Насос ЦНС 105-147 | aliphant-group.com.ua
Насос ЦНС 105-147 – многоступенчатый центробежный секционный насос для подачи горячей или холодной воды от 1° до 45°С с минимальным содержанием абразивных примесей (не более 0,21% по массе и размером твердых частиц не более 0,22 мм), при этом подвод воды к насосу должен быть с напором более 10 м.в.ст. Горизонтальное исполнение насоса ЦНС 105-147 гарантирует высокую продуктивность работы.
Корпус секционного насоса ЦНС(г) состоит из отдельных секций, количество которых меньше числа рабочих колес на единицу, поскольку первое колесо располагается в передней крышке насоса и является стандартным заводским комплектующим насоса ЦНС (ЦНС(г)). Напор насоса будет равняться сумме напоров, создаваемых рабочими колесами. При этом отдельное рабочее колесо серии ЦНС105 и ЦНСг105 дает напор 49 м.вод.ст. Рабочие колеса размещены в одном корпусе и смонтированы на одном валу с подшипниками. Конструкция насоса ЦНС дает возможность уменьшать или увеличивать напор, сохраняя при этом номинальную подачу агрегата.
Секционные насосы ЦНС широко применяются для откачивания воды в шахтах, непосредственно в системах водоснабжения, а также в гидросистемах для создания и поддержания давления.
Маркировка насоса ЦНС 105-147/ЦНСг 105-147 (условные обозначения):
• ЦНС — центрабежный насос секционный:
• подача — 105 м³/ч;
• напор — 147 м.
Насосы многоступентчатые ЦНС могут агрегатироваться электродвигателем взрывозащищенного исполнения. Стандартная комплектации насоса – общепромышленный электродвигатель.
Основным производителем секционных насосов ЦНС 105-147 является Россия. Производитель насосных агрегатов ЦНС гарантирует работоспособность собственной продукции на протяжении 18 месяцев. Наша компания обеспечивает поставку запасных частей, комплектов ЗИП, ремонт насосов ЦНС собственного производства на базе завода.
Наше предприятие имеет опыт разработки систем водоснабжения, систем отвода воды с шахт, систем водоканалов посредством использования насосов ЦНС и ЦНСг. В основном серия центробежных насосов ЦНС 105-147 и ЦНСг 105-147 используется на каменных и угольных шахтах. Угледобывающие предприятия Украины и СНГ активно используют насосы ЦНС отечественного производства на замену иностранным насосным агрегатам. Насосы ЦНС – незаменимый аналог насосных агрегатов для решения задач перекачивания воды и схожих с ней рабочих жидкостей.
В основном серия центробежных насосов ЦНС 105-147 и ЦНСг 105-147 используется на каменных и угольных шахтах. Угледобывающие предприятия Украины и СНГ активно используют насосы ЦНС отечественного производства на замену иностранным насосным агрегатам. Насосы ЦНС – незаменимый аналог насосных агрегатов для решения задач перекачивания воды и схожих с ней рабочих жидкостей.
Цену на насос, насосный агрегат, раму на насос, корпус насоса (улитку насоса), рабочие колеса насоса (крыльчатку), ротор в сборе насоса ЦНС и прочие запчасти уточняйте у менеджера отдела продаж нашей компании.
Насос ЦНС 105-147 предназначен для перекачивания химически нейтральной холодной воды, имеющей водородный показатель (рН) 7-8,5, с массовой долей механических примесей не более 0,2% и размером твердых частиц не более 0,2 мм, микротвердостью не более 1,46 ГПа и плотностью не более 1500 кг/м³, с температурой до 45° С (ЦНСг до 105° С). Насос ЦНС 105-147 — центробежный горизонтальный высоконапорный многоступенчатый насос секционного типа с колесами одностороннего входа. Входной патрубок насоса направлен горизонтально, напорный — вертикально вверх. Концевое уплотнение вала насоса — двойное сальниковое. Конструкцией торцовых уплотнений предусмотрена возможность их периодической промывки от продуктов эрозии, коррозии и других взвешенных частиц. Опорами ротора насоса служат подшипники скольжения с картерной либо принудительной смазкой. В качестве соединительной муфты между двигателем и насосом используется упругая пластинчатая муфта, обеспечивающая снижение виброактивности. Для охлаждения подшипников предусмотрена полость в корпусе. Материал деталей проточной части насоса ЦНС 105-147: рабочие колеса, направляющий аппарат изготовлены из углеродистой стали 35Л, корпус направляющего аппарата изготовлен из серого чугуна марки СЧ-20. Максимальное допустимое давление на входе в насос не более 0,25 МПа ( 2,5 кгс/см²). Насос выпускается в климатическом исполнении УХЛ и категории размещения при эксплуатации 4 по ГОСТ 15150-69. Характеристики насоса ЦНС(г) 105-147: ООО “ФАСТЕХ” уже несколько лет является одним из ведущих поставщиков насосного оборудования, такого как секционные насосы, высоконапорные насосы, насосы для чистой воды, насосы для систем холодного и горячего водоснабжения и др., а так же предлагает такую востребованную услугу, как ремонт насосов. |

Насос ЦНС 105-147
Насос многоступенчатый секционный ЦНС 105-147 (ЦНСг 105-147)
— центробежный, горизонтальный, многоступенчатый, высоконапорный, однокорпусной секционного типа с колесами одностороннего входа и гидравлическим устройством разгрузки осевого усилия ротора (гидропятой). Используется в системах водоснабжения, а также в контурах холодной воды жилых зданий, промышленных объектов, для подачи воды в паровые котлы котельных ТЭЦ, для подачи воды в нефтеносные пласты, а также для водоотлива каменноугольных шахт.
