
Допустим, у нас есть 4 организма. Корова, свинья, человек и ящерица, и мы хотим узнать их эволюционную историю. Как понять, какое дерево нам нужно? Ведь можно построить несколько вариантов их эволюции.
Универсального ответа на этот вопрос нет. Один из ответов является местным вариантом бритвы Оккама. Нужно выбрать дерево, где одни и те же мутации возникали только один раз (или, наименьшее число раз). У каждого вида есть какие-то признаки, по которым его можно сравнить с другими. Например, можно рассмотреть наличие копыт. У свиньи и коровы копыта есть, а у человека и ящерицы, очевидно, нет. Есть несколько вариантов расположения этих четырех видов на дереве. Мы разберем всего два из них, но для большего числа деревьев принцип точно такой же.
У каждого вида есть какие-то признаки, по которым его можно сравнить с другими. Например, можно рассмотреть наличие копыт. У свиньи и коровы копыта есть, а у человека и ящерицы, очевидно, нет. Есть несколько вариантов расположения этих четырех видов на дереве. Мы разберем всего два из них, но для большего числа деревьев принцип точно такой же.
Допустим, мы построили два дерева и хотим узнать, какое из них лучше соответствует действительности.
Для первого дерева достаточно предположить, что копыта возникли один раз у общего предка свиньи и коровы. То есть достаточно было произойти одному изменению.
Для второго же дерева копыта либо были у общего предка всех четырех организмов и дважды, в линиях человека и ящерицы, утрачивались, тогда нужно, чтобы произошло два изменения для объяснения этой модели. Либо копыт у общего предка не было, и, тогда, они должны были возникнуть независимо в линиях коровы и свиньи, что, опять же, требует двух изменений.
Значит, мы считаем первое дерево лучше второго. Конечно, не всегда все так просто, и иногда есть несколько равносильных деревьев, и, сказать, какое из ни лучше, в рамках этого метода нельзя. Но, изучая не один признак, а множество, можно разрешить этот вопрос.
Конечно, не всегда все так просто, и иногда есть несколько равносильных деревьев, и, сказать, какое из ни лучше, в рамках этого метода нельзя. Но, изучая не один признак, а множество, можно разрешить этот вопрос.
Какие признаки брать и при чем тут ДНК?
Раньше деревья строили на основе особенностей развития организмов, внешнего и внутреннего строения и, позднее, биохимических особенностей, из-за чего они часто получались неточными, грубыми и, самое главное, сильно зависели от мнений и предпочтений, господствовавших в голове их создателей.
Сейчас имеется гораздо более надежный и объективный метод. Если мы посмотрим на молекулу ДНК, то мы увидим, что это полимерная цепь, состоящая из мономеров четырех типов, последовательность которых, как раз, и является генетическим кодом. Иначе говоря, ДНК можно представить как текст, написанный четырехбуквенным алфавитом (которые в биоинформатике называют A, T, G и C). И, каждую позицию на ДНК можно взять в качестве признака, который может принимать значения любой из четырех букв, или, быть выпавшим.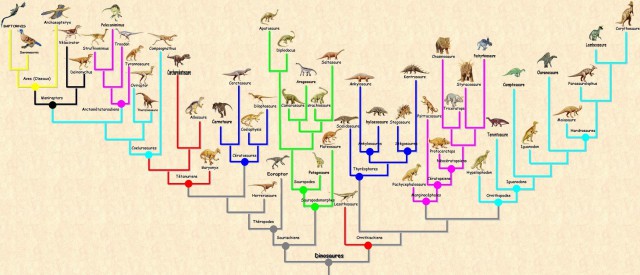
При каждом делении клетки происходит копирование этого текста и иногда при копировании происходят ошибки. Соответственно, чем больше раз тексты копировались независимо друг от друга, тем больше между ними различий. Однако, сравнивать полные геномы не имеет смысла из-за того, что постоянно встречаются такие мутации, которые невозможно описать современными алгоритмами постройки деревьев. Хромосома может разделиться на несколько, две могут сливаться в одну. Вирусы могут встраивать в ДНК клетки свой генетический материал. Гены могут удваиваться или теряться. Список подобных мутаций весьма обширен, так что обычно сравнивают небольшие участки ДНК, про которые известно, что в них происходили лишь небольшие мутации, такие, как замены единичных букв или небольшие удаления и вставки. Такими участками ДНК могут, например, быть одинаковые гены разных видов. Также часто строят деревья по белкам, но различия между методами построения по белкам и по ДНК чисто технические, и для понимания сути не важны.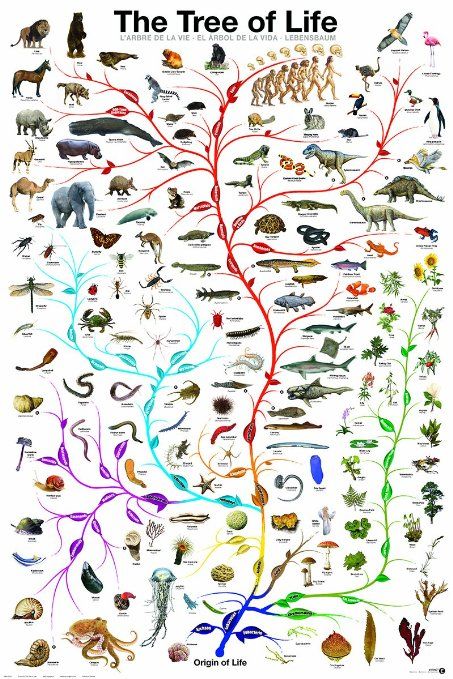 Деревья, отражающие эволюцию целых организмов, обычно, делают на основе множества таких деревьев, построенных по небольшим участкам ДНК, или, по отдельным генам, про которые заранее известно, что их эволюция совпадает с эволюцией организмов.
Деревья, отражающие эволюцию целых организмов, обычно, делают на основе множества таких деревьев, построенных по небольшим участкам ДНК, или, по отдельным генам, про которые заранее известно, что их эволюция совпадает с эволюцией организмов.
Как строят деревья?
Есть три принципиальных способа постройки дерева. Первый — это перебор всех возможных деревьев, которые можно построить для данных организмов. Не всех правдоподобных, а всех вообще. Но даже для дерева с двадцатью листьями такой перебор уже слишком сложен, а часто необходимо строить деревья намного большего размера. Вторая — это выращивание дерева. При таком подходе сначала строится маленькое простое дерево, а затем к нему добавляется по одной ветви так, чтобы каждый раз выходило хорошее дерево. И так, пока оно не будет построено полностью., и третье — это эвристические методы, с помощью которых строится какое-то правдоподобное дерево по определенному алгоритму. Описанный ранее метод оценки качества дерева, как раз и нужен, например, для ответа на вопрос, какое из построенных деревьев лучше, или, куда поставить новую веточку при выращивании дерева.
В конце хотел бы рассказать об интересном применении деревьев. Посмотрим на эту карту расселения человечества по земле.
Если подумать, это не что иное, как дерево, нанесенное на карту земли, где листьями являются разные народы, а стрелки и их ветвление показывают, кто когда с кем разошелся. Эту карту так и сделали. Прочитали ДНК разных народов, построили дерево, приняли во внимание места исконного обитания и учли археологию. А время расставили, во-первых, по данным археологов, а во-вторых, зная, что у человека на поколение происходит около 80 мутаций и, зная число различий между ДНК разных народов, прикинули, сколько времени назад разделились их эволюционные пути.
Как прочитать эволюцию по генам?
Статья на конкурс «био/мол/текст»: Проникновение в тайны эволюции — одно из самых захватывающих направлений в современной биологии. Однако тут есть небольшая проблема: пока не изобретена машина времени, чтобы можно было своими глазами увидеть, как развивалась жизнь на Земле. Впрочем, в наше время существуют методики, которые позволяют приподнять завесу тайны над эволюцией, и одна из основных среди них — построение филогении всего живого, то есть «древа жизни». Для этого можно использовать различные признаки, главный среди которых — это последовательность ДНК, в которой закодировано все разнообразие современных и ископаемых существ. В этой статье рассказывается о методиках построения таких филогений, частично заменяющих ученым машину времени.
Впрочем, в наше время существуют методики, которые позволяют приподнять завесу тайны над эволюцией, и одна из основных среди них — построение филогении всего живого, то есть «древа жизни». Для этого можно использовать различные признаки, главный среди которых — это последовательность ДНК, в которой закодировано все разнообразие современных и ископаемых существ. В этой статье рассказывается о методиках построения таких филогений, частично заменяющих ученым машину времени.
Эта статья представлена на конкурс научно-популярных работ «био/мол/текст»-2013 в номинации «Лучший обзор».
Спонсор конкурса — дальновидная компания Thermo Fisher Scientific. Спонсор приза зрительских симпатий — фирма Helicon.
Что такое филогения и филогенетический анализ?
Филогения всех живых существ, или древо жизни, является нашим представлением о степени родства организмов и о том, как шла эволюция живых существ. Кто является ближайшим родственником человека, и каким был наш общий предок? Вымерли ли динозавры, или их потомки до сих пор живут рядом с нами? Произошли ли теплокровность и способность к полету среди позвоночных единожды? Откуда вообще взялись позвоночные? На все эти вопросы уже есть ответы, и получены они были главным образом с помощью филогенетического анализа.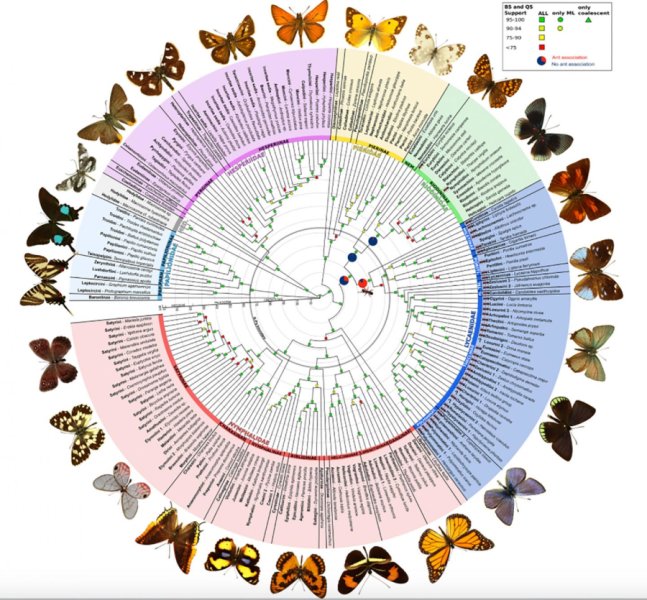
Филогения фактически является той основой, на которую «навешиваются» знания об организмах. Именно она наделяет биологию важным качеством — предсказательностью. Зная те или иные свойства организмов одного вида, с помощью филогении мы можем судить о свойствах родственных ему существ, и даже проследить эволюцию признаков. Древо жизни используется не только в теоретической биологии, но также и в прикладных науках. Например, в медицине и фармакологии филогении используются для того, чтобы понять, откуда были завезены тe или иные вирусы или бактерии, и какие лекарства на них действуют лучше всего [19].