Используется в системах водоснабжения, а также в контурах холодной воды жилых зданий, промышленных объектов, для подачи воды в паровые котлы котельных ТЭЦ, для подачи воды в нефтеносные пласты, а также для водоотлива каменноугольных шахт.Насос ЦНСг 105-147 предназначен для перекачивания нейтральной горячей воды в системах горячего водоснабжения промышленных, административных и жилых объектов.
Условия эксплуатации насоса ЦНС 105-147
Перекачиваемая среда — чистые и нефтепромысловые воды не содержащие сероводород, с температурой от 1 до 45 °С (ЦНСг 105-147 — от 45 до 105 °С), с водородным показателем pH 7-8,5, содержанием твердых включений размером до 0,1 мм, микротвердостью не более 1,47 гПа (14,7 кгс/см²), объемная концентрация которых не превышает 0,1%. Максимально допустимое давление на входе — 0,3 МПа, минимальное — 0,02-0,06 МПа. Для насоса ЦНСг 105-147 давление на входе не должно быть менее 0,1 МПа.Насос ЦНС 105-147 изготавливается в общепромышленном исполнении для эксплуатации в помещениях взрывоопасной зоны класса В-1б или пожароопасной зоны класса П-1 в соответствии с «Правилами устройства электроустановок» (ПУЭ).
 Климатическое исполнение УХЛ, категория размещения 4 по ГОСТ 15150.
Климатическое исполнение УХЛ, категория размещения 4 по ГОСТ 15150.Технические характеристики насоса ЦНС 105-147
| Типоразмер | Подача, м³/ч; | Напор, м | Число ступеней насоса | Допускаемый кавитационный запас, м | Мощность, кВт | КПД, % | Частота вращения, об/мин | Масса насоса, кг |
| ЦНС 105-147 | 105 | 147 | 3 | 5,5 | 75 | 65 | 3000 | 518 |
Габаритные размеры насоса ЦНС 105-147
| Типоразмер | L | L1 | L2 | L3 | L4 | L5 | B | B1 | H | H1 | H2 | d |
| ЦНС 105-147 | 2010 | 310 | 1500 | 800 | 350 | 346 | 468 | 400 | 815 | 660 | 430 | 28 |
Устройство насоса ЦНС 105-147
- Насос ЦНС 105-147 состоит из корпуса и ротора.
 К корпусу относятся всасывающая и нагнетательная крышки, корпуса направляющих аппаратов с направляющими аппаратами и кронштейн. Корпуса направляющих аппаратов, крышки всасывания и нагнетания стягиваются стяжными шпильками.
К корпусу относятся всасывающая и нагнетательная крышки, корпуса направляющих аппаратов с направляющими аппаратами и кронштейн. Корпуса направляющих аппаратов, крышки всасывания и нагнетания стягиваются стяжными шпильками. - Стыки корпусов направляющих аппаратов уплотняются резиновыми кольцами средней твердости (ГОСТ 6467-79). Исполнение колец зависит от назначения насоса.
- Ротор насоса состоит из вала, на котором установлены рабочие колеса, дистанционная втулка и диск гидравлической пяты. Все эти детали стягиваются на валу гайкой вала. Места выхода вала из корпуса уплотняются сальниковой набивкой, пропитанным антифрикционным составом.
- Опорами ротора служат два радиальных сферических подшипника (№1608 ГОСТ 5720-75), которые установлены в кронштейнах, по скользящей посадке, позволяющей перемещаться ротору в осевом направлении на величину «хода» ротора. Места выхода вала из корпусов подшипников уплотняются манжетами. Подшипниковые камеры закрыты крышками, закрепляемыми болтами и гайками.
- Корпус направляющего аппарата, аппарат направляющий, колесо рабочее, кольца уплотняющие в своей совокупности образуют секцию насоса.