Построение древа жизни является задачей вовсе не тривиальной, и это направление науки, как ни странно, можно считать относительно новым. Разные исследователи пытались проанализировать родственные отношения организмов с самых ранних времен, однако настоящая филогенетическая «революция» случилась только в 50—60-х годах XX века. До 80-х годов деревья строились главным образом на основании морфологических данных, но привлечение ДНК было лишь вопросом времени, поскольку именно в этой молекуле закодированы все признаки организма.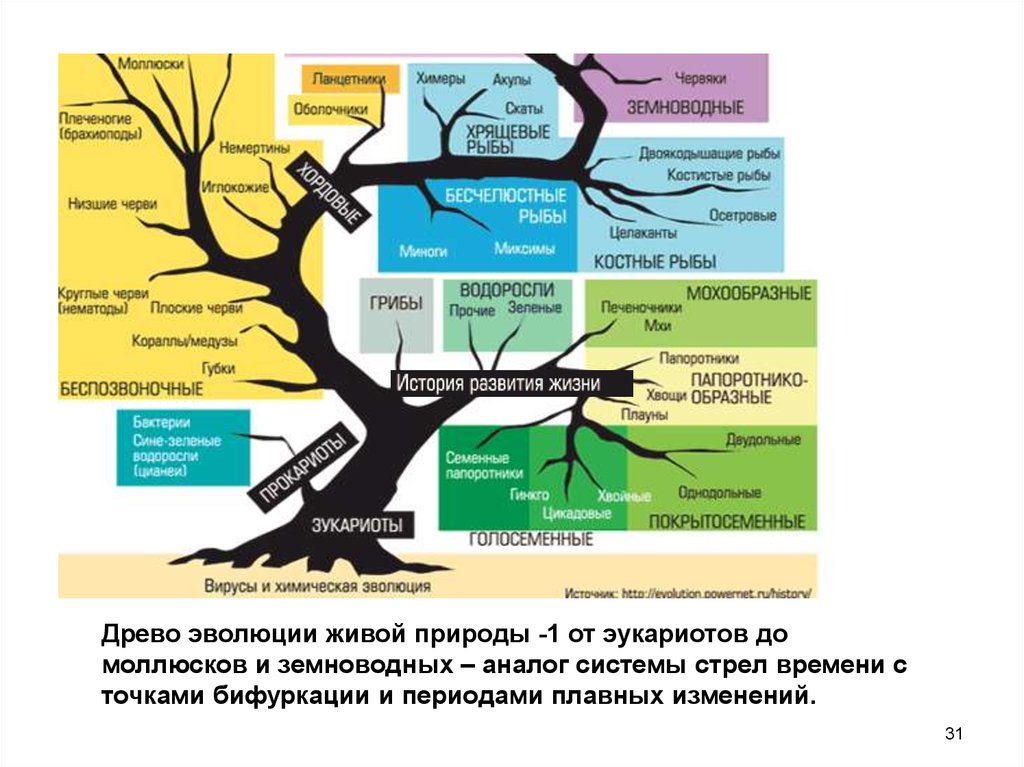
Немного о ДНК
Чтобы понять, как анализируют ДНК, надо вспомнить, как она устроена. ДНК, или дезоксирибонуклиновая кислота, — это очень длинная молекула, которая находится в ядре клетки. ДНК, как правило, состоит из двух закрученных спиралей, а каждая спираль состоит из множества нукеотидов. Нуклеотиды по большей части отличаются друг от друга азотистыми основаниями, которых в ДНК всего четыре: аденин, тимин, гуанин и цитозин. Именно нуклеотиды создают слабые химические связи, которыми соединяются спирали ДНК. Аденин одной спирали связывается с тимином другой спирали, а гуанин связывается с цитозином (рис. 1). Мутация происходит, когда одно основание заменяется на любое другое. Чаще всего замены происходят в парах аденин—гуанин и тимин—цитозин.
Рисунок 1. ДНК. A — аденин, C — цитозин, G — гуанин, T — тимин.
The effects of preservation and conservation treatments on the DNA of museum invertebrate fluid preserved collections
В ДНК есть последовательности нуклеотидов, которые кодируют белки, и есть участки, которые ничего не кодируют. Кодирующие последовательности — это гены. Они могут быть разной длины, но чаще всего имеют определенную структуру, по которой можно сказать — ген это или нет. Именно гены обычно используют для филогенетического анализа.
Кодирующие последовательности — это гены. Они могут быть разной длины, но чаще всего имеют определенную структуру, по которой можно сказать — ген это или нет. Именно гены обычно используют для филогенетического анализа.
Основные принципы построения филогений
Наверное, самое главное правило, которым руководствуются для построения филогений в наше время — это принцип дихотомии: считается, что из трех таксонов, два должны быть более родственны друг другу, чем третий. Поэтому филогении обычно выглядят как дихотомически разветвленные деревья. Если порядок ветвления установлен для всего дерева, то говорят, что оно полностью разрешенное. Иногда в филогениях бывают «кусты» или политомии — это те места, где порядок ветвления неясен, тогда говорят, что дерево не полностью разрешенное. Этот принцип несовершенен, потому что эволюция таксонов далеко не всегда происходит дихотомически. Когда становится понятно, что дихотомия не отражает реальный случай, исследователи привлекают другие схемы — например, филогенетические сети [8].
Методы построения филогений еще в 60-х годах XX века разделились на две основные ветви — фенетические и кладистические. В то время анализ родственных связей основывался на морфологических признаках [12]; с привлечением к построению филогений молекулярных признаков основные принципы анализа родственных связей остались фактически теми же.
- В фенетике построение филогении основано на общем сходстве двух видов — то есть, чем больше общих признаков, тем ближе они друг к другу;
- В кладистике же считается, что только уникальные для какой-либо группы признаки можно использовать для оценки родства таксонов. Родоначальником кладистического анализа является немецкий ученый Вилли Хенниг [6]. Этот автор также ввел и терминологию, которая широко используется до сих пор. Уникальные признаки называются апоморфиями; ветви, которые объединяются апоморфиями — это клады; а сама филогения называется кладограммой (рис. 2) [12].
Чтобы было более понятно, представьте три вида животных: домашнюю мышь, сумчатую мышь и кенгуру. Домашняя мышь и сумчатая мышь очень похожи друг на друга внешне, но у сумчатой мыши и кенгуру есть общая апоморфия — сумка, — что говорит о том, что эти два вида родственные. Но, естественно, филогенетический анализ основывается на гораздо большем количестве признаков, и группы могут иметь несколько апоморфий.
Домашняя мышь и сумчатая мышь очень похожи друг на друга внешне, но у сумчатой мыши и кенгуру есть общая апоморфия — сумка, — что говорит о том, что эти два вида родственные. Но, естественно, филогенетический анализ основывается на гораздо большем количестве признаков, и группы могут иметь несколько апоморфий.
Рисунок 2. Полностью разрешенная кладограмма. Каждое ветвление — это клада. Обозначенные признаки являются апоморфиями.
Principles of biological sciences
Первые шаги. ДНК—ДНК гибридизация
Первые попытки использовать ДНК в качестве основы для построения древа жизни были фенетическими. В 1984 году американские ученые Сибли и Алкист [13] впервые попытались использовать ДНК для прояснения филогении различных видов приматов. Они применили технологию, которая называется «ДНК—ДНК гибридизация». Метод основывается на том, что при копировании в ДНК постоянно происходят мутации. Это приводит к тому, что даже у двух близких родственников последовательности ДНК будут отличаться, не говоря уже о видах. Иными словами, чем дальше находятся организмы на филогенетическом древе, тем больше у них различается ДНК. В данном методе одиночные молекулы ДНК двух видов смешиваются, чтобы они могли образовать «гибридные» двойные спирали, в которых одна половина принадлежит одному виду, а вторая — другому. Затем такие «гибриды» нагреваются, и исследователь смотрит, при какой температуре двойная спираль распадается (или диссоциирует) на две части. Считается, что чем выше температура, требующаяся для распада «гибрида», тем прочнее связь молекул ДНК двух разных видов, и, соответственно, тем ближе эти виды друг к другу (рис. 3).
Иными словами, чем дальше находятся организмы на филогенетическом древе, тем больше у них различается ДНК. В данном методе одиночные молекулы ДНК двух видов смешиваются, чтобы они могли образовать «гибридные» двойные спирали, в которых одна половина принадлежит одному виду, а вторая — другому. Затем такие «гибриды» нагреваются, и исследователь смотрит, при какой температуре двойная спираль распадается (или диссоциирует) на две части. Считается, что чем выше температура, требующаяся для распада «гибрида», тем прочнее связь молекул ДНК двух разных видов, и, соответственно, тем ближе эти виды друг к другу (рис. 3).
Рисунок 3. ДНК—ДНК гибридизация. а — Нагревание ДНК двух видов, в результате которого двойная спираль распадается на две части. б — Охлаждение ДНК, в результате которого молекулы ДНК разных видов гибридизуются друг с другом. в — Нагревание ДНК, в результате которого гибридные молекулы ДНК распадаются.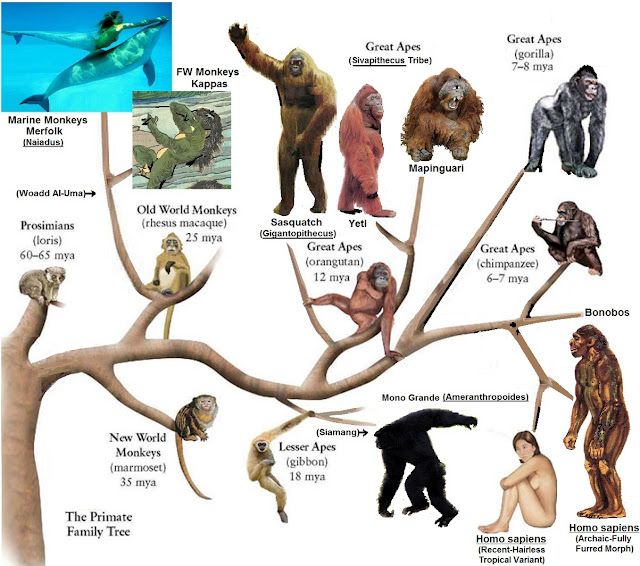
ThinkQuest, рисунок с изменениями
Очень быстро стало понятно, что такой метод не может быть очень точным. Дело в том, что гены могут гибридизоваться не только с гомологичными им генами (гены-ортологи), но и с копиями этих генов, которых в геноме может быть довольно много (гены-паралоги) [15]. Постепенно, с развитием методики секвенирования генов , главным источником для построения филогений стали последовательности ДНК или белков, записанные в виде компьютерных файлов. В последние годы скорость накопления генетической информации растет все увеличивающимися темпами, что окончательно утверждает филогению как метод анализа и обработки биологических текстов.
О стремительном развитии технологий секвенирования ДНК и их роли в науке и обществе можно прочесть в статьях «454-секвенирование (высокопроизводительное пиросеквенирование ДНК)» [20], «Важнейшие методы молекулярной биологии и генной инженерии» [21], «Код жизни: прочесть не значит понять» [22], «Перевалило за тысячу: третья фаза геномики человека» [23] и «Огурцы-убийцы, или как встретились Джим Уотсон и Гордон Мур» [24].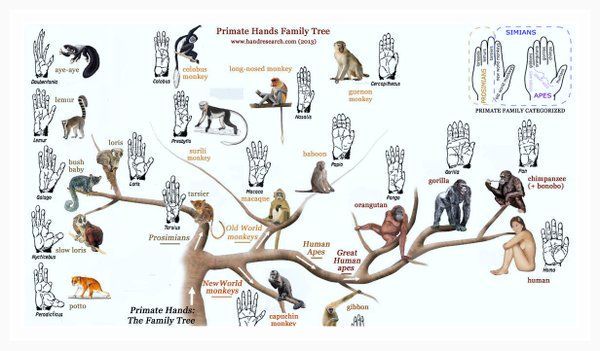 — Ред.
— Ред.
Метод матрицы расстояний (distance matrix)
Метод матрицы расстояний, по сути, является фенетическим. Его основа — расчет попарных различий между соответствующими генами всех видов, участвующих в таком анализе. Делается это следующим образом: гены каждого анализируемого вида сравниваются по каждой позиции нуклеотидов, и чем больше найдено отличий, тем больше будет «расстояние» между видами. Затем строится матрица, в которую заносится это значение для каждой возможной пары сравниваемых генов. Далее матрица расстояний является входной информацией для алгоритмов построения деревьев.
Самый популярный среди подобных алгоритмов — это метод ближайших соседей (neighbour joining). Среди анализируемых видов находят два с минимальными различиями в последовательности (т.е., максимально похожие). Исходя из составленной матрицы, данные об этих видах «объединяются», и далее они участвуют в анализе в объединенном состоянии. Виды один за другим проходят эту процедуру до тех пор, пока не будет найдено одно, полностью разрешенное дерево. Этот алгоритм хорош тем, что он относительно прост и подходит для обработки больших наборов данных (рис. 4) [3].
Этот алгоритм хорош тем, что он относительно прост и подходит для обработки больших наборов данных (рис. 4) [3].
Рисунок 4. Метод ближайшего соседа
Wikipedia
Разные авторы, однако, перечисляют некоторые минусы метода ближайших соседей. Например, есть мнение, что этот метод хуже работает с таксонами, которые филогенетически далеки друг от друга [4], [17]. Также недостатком можно считать и то, что метод всегда выдает дерево с одним-единственным возможным вариантом ветвления [3]. Это происходит потому, что алгоритм подразумевает построение одной филогении без сравнения с другими, тогда как в кладистических методах оцениваются деревья с различным порядком ветвления. Несмотря на то, что в серьезных филогенетических анализах методы матрицы расстояний сейчас почти не используются, они применяются, например, для быстрого построения филогений близкородственных бактерий и вирусов [18].
Этот подход получил большую популярность при анализе морфологических данных, а также какое-то время применялся и для молекулярных исследований. Первый этап анализа — это создание матрицы признаков. Каждый признак должен иметь хотя бы два состояния. Состояний может быть больше, в морфологии они могут описывать разные формы и структуры. Если на кладограмме у какого-то таксона или группы таксонов состояние отличается от предкового, то это называется «переходом из одного состояния в другое». Суть этого алгоритма в том, чтобы найти такое дерево, где присутствует наименьшее суммарное число переходов из одного состояния в другое для всех признаков. В этом случае кладограмма и отображаемая на ней эволюция будут считаться наиболее экономными, а, значит, и более вероятными [3], [12], [16], [17].
Первый этап анализа — это создание матрицы признаков. Каждый признак должен иметь хотя бы два состояния. Состояний может быть больше, в морфологии они могут описывать разные формы и структуры. Если на кладограмме у какого-то таксона или группы таксонов состояние отличается от предкового, то это называется «переходом из одного состояния в другое». Суть этого алгоритма в том, чтобы найти такое дерево, где присутствует наименьшее суммарное число переходов из одного состояния в другое для всех признаков. В этом случае кладограмма и отображаемая на ней эволюция будут считаться наиболее экономными, а, значит, и более вероятными [3], [12], [16], [17].
Тут возникает вопрос: почему мы вообще считаем, что эволюция должна быть экономной? Дело в том, что это соответствует главному методологическому принципу науки, который заключается в том, что из нескольких равновероятных объяснений надо выбирать наиболее простое, с привлечением как можно меньшего количества сущностей. Этот метод еще называется «Бритвой Оккама». В одной из книг по филогении [3] есть шутливый пример. Представьте, что в одном и том же городе где-то в Северной Америке в соответствующую службу поступает два звонка о том, что по улицам гуляет тигр. Понято, что легче всего предположить, что это один и тот же тигр, который сбежал из зоопарка. Гипотеза, что в городе, где тигров в природе никогда не было, откуда-то появилось сразу же два таких хищника, гораздо менее вероятна.
В одной из книг по филогении [3] есть шутливый пример. Представьте, что в одном и том же городе где-то в Северной Америке в соответствующую службу поступает два звонка о том, что по улицам гуляет тигр. Понято, что легче всего предположить, что это один и тот же тигр, который сбежал из зоопарка. Гипотеза, что в городе, где тигров в природе никогда не было, откуда-то появилось сразу же два таких хищника, гораздо менее вероятна.
Эволюция признака — тоже событие нечастое, и когда мы видим два похожих по строению органа, то мы предполагаем, что орган произошел один раз [3]. Это не означает, что признак действительно произошел только один раз, просто это наиболее вероятно. Кладограмма строится на основании многих признаков, и чем больше апоморфий характеризует ту или иную ветвь, тем больше доверия она вызывает.
Плюс метода наибольшей экономии в том, что он интуитивно понятен и довольно прост, но в молекулярных анализах он очень быстро потерял популярность. Один из его недостатков в том, что он не учитывает длину ветвей, которая отображает количество замен нуклеотидов во время эволюции той или иной клады [3]. Некоторые ветви на дереве будут длиннее, потому что скорость эволюции там была выше. При использовании метода наибольшей экономии длинные ветви будут «притягиваться» друг к другу. Этот феномен возникает потому, что чем больше замен нуклеотидов в двух ветвях, тем выше шанс на то, что некоторые из них случайно совпадут, и будут расцениваться как общие апоморфии, даже если это абсолютно не соответствует реальному положению дел.
Некоторые ветви на дереве будут длиннее, потому что скорость эволюции там была выше. При использовании метода наибольшей экономии длинные ветви будут «притягиваться» друг к другу. Этот феномен возникает потому, что чем больше замен нуклеотидов в двух ветвях, тем выше шанс на то, что некоторые из них случайно совпадут, и будут расцениваться как общие апоморфии, даже если это абсолютно не соответствует реальному положению дел.
Другой минус в том, что метод не учитывает разные модели замены нуклеотидов [17]. Например, в методе наибольшей экономии аденин имеет одинаковую вероятность уступить место как тимину, так и цитозину, хотя, как уже отмечалось выше, в организме аденин скорее заменится на цитозин, чем на тимин.
Методы, основанные на моделях эволюции
Наиболее часто используемые методы построения филогений на основе молекулярных данных основываются на моделях эволюции. Один из первых стал метод максимального правдоподобия (maximum likelihood). Для расчета кладограммы, помимо последовательности ДНК, надо выбрать модель замены нуклеотидов, на основании которой будут рассчитываться вероятности. Также в расчет берется длина ветви или эволюционная дистанция между двумя таксонами. Во время анализа рассчитывается, какая длина ветви наиболее вероятна с точки зрения выбранной модели, вероятности всех ветвей кладограммы умножаются, и кладограмма, имеющая наибольшую вероятность, считается правильной [3], [16], [17].
Также в расчет берется длина ветви или эволюционная дистанция между двумя таксонами. Во время анализа рассчитывается, какая длина ветви наиболее вероятна с точки зрения выбранной модели, вероятности всех ветвей кладограммы умножаются, и кладограмма, имеющая наибольшую вероятность, считается правильной [3], [16], [17].
Последний и, наверное, самый популярный в наше время метод — это Байесовский вывод (Bayesian inference). Он, в общем, похож на метод максимального правдоподобия, поскольку также основывается на модели и длине ветвей. Но отличие Байесовского вывода в том, что тут берется в расчет еще один фактор — апостериорная вероятность (posterior probablity), которая рассчитывается на основании как исходных данных, так и полученных результатов анализа [3], [16], [17]. Это не очень понятно интуитивно, но суть в том, что в ходе анализа исследователь получает новые данные, которые тоже можно применить.
Приведу очень простой пример. Пусть у нас есть мешок с сотней шариков, половина их которых красные и половина — белые.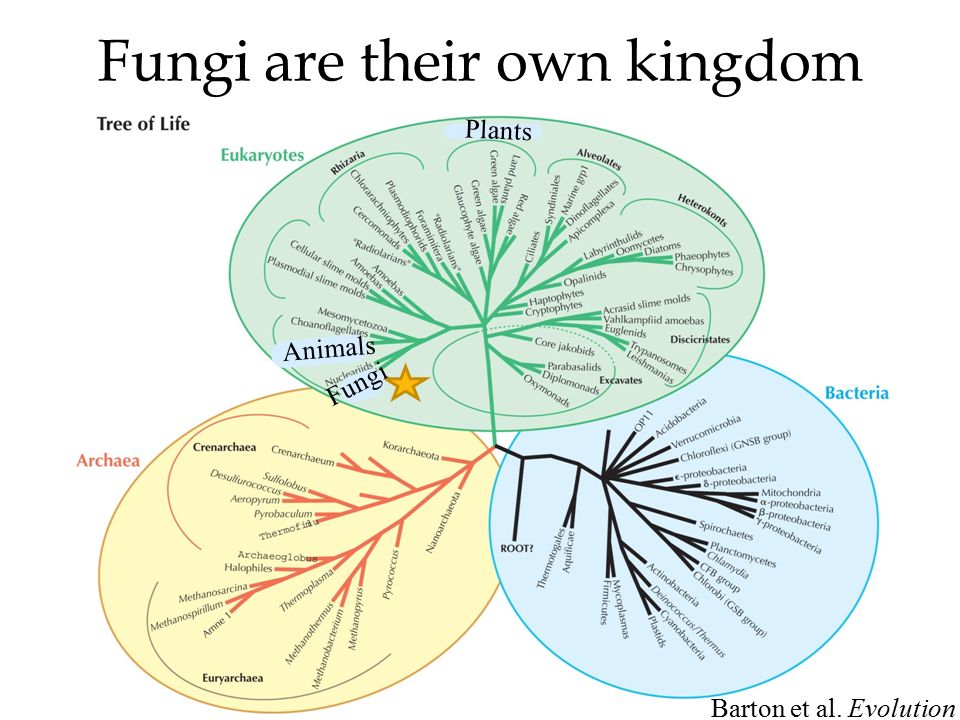 Изначально вероятность вытащить шарик как белого, так и красного цвета равна 50%. Но, допустим, мы вытащили 20 красных и 40 белых шариков, и в мешке остались 30 красных и 10 белых шариков. Это означает, что к текущему моменту шанс вытащить красный шарик равен 75%, а белый — 25%, что кардинальным образом отличается от исходного состояния. В Байесовском выводе используются похожая логика, хотя, конечно же, расчеты там гораздо сложнее.