- Для уравновешивания осевого усилия в насосе применяется гидравлическая пята, состоящая из диска гидравлической пяты, кольца гидравлической пяты, втулки разгрузки и втулки дистанционной.
- Ротор насоса приводится во вращение от электродвигателя через упругую втулочно-пальцевую муфту, состоящую из двух полумуфт, которые соединяются между собой через резиновые втулки, установленные на цилиндрические стальные пальцы, жестко закрепляемые в полумуфте электродвигателя.
- Конструкция насоса ЦНСг 105-147 для горячей воды предусматривает охлаждение подшипников водой от постороннего источника за счет сквозных резьбовых отверстий в нижней части переднего и заднего кронштейнов. Также в нем отсутствует втулка гидрозатвора, устройство для выпуска воздуха и система обводнения.
 К корпусу относятся всасывающая и нагнетательная крышки, корпуса направляющих аппаратов с направляющими аппаратами и кронштейн. Корпуса направляющих аппаратов, крышки всасывания и нагнетания стягиваются стяжными шпильками.
К корпусу относятся всасывающая и нагнетательная крышки, корпуса направляющих аппаратов с направляющими аппаратами и кронштейн. Корпуса направляющих аппаратов, крышки всасывания и нагнетания стягиваются стяжными шпильками.

| Типоразмер | Входной патрубок, мм | Напорный патрубок, мм | ||||||||||||||
| Ду | Д1 | Д2 | Д3 | Д4 | Д5 | h | n | Ду | Д1 | Д2 | Д3 | Д4 | Д5 | h | n | |
| ЦНС 105-147 | 125 | 152 | 170 | 190 | 220 | 270 | 27 | 8 отв Ø24 | 100 | 128 | 145 | 160 | 200 | 250 | 30 | 8 отв Ø26 |
Подготовка насоса ЦНС 105-147 к работе
- Проверните ротор насоса вручную и убедитесь в отсутствии заклинивания. Проверьте установку ротора по риске. Проверка положения риски производится при роторе, сдвинутом до упора в сторону всасывания. Риска должна быть заподлицо с торцовой плоскостью крышки подшипника переднего кронштейна.
- Набивку и подтяжку сальников производите таким образом, чтобы при работе насоса между валом (гайкой вала) и сальником протекала перекачиваемая жидкость в количестве 15-30 л/ч.
- При получении насоса без электродвигателя перед монтажом произвести расточку ступицы полумуфты электродвигателя и изготовить шпоночный паз по соответствующим размерам вала электродвигателя.
- Проверьте центровку насоса и электродвигателя и правильность направления вращения электродвигателя. Ротор электродвигателя должен вращаться против движения часовой стрелки, если смотреть со стороны приводного конца вала. Обратное вращение ротора не допускается. При неправильном вращении в соответствии с эксплуатационной документацией на электродвигатель установите обратное вращение.
Внимание! Вставляйте пальцы в муфту только после того, как убедитесь в правильности направления вращения ротора электродвигателя. - Проверьте наличие смазки в подшипниковых камерах, сняв крышки подшипников.
- После проверки исправности и готовности к действию приступают к заливке насоса и всасывающего трубопровода жидкостью из нагнетательного трубопровода или через отверстие М20х1,5 в крышке всасывания.
В крышке всасывания насосов ЦНС 105-147 вмонтирован клапан для выпуска воздуха. При заливке насоса клапан поднимается, и воздух выпускается через клапан. В рабочем положении клапан прижимается к гнезду. Насос заливают до тех пор, пока через сливную трубку начнет бить струйка жидкости без воздушных пузырьков. Одновременно проверяют герметичность всасывающего трубопровода, приемного клапана, системы обводнения, пробок и устраняют места течи.
 Риска должна быть заподлицо с торцовой плоскостью крышки подшипника переднего кронштейна.
Риска должна быть заподлицо с торцовой плоскостью крышки подшипника переднего кронштейна.
Техническое обслуживание насоса ЦНС 105-147
- При эксплуатации электронасосного агрегата необходимо вести наблюдения за его техническим состоянием, режимами работы
насоса и электрооборудования, нагревом подшипников, за внешними утечками через гидравлическую пяту и сальники и периодически производить техническое обслуживание.
- При техническом обслуживании электрооборудования необходимо руководствоваться эксплуатационной документацией на электрооборудование.
- При техническом обслуживании насоса ЦНС 105-147 выполняйте следующие работы:
а) проверяйте правильность центровки валов насоса и электродвигателя;
б) следите за величиной износа деталей гидравлической пяты по риске;
— при выходе риски от торца крышки подшипника на величину более 3ч5 мм разберите устройство гидравлической пяты, снимите одно или несколько регулировочных колец суммарной толщиной равной величине смещения ротора и поставьте его (их) между диском гидравлической пяты и гайкой вала.
При значительном износе деталей гидравлической пяты замените их без снятия регулировочных колец.