Изначально вероятность вытащить шарик как белого, так и красного цвета равна 50%. Но, допустим, мы вытащили 20 красных и 40 белых шариков, и в мешке остались 30 красных и 10 белых шариков. Это означает, что к текущему моменту шанс вытащить красный шарик равен 75%, а белый — 25%, что кардинальным образом отличается от исходного состояния. В Байесовском выводе используются похожая логика, хотя, конечно же, расчеты там гораздо сложнее.
Несмотря на все видимые плюсы двух последних методов, тут тоже можно найти некоторые сложности. Главная их слабость в том, что каждый исследователь вынужден подбирать модели самостоятельно, и совсем не обязательно, что он сделает выбор правильно. Но у этой проблемы есть решение. Во-первых, есть программы, которые могут помочь подобрать модель; во-вторых, уже есть алгоритмы на основе Байесовского метода, которые могут «прыгать» с модели на модель, тем самым тестируя их. Еще одна проблема, скорее всего, решаемая с развитием техники, заключается в том, что обсчеты филогений с использованием последних двух методов довольно сложные и требуют много времени и хороших компьютеров.
Все же насколько достоверны филогении?
Думаю, что внимательный читатель заметил, что многие перечисленные методы основаны на вероятностях, и у него может возникнуть закономерный вопрос: как можно доверять филогении, если всегда есть шанс, что построенное дерево ошибочно и не соответствует действительному ходу эволюции? Действительно, методы несовершенны, но на этот вопрос ответ есть.
Во-первых, в филогенетических методах есть понятие «поддержка: чем больше уникальных признаков поддерживают дерево или какую-то его ветвь, тем больше доверия они вызывают [12]. Само дерево может иметь низкую поддержку, зато свидетельств в пользу отдельных его ветвей может быть так много, что корректность не вызовет сомнений. Для подтверждения результата исследователи могут использовать совокупности признаков: последовательности ДНК, РНК и белков, морфологические данные, особенности поведения организмов и многое другое [11]. Когда независимые признаки подтверждают друг друга, уверенность в результате гораздо выше.
Второй ответ на поставленный вопрос еще более обнадеживающий. Его дают эксперименты, проведенные на разных организмах, для которых известна генеалогия, то есть настоящая эволюционная история [1], [5], [7], [10]. Можно привести в пример опыт с мышами, когда филогенетический анализ провели на основе ДНК 24-х линий этих животных. Оказалось, что наблюдаемая последовательность поколений и полученная филогения почти полностью соответствуют друг другу [1]. Это значит, что используемые методы как минимум способны правильно отображать эволюцию.
Плюсы и минусы молекулярных методов построения филогений
У молекулярных методов есть много преимуществ перед морфологическим анализом. Во-первых, ДНК содержит в себе множество данных, которые можно использовать в расчетах, — ведь в генах могут содержаться сотни нуклеотидов. Чаще всего для оценки родства используют больше одного гена, тогда как для анализа на основе морфологических данных используют несколько десятков признаков.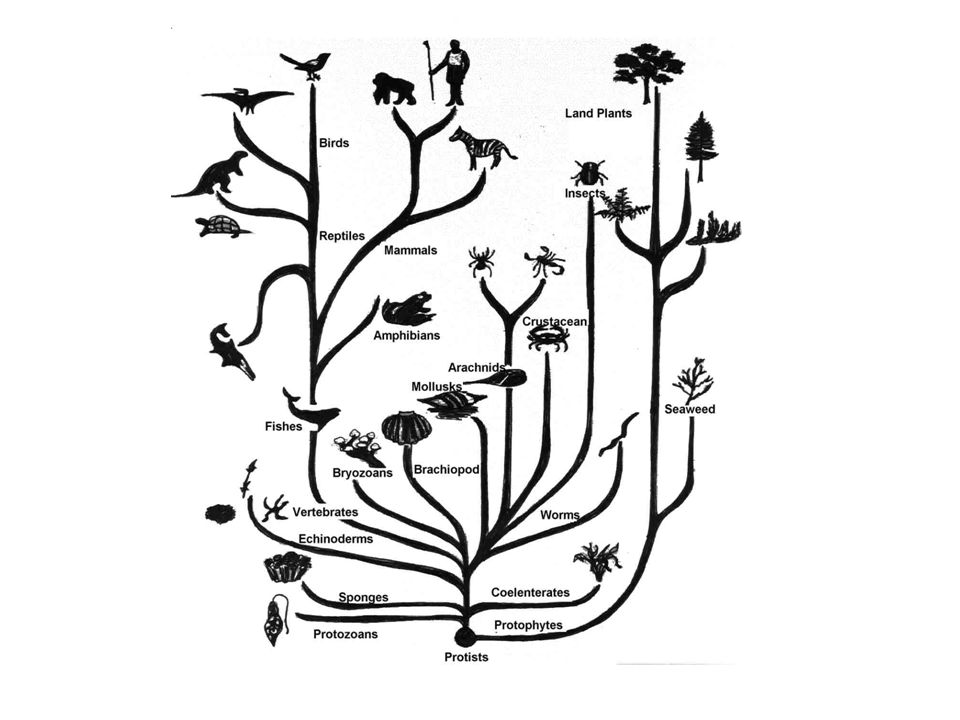 Во-вторых, анализ ДНК считается более объективным. Дело в том, что морфологические признаки разные люди могут трактовать и кодировать по-разному, тогда как нуклеотиды всегда одинаковы . В-третьих, ДНК можно использовать как для анализа групп высоких рангов, так и для выяснения отношений между видами, и даже между отдельными индивидами. Морфологический же анализ более достоверен при работе с таксонами высоких рангов, чем на уровне видов, — просто потому, что чем выше ранг, тем лучше отличаются группы, и тем легче отличить аналогичный признак от гомологичного.
Во-вторых, анализ ДНК считается более объективным. Дело в том, что морфологические признаки разные люди могут трактовать и кодировать по-разному, тогда как нуклеотиды всегда одинаковы . В-третьих, ДНК можно использовать как для анализа групп высоких рангов, так и для выяснения отношений между видами, и даже между отдельными индивидами. Морфологический же анализ более достоверен при работе с таксонами высоких рангов, чем на уровне видов, — просто потому, что чем выше ранг, тем лучше отличаются группы, и тем легче отличить аналогичный признак от гомологичного.
В частности, анализ консервативных последовательностей рибосомальных РНК микроорганизмов позволил установить, что все живое на Земле делится не на два царства, как считали несколько десятилетий назад, — эукариот и бактерий, — а на три: эукариот, бактерий и архей. Морфологическое сходство бактерий и архей с лихвой окупается огромной разницей их молекулярного устройства. Честь этого открытия принадлежит Карлу Вёзе. См. также статьи: «Карл Вёзе (1928—2012)», «Эволюция между молотом и наковальней, или как микробиология спасла эволюцию от поглощения молекулярной биологией». — Ред.
См. также статьи: «Карл Вёзе (1928—2012)», «Эволюция между молотом и наковальней, или как микробиология спасла эволюцию от поглощения молекулярной биологией». — Ред.
Несмотря на то, что преимущество молекулярного анализа кажется вполне обоснованным, есть все же и несколько причин, по которым морфологию нельзя отправить «в отставку».
Первая причина заключается в том, что не каждый организм подходит для выделения ДНК. Он должен быть собран и сохранен специальным образом, иначе эта молекула просто разрушается. Множество редких и интересных видов было описано много десятков лет назад, когда еще даже про ДНК ничего не знали, и в наши дни не очень понятно, где их искать и как собирать. В первую очередь это касается мелких членистоногих, — особенно насекомых, которых чаще всего хранят сухими. То же самое можно сказать и о палеонтологических находках вымерших видов. Для оценки родства таких групп можно использовать только морфологические методы.
Вторая причина заключается в том, что далеко не всегда результаты молекулярных филогенетических методов вызывают доверие. Иногда бывает так, что они не совпадают с устоявшимися «классическими» взглядами. Это, конечно, не означает, что именно молекулярные данные неверны, просто такие несовпадения являются «звоночком», что где-то закралась ошибка. Несовпадения могут быть не только из-за ошибок в самом анализе, но и из-за того, что были неправильно выбраны гены. Гены, мутирующие с высокой скоростью, подходят для выяснения родства между видами, но не походят для анализа групп более высоких рангов. Но гомологичные гены в разных группах организмов могут меняться с разной скоростью, поэтому гены, подходящие для анализа одной группы, могут не подходить для другой группы того же ранга. В общем, подбор нужных участков ДНК может оказаться не очень легкой работой, особенно если учесть, что далеко не все гены у всех видов хорошо изучены.
Иногда бывает так, что они не совпадают с устоявшимися «классическими» взглядами. Это, конечно, не означает, что именно молекулярные данные неверны, просто такие несовпадения являются «звоночком», что где-то закралась ошибка. Несовпадения могут быть не только из-за ошибок в самом анализе, но и из-за того, что были неправильно выбраны гены. Гены, мутирующие с высокой скоростью, подходят для выяснения родства между видами, но не походят для анализа групп более высоких рангов. Но гомологичные гены в разных группах организмов могут меняться с разной скоростью, поэтому гены, подходящие для анализа одной группы, могут не подходить для другой группы того же ранга. В общем, подбор нужных участков ДНК может оказаться не очень легкой работой, особенно если учесть, что далеко не все гены у всех видов хорошо изучены.
Третья причина — это высокая стоимость секвенирования генов. Для построения филогении одного небольшого рода можно легко потратить пару тысяч долларов. А если учесть, что гены не всегда подбирают правильно с первого раза, или некоторые экземпляры оказываются непригодными для секвенирования, то анализ надо проводить повторно, и цена может быть больше, чем предполагалось изначально. Анализ же на основе морфологических признаков обходится гораздо дешевле.
Анализ же на основе морфологических признаков обходится гораздо дешевле.
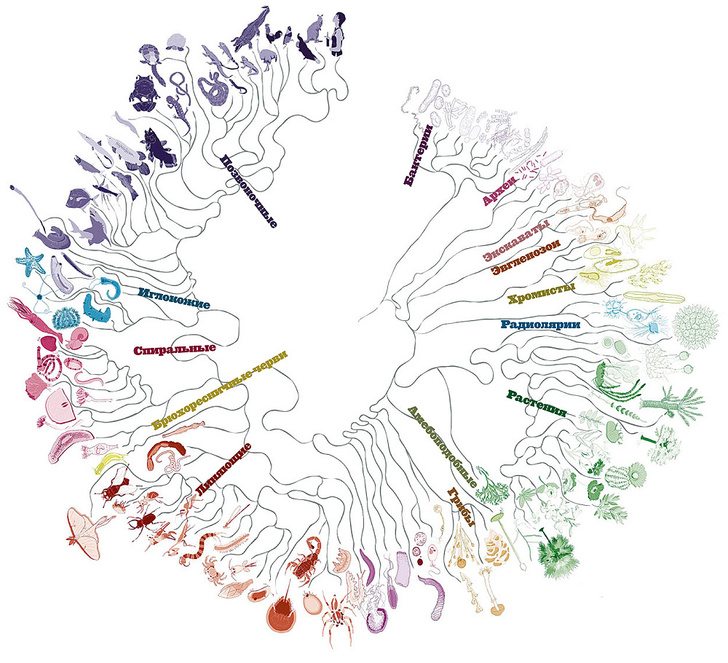
Анализ ДНК, безусловно, стал довольно популярным и быстроразвивающимся подходом построения филогений в наши дни. Сейчас специалисты уже используют не просто отдельные гены: в последние годы появились филогенетические исследования на основе более десятка генов или целых митохондриальных геномов [14]. Запущены проекты секвенирования целых геномов разных видов [9], а также проекты для объединения всего живого мира в единое «древо жизни» (рис. 5) [2]. В качестве частного примера можно привести исследование, в результате которого была уточнена «родословная» членистоногих [25]. Наверное, наука сейчас переживает один из самых интересных периодов в развитии анализа ДНК, когда уже видно, что это направление масштабно и многообещающе, и что есть еще очень многое, что нам предстоит узнать о геномах разных организмов. Однако насколько молекулярные методы в филогениях можно развивать, и где граница их применения — покажет будущее.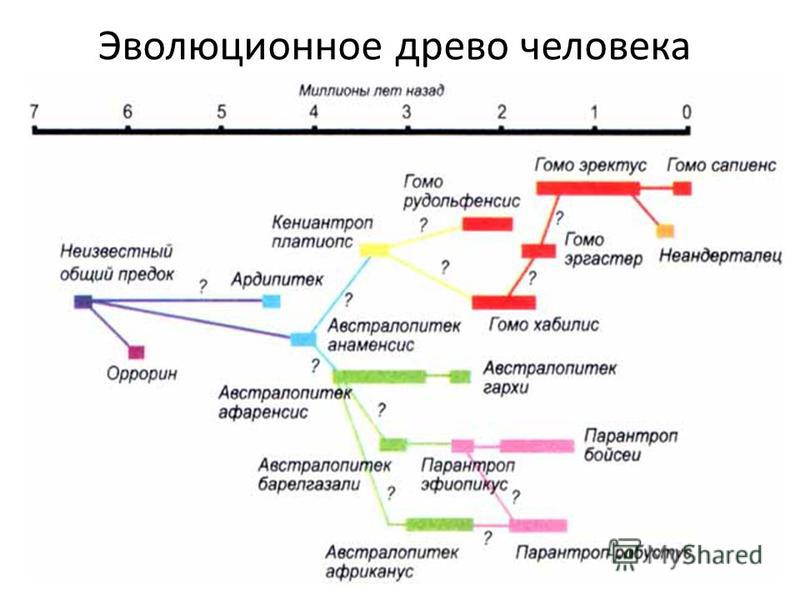
Рисунок 5. Филогения всех живых существ, или «древо жизни»
Ingmur, рисунок с изменениями
- W. Atchley, W. Fitch. (1991). Gene trees and the origins of inbred strains of mice. Science. 254, 554-558;
- Assembling Tree of Life (AToL). (2007). The National Science Foundation;
- Baum D.A. and Smith S.D. Tree thinking: an introduction to phylogenetic biology. Greenwood Village, CO: Roberts and Company Publishers, Inc., 2012;
- William J. Bruno, Nicholas D. Socci, Aaron L. Halpern. (2000). Weighted Neighbor Joining: A Likelihood-Based Approach to Distance-Based Phylogeny Reconstruction. Molecular Biology and Evolution. 17, 189-197;
- Douglas T. (1999-2001). 29+ Evidences for Microevolution. TalkOrigins Archive;
- Hennig W. Grundzüge einer Theorie der phylogenetischen Systematik.
 Berlin: Deutscher Zentralverlag, 1950;
Berlin: Deutscher Zentralverlag, 1950; - D. Hillis, J. Bull, M. White, M. Badgett, I. Molineux. (1992). Experimental phylogenetics: generation of a known phylogeny. Science. 255, 589-592;
- V. Kunin. (2005). The net of life: Reconstructing the microbial phylogenetic network. Genome Research. 15, 954-959;
- Jenna Morgan Lang, Aaron E. Darling, Jonathan A. Eisen. (2013). Phylogeny of Bacterial and Archaeal Genomes Using Conserved Genes: Supertrees and Supermatrices. PLoS ONE. 8, e62510;
- Michael M. Miyamoto, Walter M. Fitch. (1995). Testing Species Phylogenies and Phylogenetic Methods with Congruence. Systematic Biology. 44, 64;
- Maureen A. O’Malley, Orkun S. Soyer. (2012). The roles of integration in molecular systems biology. Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences.
 43, 58-68;
43, 58-68; - Schuh R.T. and Brower A.V.Z. (2003). Biological systematics: principles and applications (2nd Edition). Ithaca: Cornell University Press, 2009;
- Charles G. Sibley, Jon E. Ahlquist. (1984). The phylogeny of the hominoid primates, as indicated by DNA-DNA hybridization. J Mol Evol. 20, 2-15;
- Nan Song, Ai-Ping Liang, Cui-Ping Bu. (2012). A Molecular Phylogeny of Hemiptera Inferred from Mitochondrial Genome Sequences. PLoS ONE. 7, e48778;
- Mark Springer, Carey Krajewski. (1989). DNA Hybridization in Animal Taxonomy: A Critique from First Principles. The Quarterly Review of Biology. 64, 291-318;
- Wiley E.O. and Lieberman B.S. Phylogenetics: the theory of phylogenetic systematics (2nd Edition). Hoboken, NJ: Wiley-Blackwell, 2011;
- Ziheng Yang, Bruce Rannala.
 (2012). Molecular phylogenetics: principles and practice. Nat Rev Genet. 13, 303-314;
(2012). Molecular phylogenetics: principles and practice. Nat Rev Genet. 13, 303-314; - Longyu Zheng, Tawni L. Crippen, Leslie Holmes, Baneshwar Singh, Meaghan L. Pimsler, et. al.. (2013). Bacteria Mediate Oviposition by the Black Soldier Fly, Hermetia illucens (L.), (Diptera: Stratiomyidae). Sci Rep. 3;
- Мамонты, кости и лекарственная устойчивость: новые технологии позволяют изучать эволюцию возбудителей инфекционных заболеваний;
- 454-секвенирование (высокопроизводительное пиросеквенирование ДНК);
- Важнейшие методы молекулярной биологии и генной инженерии;
- Код жизни: прочесть не значит понять;
- Перевалило за тысячу: третья фаза геномики человека;
- Огурцы-убийцы, или Как встретились Джим Уотсон и Гордон Мур;
- Уточнение «родословной» членистоногих.
OneZoom Древо Жизни Исследователь
Интерактивная карта эволюционных связей между всеми известными науке живыми существами. Откройте для себя своих фаворитов, посмотрите, какие виды находятся под угрозой, и поразитесь разнообразию жизни на Земле.
Наш проводник по древу жизни разработан так, чтобы быть легко доступным для всех. Мы также предоставляем образовательные инструменты для учителей, программное обеспечение для ученых и интерактивные экспонаты для музеев. Это бесплатный ресурс сообщества без рекламы.
Что такое дерево жизни?
Древо жизни показывает, как связана вся жизнь на земле.
Каждый лист представляет отдельный вид.
Ветви показывают, как эти многочисленные виды произошли от общих предков на протяжении миллиардов лет.
В нашем интерактивном древе жизни вы можете исследовать отношения между 2 235 076 видов и чудес на 105 381 изображений на одной масштабируемой странице.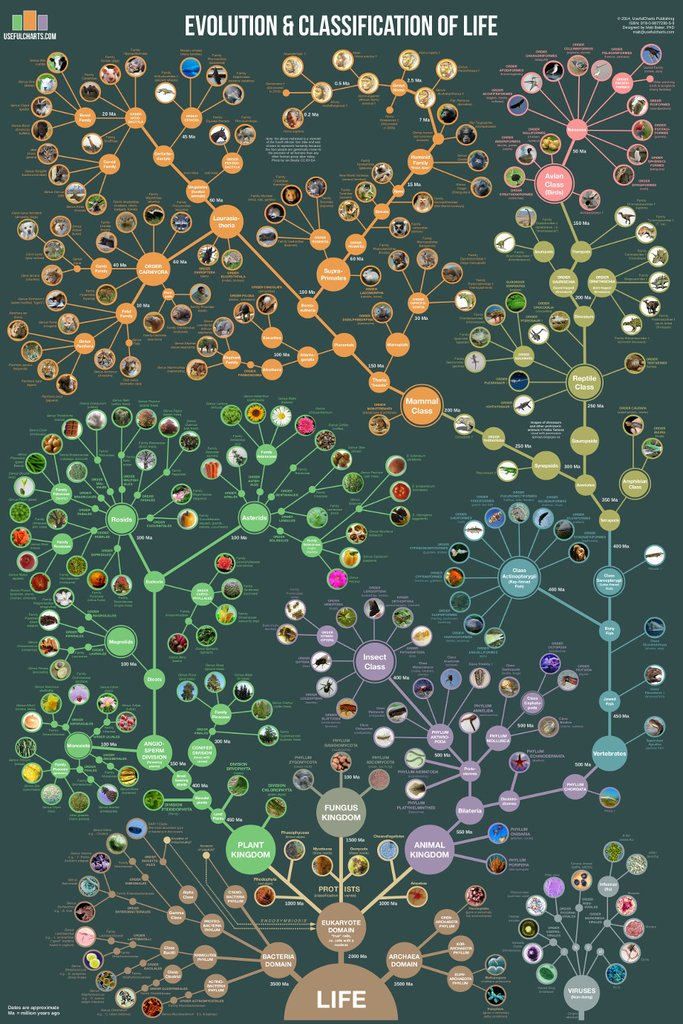
Популярные места для начала изучения…
- Человек
- Плотоядные
- Лягушки и жабы
- Орхидеи
- Совы, сипухи
- Бабочки и мотыльки
- Серый волк (и домашняя собака)
- Водяные лилии
- Осьминоги
- Попугаи
- Твердые кораллы
- Семья Роуз
- Слоны
- Грибообразующие грибы
- Лучепёрые рыбы
- Стрекозы и стрекозы
- Дуб
- Колибри
- Жуки
- хамелеон
Начните с истоков жизни
Биоразнообразие и сохранение.
 ..
.. Мы хотим помочь каждому оценить биоразнообразие и необходимость его сохранения.
Красные листья на нашем древе жизни показывают виды, которые, как известно, находятся под угрозой исчезновения.
Зеленые листья сейчас в порядке, но могут оказаться под угрозой исчезновения.
Большинство листьев серые, а это означает, что неизвестно, насколько им угрожает опасность.
Для всех
- Ваше имя на дереве
- Отправить tr-eCard
Для образования
- Программа запуска дисплея
- Учебные материалы
- Инструменты разработчика
- Индекс популярности
Великолепная программа, блестяще интуитивно понятная визуализация древа жизни
Лучший инструмент для изучения древа жизни на этой планете, который я когда-либо видел.
Одна из самых крутых вещей, что я видел в интернете
Проект визуализации Genius, отображающий все древо жизни в виде единой исследуемой диаграммы.
Это произведет революцию в том, как мы преподаем и понимаем Древо Жизни. Это бесценный инструмент для передачи грандиозного масштаба истории жизни.
OneZoom — один из самых захватывающих проектов визуализации, которые я видел за последние годы!
Всю неделю я рассказывал людям, какой невероятный OneZoom!
Замечательная идея и ресурс!
Лучшее интерактивное дерево жизни!
На мой взгляд, это лучшая визуализация эволюции для неспециалиста.
- Поддержите листовку прямо сейчас!
29 сентября 2022 г.
Доноры теперь могут возобновить свою поддержку спонсируемых отпусков. Спонсорство предоставляется только на четырехлетний период, однако те, кто не продлевает подписку, пока останутся на нашем дереве жизни. Доноры получат напоминание по электронной почте до истечения срока их спонсируемых отпусков.
Подробнее…
6 февраля 2022 г.
Официально выпущена версия OneZoom 3.
 6 (псевдоним: music_furry_lobster). Теперь мы показываем карты встречаемости (распространения) видов во всплывающих вкладках при нажатии на вид, и у нас есть инструкции о том, как запустить специальную версию OneZoom с вашими собственными изображениями вместо наших собственных изображений или вместе с ними. Есть также несколько незначительных изменений в дереве, с более поздним расположением первых нескольких (архейных) ветвей и добавлением дронта!
6 (псевдоним: music_furry_lobster). Теперь мы показываем карты встречаемости (распространения) видов во всплывающих вкладках при нажатии на вид, и у нас есть инструкции о том, как запустить специальную версию OneZoom с вашими собственными изображениями вместо наших собственных изображений или вместе с ними. Есть также несколько незначительных изменений в дереве, с более поздним расположением первых нескольких (архейных) ветвей и добавлением дронта!Подробнее…
14 декабря 2021 г.
Объявление о публикации «Динамическая визуализация деревьев с миллионом верхушек: проект OneZoom», опубликованной сегодня в журнале Methods in Ecology and Evolution. Эта публикация сопровождает OneZoom версии 3.5 «Шоколадная морская звезда» и объясняет многие научные аспекты проекта OneZoom.
Подробнее…
15 октября 2021 г.
Выпущена версия OneZoom 3.5 «Шоколадная морская звезда» с новыми и улучшенными цветовыми схемами и стилями, а также с существенными улучшениями содержимого и интерпретаций страниц на всем сайте.

Подробнее…
16 октября 2020 г.
Выпущена версия OneZoom 3.4.1 с новыми общедоступными API, предоставляющими информацию о популярности видов и репрезентативных изображений для групп видов, а также другие инструменты, предназначенные для использования исследователями и другими научными коммуникационными проектами.
Подробнее…
- Поддержите листовку прямо сейчас!
- Хронология
- Подтверждения
- О OneZoom
- Команда
- Часто задаваемые вопросы
- Начать изучение
- Музейные экспонаты
- Работайте с нами
- Индекс популярности
- Поддержите нашу работу
- Ян Вонг и Джеймс Розинделл (2021) Динамическая визуализация деревьев с миллионом кончиков : проект OneZoom. Методы экологии и эволюции 00 : 1–11. дои: 10.1111/2041-210X.13766
- Джеймс Розинделл и Ян Вонг (2018) Биоразнообразие, древо жизни и научная коммуникация в Филогенетическом разнообразии под ред. Роза А. Шерсон и Дэниел П. Фейт
- Джеймс Розинделл и Люк Хармон (2012) OneZoom: исследователь фракталов Древа жизни. Биология PLoS 10 : e1001406. doi: 10.1371/journal.pbio.1001406
- Исследования эволюции человека
- Климат и эволюция человека
- Влияние климата на эволюцию человека
- Выживание адаптивного
- Интерактивная временная шкала эволюции человека
- Восточноафриканские исследовательские проекты
- Полевой блог Olorgesailie
- 2011 Рассылки Олоргесайли
- 2004 Олоргесайские депеши
- 1999 Олоргесайские депеши
- Проект бурения в Олоргесайли
- Канам, Кения
- Канджера, Кения
- Ол Педжета, Кения
- Олоргесайли, Кения
- Эволюция человеческих инноваций
- Приключения в рифтовой долине: интерактив
- Полевой блог Olorgesailie
- Азиатские исследовательские проекты
- «Хоббиты» на Флоресе, Индонезия
- Древнейшие люди в Китае
- Бозе, Китай
- Антропоцен: эпоха людей
- Экспертиза окаменелостей: интерактивная
- Что нового в Human Origins?
- Цифровой архив зубов копытных и хищников
- инструкции
- Хищник
- Зубы копытных
- Климат и эволюция человека
- Доказательства эволюции человека
- Поведение
- Поведение приматов
- Следы
- Следы из Кооби Фора, Кения
- Следы Лаэтоли
- Следы Энгаре Серо, Танзания
- Каменные инструменты
- Инструменты раннего каменного века
- Hammerstone из Majuangou, Китай
- Ручной топор и тектиты от Bose, Китай
- Ручной топор из Европы
- Ручной топор из Индии
- Oldowan Tools из Локалалей, Кения
- Олдувайский чоппер
- Каменные инструменты из Majuangou, Китай
- Инструменты среднего каменного века
- Инструменты позднего каменного века
- Бурин от Laugerie Haute & Basse, Дордонь, Франция
- Ла-Мадлен, Дордонь, Франция
- Инструменты раннего каменного века
- Получение еды
- Костяные инструменты
- Расчлененные кости животных из Гоны, Эфиопия
- Катанда Костяной наконечник гарпуна
- Старейшее деревянное копье
- Проколотая лопатка лошади
- Каменные лезвия серпа
- Точка снаряда
- Переноска и хранение
- Самая старая керамика
- Фрагмент керамики
- Очаги и укрытия
- Орудия из измененного огнем камня
- Приют Терра Амата
- Погребение
- Кафзе: старейшее преднамеренное захоронение
- Информация о записи
- Ассирийская цилиндрическая печать
- Налет охры Бломбос
- Кость Ишанго
- Изготовление одежды
- Костяные шила
- Иглы из кости и слоновой кости
- Искусство и музыка
- Статуэтки
- Резной бегущий лев из слоновой кости
- Женский торс цвета слоновой кости
- Фигурка лошади из слоновой кости
- Скульптура лошади из слоновой кости
- Леди Брассемпуи
- Фигурка человека-льва
- Виллендорф Венера
- Ювелирные изделия
- Бусины из древних ракушек
- Резной костяной диск
- Ожерелье из кроманьонской раковины
- Старейшие известные бусины из ракушек
- Музыкальные инструменты
- Древняя флейта
- Наскального искусства
- Древние пигменты
- Мемориальная доска Аполлона-11
- Другие декорированные предметы
- Резной жезл из рога с лошадьми
- Геометрический прямоугольный разрез кости
- Тата Мемориальная доска
- Статуэтки
- Окаменелости человека
- Разновидность
- Окаменелости
- Таинственный Череп Интерактивный
- Шанидар 3 — Скелет неандертальца
- 3D коллекция
- Артефакты
- Фигурка бизона
- Резная бусина в виде лошадиной головы
- Резной мамонт из слоновой кости
- Клинописная глиняная табличка
- Обожженный глиняный бизон
- Венера из обожженной глины из Долни Вестонице
- Леспуг Венера
- Окаменелости
- Приматы
- Артефакты
- Генетика
- Один вид, живущий по всему миру
- Изменение цвета кожи человека
- Древняя ДНК и неандертальцы
- Знакомства
- Интерактивная временная шкала эволюции человека
- Генеалогическое древо человека
- Снимки во времени
- Сварткранс, Южная Африка
- Олоргесайли, Кения
- Шанидар, Ирак
- Поведение
- Характеристики человека
- Ходьба в вертикальном положении
- Инструменты и еда
- Тела
- Мозги
- Социальная жизнь
- Язык и символы
- Люди меняют мир
- Образование
- Введение в эволюцию человека
- Планы урока
- Классификация гаек и болтов: произвольно или нет? (6-8 классы)
- Сравнение хромосом человека и шимпанзе (9-12 классы)
- Сравнение черепов гоминидов: лаборатория «Черепа» (9–12 классы)
- Исследование общего происхождения: формулирование объяснений и моделей (9–12 классы)
- Окаменелости и модели миграции у ранних гоминидов (9–12 классы)
- Для студентов колледжа
- Забавные факты
- Почему у нас мурашки по коже?
- Куры, шимпанзе и вы — что у них общего?
- Бабушки и дедушки уникальны для людей
- Насколько мы сильны?
- Люди удобные!
- Люди: бегущая обезьяна
- Наш большой голодный мозг!
- Наши глаза говорят это!
- Набор инструментов раннего человека
- Короткошерстный человек!
- Щелкунчик»
- Что вши могут рассказать нам об эволюции человека?
- При чем здесь кишечник?
- Почему палеоантропологи любят Люси?
- Зачем нам зубы мудрости?
- Глоссарий происхождения человека
- Преподавание эволюции на человеческих примерах
- Часто задаваемые вопросы
- Рекомендуемые книги
- Экспонат
- Экспонат
- Экспонат План этажа Интерактивный
- Распечатать план этажа PDF
- Реконструкции ранних людей
- Передвижная выставка «Происхождение человека»
- Публичная библиотека округа Честерфилд
- Библиотека округа Ориндж
- Публичная библиотека Андовера
- Публичная библиотека Эфраты
- Публичная библиотека Эльвайна
- Публичная библиотека Сидар-Сити
- Библиотека Милпитаса; Милпитас, Калифорния
- Библиотека округа Спокан
- Публичная библиотека Коттедж-Гроув
- Библиотека округа Пуэбло
- Библиотека округа Спрингфилд-Грин
- Публичная библиотека Пеории
- Публичная библиотека городка Орион
- Публичная библиотека Скоки
- Бесплатная публичная библиотека Вайкоффа
- Публичная библиотека округа Томпкинс
- Библиотека Отиса
- Бесплатная библиотека Флетчера
- Публичная библиотека Бангора
- Путеводитель по выставке
- Экспонат
- О нас
- Благодарности
- События
- Команда программы «Происхождение человека»
- Комитет по более широкому социальному воздействию
- Что мы делаем
- Участники и ресурсы для участников
- Конни Бертка
- Джим Миллер
- Франциска Чо (заслуженный)
- Эллиот Дорфф
- Дэвид Хаберман
- Бетти Холли
- Нэнси Хауэлл
- Фред Эдуордс (почетный)
- Рэнди Исаак (почётный)
- Уэс Маккой
- Ли Медоуз
- Мустансир Мир
- Питер Ф.
9 Для науки
Что говорят…
Другие одобрения
Стать спонсором…

OneZoom — зарегистрированная некоммерческая организация в Великобритании, все наши продукты доступны бесплатно. 805 таких же, как вы, помогли нам своими пожертвованиями. Их имена появились в качестве соавторов на 1433 листьях нашего древа жизни.
Только одно имя на листе, так что покупайте любимое, пока есть возможность, для себя или в подарок.
Чем мы занимались…
О нас
Быстрые ссылки
Подпишитесь на нас
Электронная почта
Twitter
Facebook
Об OneZoom
Наша миссия
Наша миссия состоит в том, чтобы продвигать образование общественности по вопросам эволюции, биоразнообразия и сохранения разнообразия жизни на Земле.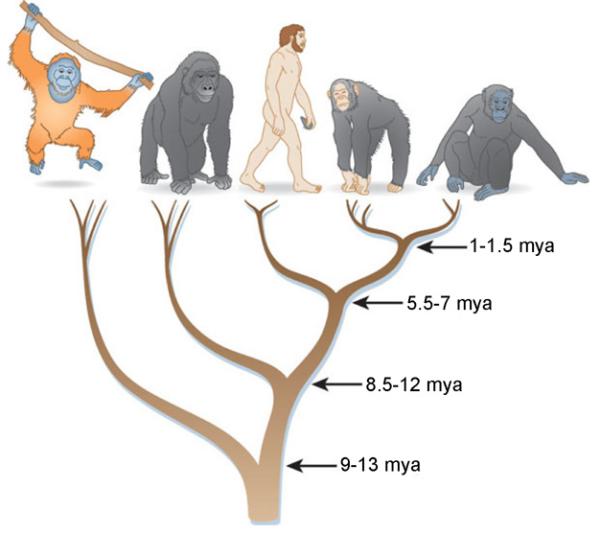 Мы некоммерческая организация, зарегистрированная в Великобритании.
Мы некоммерческая организация, зарегистрированная в Великобритании.
Наше программное обеспечение
Версия OneZoom 3.6-158-ga94e5846
Программное обеспечение OneZoom позволяет исследовать древо жизни совершенно по-новому, подобно просмотру географической карты. Это делается с помощью методов из раздела математики, известного как фрактальная геометрия, которые позволяют нам сжать все древо жизни на одной странице. OneZoom назван так потому, что вся информация находится на одной странице, все, что вам нужно сделать, это увеличить масштаб, чтобы открыть дополнительные детали.
Мы надеемся, что вы получите удовольствие от изучения дерева жизни OneZoom. Мы, безусловно, получили удовольствие от его разработки, даже после многих тысяч часов работы с OneZoom мы все еще часто удивляемся тому, что он рассказывает об окружающем нас мире. Большая часть нашего исходного кода общедоступна, что позволяет заинтересованным разработчикам и ученым вносить улучшения, если они того пожелают.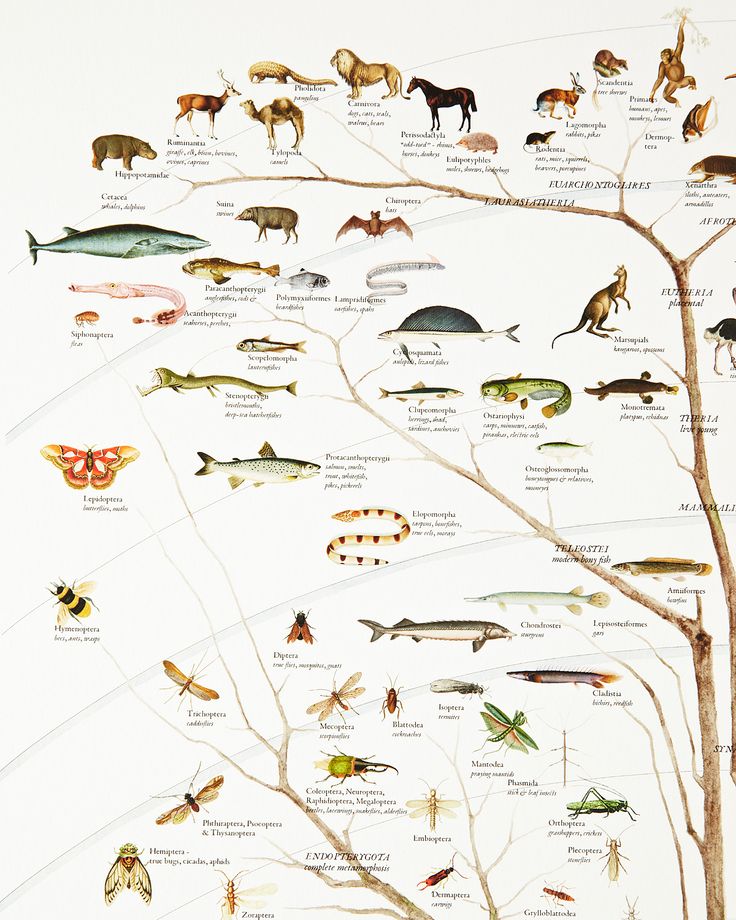
Организация OneZoom
OneZoom является благотворительной организацией (некоммерческой), зарегистрированной под номером 1163559.в Соединенном Королевстве. Мы стремимся обеспечить легкий доступ к научным знаниям о биоразнообразии и эволюции, а также повышать осведомленность о разнообразии жизни на Земле и необходимости ее сохранения. Преследуя эти цели, мы в настоящее время сосредоточены на развитии этой визуализации древа жизни. Наш зарегистрированный адрес: OneZoom, Office 7, 35-37 Ludgate Hill, London, EC4M 7JN, UK.
Проект OneZoom имеет долгую историю развития, начиная с первой версии OneZoom, которая была задумана в 2011 году и выпущена под лицензией с открытым исходным кодом в 2012 году. В настоящее время кодовая база OneZoom улучшается и поддерживается благотворительной организацией OneZoom с момента основания благотворительной организации. формирование в 2015 году.
Наши сотрудники
Компания OneZoom рада сотрудничеству с Лондонским Линнеевским обществом с 2017 года. разнообразие и доступность
разнообразие и доступность
OneZoom придерживается принципов равенства и разнообразия. Мы не будем дискриминировать по признаку возраста, инвалидности, пола, смены пола, расы, религии или сексуальной ориентации. Наши учебные материалы открыты для всех, и мы предпринимаем шаги, чтобы сделать их доступными настолько, насколько это целесообразно для проекта нашего масштаба. Недавние шаги включают дружественный режим для дальтоников для нашего главного исследователя древа жизни и добавление заголовка «Mx» на нашу спонсорскую страницу для людей с небинарной гендерной идентичностью. Мы приветствуем все отзывы о том, как мы можем сделать нашу работу более доступной.
Спонсорская схема
Имена жертвователей благотворительной организации OneZoom могут появляться на листьях дерева. Это благодарность за индивидуальную поддержку, это не реклама и не ссылки на внешние страницы. Пожертвования способствуют достижению общих целей благотворительной организации OneZoom. Обратите внимание, что пожертвования не вносят прямого вклада (например) в сохранение определенного вида, и при этом сами виды не названы в честь спонсоров (за исключением того, что иногда люди спонсируют вид, который уже был назван в их честь или который совпадает с их именем).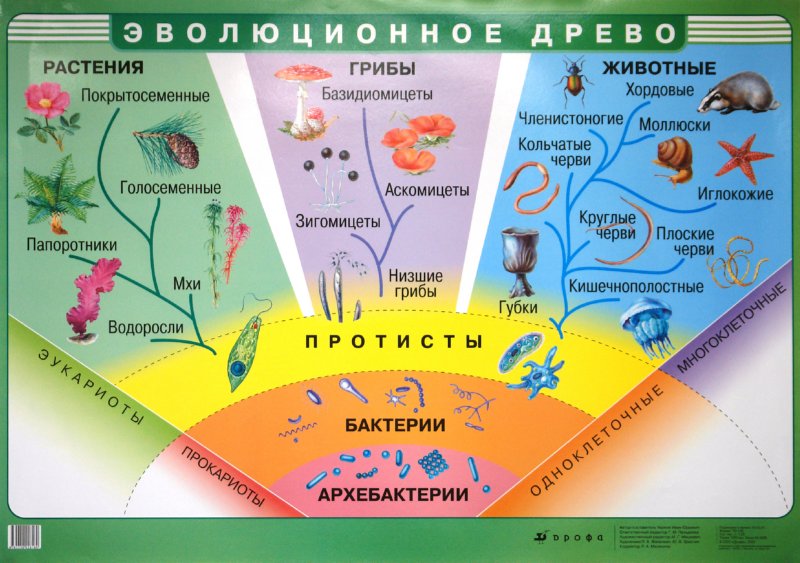 ).
).
Мы особенно благодарны профессору Ричарду Докинзу и профессору Джонатану Дрори, которые сделали самые щедрые пожертвования для спонсорства.
Для каждого вида (листа) дерева допускается только один спонсор, спонсорство длится 4 года и включает первый отказ при продлении. Более популярные виды требуют больших пожертвований на спонсорство. Вы можете спонсировать, выбирая листья в проводнике дерева, или, если вы просматриваете в общественном месте, где эта функция может быть отключена, вы можете спонсировать на веб-сайте onezoom.org. Вы можете искать дерево по именам спонсоров, а также по названиям видов, если вы хотите найти конкретный спонсируемый лист дерева.
Финансирующие органы
OneZoom нуждается в финансировании, чтобы продолжить свою работу. Мы стремимся заинтересовать частные фонды, исследовательские советы, музеи и другие организации, которые, как мы надеемся, будут рады участвовать в OneZoom. Если вы работаете в организации, которая потенциально заинтересована в дальнейшем развитии OneZoom, свяжитесь с нами, мы будем рады получить от вас известие.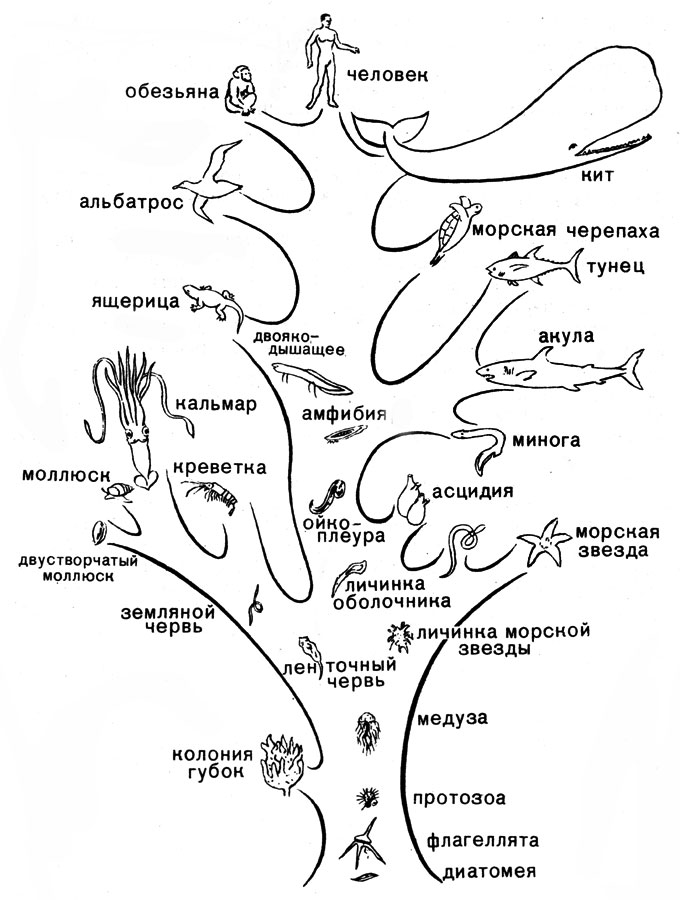
Как цитировать OneZoom
Если вы использовали OneZoom в обучении или исследованиях и хотите процитировать проект, используйте
Основная команда OneZoom (2021 г.). OneZoom Tree of Life Explorer Version 3.5 URL: http://www.onezoom.org
Мы также признательны за цитирование наших научных публикаций о разработке OneZoom
Другие ссылки
Данные, использованные для построения основного дерева на этом сайте, получены из вручную собранных источников, но в значительной степени опираются на проект Open Tree of Life, которому мы чрезвычайно благодарны.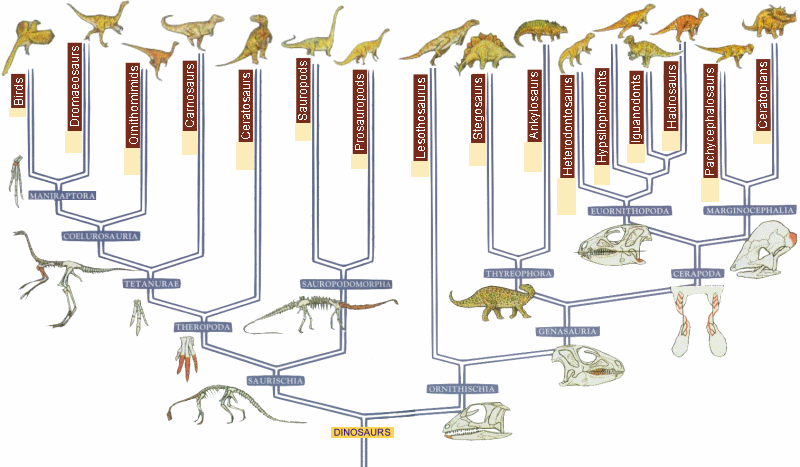
Изображения видов на дереве были собраны из Интернета проектом «Энциклопедия жизни» (EoL). Мы должны поблагодарить EoL за их фантастический ресурс и полезные ответы на наши различные запросы. Для получения дополнительной информации о любом изображении увеличьте значок копирайта рядом с изображением, этот символ также может служить ссылкой на источник изображения.
В текущей версии красные листья дерева соответствуют видам, находящимся под угрозой исчезновения согласно Красному списку исчезающих видов МСОП.
Фрактальный макет для нашего «политомического» древовидного представления, также используемого в проекте «Одно дерево — одна планета», похож на Lifemap: Exploring the Entire Tree of Life PLoS Biol 14(12): e2001624 (2016) и черпает вдохновение из него. Дэмиен М де Вьен.
Люди и кредиты
Пожалуйста, посетите нашу страницу команды на странице команды для получения полного списка нынешних и бывших членов команды OneZoom
Мы благодарим Джейми Лентина за его огромный вклад в кодовую базу OneZoom и особенно за разработку версии 3.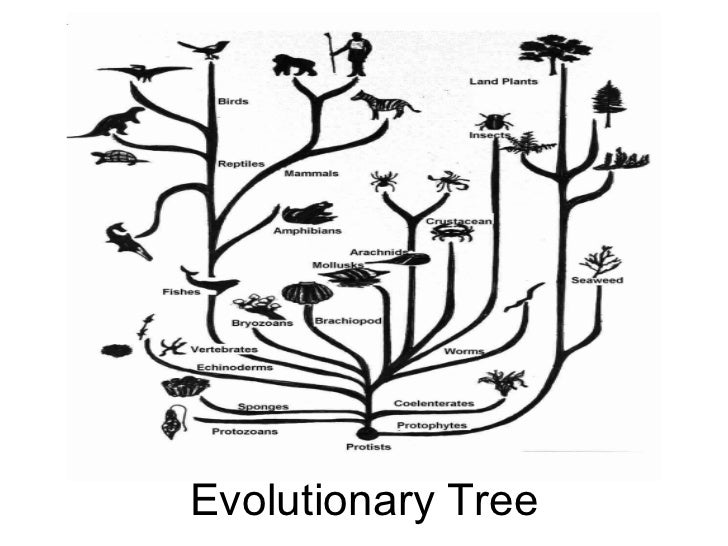 3 для поддержки Проект «Одно дерево — одна планета».
3 для поддержки Проект «Одно дерево — одна планета».
Мы благодарим Кай Чжуна за его огромный вклад в кодовую базу OneZoom и особенно за рефакторинг кода и добавление динамического кэширования, позволяющее отображать полное древо жизни.
Мы благодарим всю команду проекта One Tree One Planet за их поддержку в дальнейшем развитии OneZoom, чтобы сделать его пригодным для использования в рамках One Tree One Planet: Назиху Местауи, Дугласа Солтиса, Памелу Солтис, Роберта Гуралника и Мэтта Гитценданнера
Мы поблагодарить Линнеевское общество Лондона за их поддержку в качестве соавтора, за размещение OneZoom на многочисленных мероприятиях и за продвижение Древа Линнеевского общества в OneZoom.
Мы благодарим Джонатана Саттона за его значительный вклад в кодовую базу OneZoom во время его летнего размещения в 2016 году.
Мы благодарим Элен Морлон за ее поддержку в продвижении и развитии проекта OneZoom с тех пор, как она присоединилась к команде в 2016 году.
Мы благодарим Джонатана Дрори за его советы и поддержку в управлении благотворительной организацией OneZoom.
Мы благодарим профессора Ричарда Докинза и профессора Элис Робертс за их любезную постоянную рекламу OneZoom на мероприятиях и в социальных сетях.
Мы благодарим профессора Дункана Гиллиса, который помогал Джеймсу в руководстве и найме Кай Чжуна.
Мы также хотели бы поблагодарить следующих людей (в алфавитном порядке) за предоставление широкого спектра помощи и советов на различных этапах истории проекта OneZoom: Остин Берт, Джозеф Браун, Ной Констант, Стивен Корнелл, Джоэл Кракрафт, Мик Кроули, Джонатан Истман, Рампал Этьен, Роберт Эверс, Рич ФитцДжон, Джеймс Фостер, Мэтт Гитценданнер, Марк Хаммонд, Лиза Хармон, Мишель Харрисон, Клаас Хартманн, Джейкоб Хепуорт-Белл, Дэвид Хиллис, Ник Исаак, Шон Джордан, Уолтер Джетц, Джефф Джой, Джеймс Кирстед, Майк Кипарски, Билл Кунин, Саймон Леви, Кертис Лайл, Джорджина Мейс, Катриона МакКаллум, Линси Макиннес, Арне Мурс, Джек Ниджар, Лаура Нуньес, Дэвид Орм, Гильермо Орти, Ян Оуэнс, Род Пейдж, Матишалин Патель, Уильям Пирс, Мэтт Пеннелл, Альберт Филлимор, Жаклин Филлимор, Энди Первис, Алекс Пайрон, Дональд Куик, Эрика Бри Розенблюм, Джилл Розинделл, Оуэн Розинделл, Мартин Сэйерс, Нилай Шах, Грэм Слейтер, Дуг Солтис, Мэтью Спейт, Дэвид Танк, Гэвин Томас, Нина Торнхилл, Оливер Уорсфолд, Эми Зэнн
Все версии программного обеспечения OneZoom, начиная с 2. 0 и выше, были разработаны совместно доктором Джеймсом Розинделлом, который в основном занимался визуализацией, и доктором Яном Вонгом, который в основном занимался синтезом данных дерева. Ян является соавтором с профессором Ричардом Докинзом книги «История предков : история жизни», в которой широко использовались визуализации OneZoom. Исследование, проведенное для книги, в свою очередь, способствовало набору данных, используемому программным обеспечением OneZoom.
0 и выше, были разработаны совместно доктором Джеймсом Розинделлом, который в основном занимался визуализацией, и доктором Яном Вонгом, который в основном занимался синтезом данных дерева. Ян является соавтором с профессором Ричардом Докинзом книги «История предков : история жизни», в которой широко использовались визуализации OneZoom. Исследование, проведенное для книги, в свою очередь, способствовало набору данных, используемому программным обеспечением OneZoom.
Джеймс изначально разработал концепцию OneZoom в 2011 году. Первоначально он работал над проектом только в свободное время, в частности, прислушиваясь к советам Люка Хармона. После этого Джеймс продолжил разработку OneZoom в рамках своей исследовательской программы в Имперском колледже Лондона, которая финансировалась исследовательской стипендией (NERC). Первая версия 1.0 программного обеспечения и связанная с ней научная статья, авторами которой являются Джеймс и Люк, были опубликованы в 2012 г. Все текущие мероприятия OneZoom управляются независимой благотворительной организацией OneZoom с момента ее основания в 2015 г.
Генеалогическое древо человека | Программа Смитсоновского института по происхождению человека
Генеалогическое древо человека | Программа Смитсоновского института по происхождению человекаПерейти к основному содержанию