— сборку насоса производите в последовательности, обратной разборке;
— проверьте положение риски. - Производите не реже, чем через 200 часов работы насоса пополнение камеры подшипников смазкой, а через 500 часов работы производите полную смену смазки. Для смазки подшипников используется Литол 24 ГОСТ 21150-75.

 Для смазки подшипников используется Литол 24 ГОСТ 21150-75.
Для смазки подшипников используется Литол 24 ГОСТ 21150-75.Пример условного обозначения: Насос ЦНС 105-147-1С УХЛ4, где
ЦНС — тип насоса;
105 — подача в номинальном режиме, м³/ч;
147 — напор в номинальном режиме, м;
1 — конструктивное исполнение;
С — тип уплотнения вала насоса — сальниковое;
УХЛ — климатическое исполнение для районов с умеренным и холодным климатом по ГОСТ 15150-69;
4 — категория размещения насоса при эксплуатации по ГОСТ 15150-69.
ООО “Компани “Плазма”® является одним из ведущих поставщиков насосов и насосных агрегатов, таких как насосы для чистой или загрязненной воды, насосы для абразивных жидкостей, химические и нефтяные насосы, гидравлические насосы и т.д. Если Вам необходимо купить насос для перекачивания чистой воды, насос для систем водоснабжения или отопления или другое промышленное оборудование, свяжитесь с нашими специалистами, и получите более подробную информацию.
Насос ЦНС 105-147 насос ЦНСг 105-147 ЦНС 105 147 цена от производителя
Насос ЦНС 105-147
Производительность 105 м3/час ( рабочая зона 80-130 м3/час)
Напор 147 метра ( рабочая зона 122-160 метра)
Кавитационный запас насоса 5,5 метра
Электродвигатель 75 кВт 3000 об/мин АИР250S2 4АМ225S2
Габаритные размеры насоса ЦНС 105-147
Патрубки всасывающий (вход) и напорный (выход) 125 мм
Масса насоса 518 кг
Габаритные размеры насоса ЦНС 105-147 с электродвигателем
Дилина х Ширина х Высота в мм 2120х640х920
Масса агрегата 1140 кг
Ранее насос ЦНС 105-147 выпускался под маркировкой 5МС-10х2 5МСг-10х3
Насос ЦНС 105-147 на раме с двигателем
Насос ЦНС 105-147 горизонтальный, секционный центробежный насос, предназначен для перекачивания чистой и технической воды с температурой до 105 градусов, Рн 7-8,5, с мех.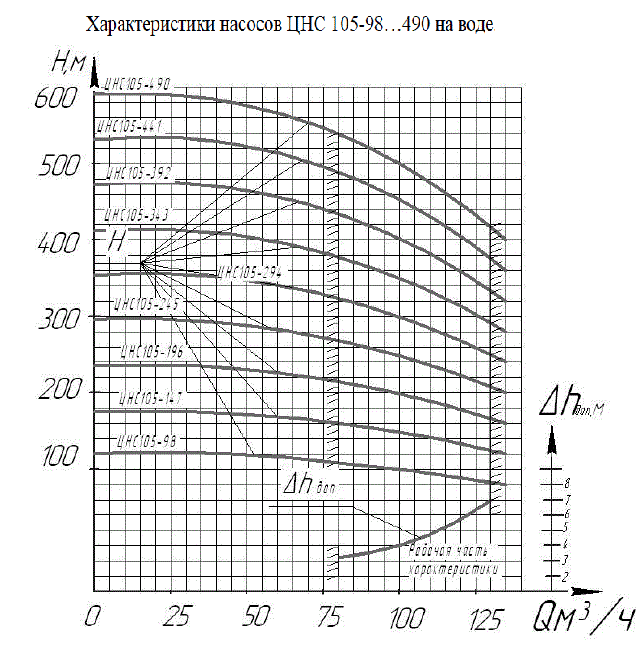 примесями не более 0,1 процента. Широко применяются в шахтах каменноугольных и системах подачи воды.
примесями не более 0,1 процента. Широко применяются в шахтах каменноугольных и системах подачи воды.
По конструкции насос ЦНС 105-147 представляет собой последовательно соединенные центробежные насосы , то есть секции с рабочими колесами, каждая секция добавляет напор если рассматривать ЦНС-105 на 49 метра напора.
Также для ремонта насоса ЦНС 105-147 запчасти : аппарат направляющий ЦНС 105-147, вал секционный ЦНС 105-147, втулка гидрозатвора ЦНС 105-147, втулка дистанционная ЦНС 105-98, втулка подшипника ЦНС 105-147, диск разгрузки ЦНС 105-147, рабочее колесо ЦНС 105-147, гайка ротора ЦНС 105-147, полумуфта насоса ЦНС 105-147,
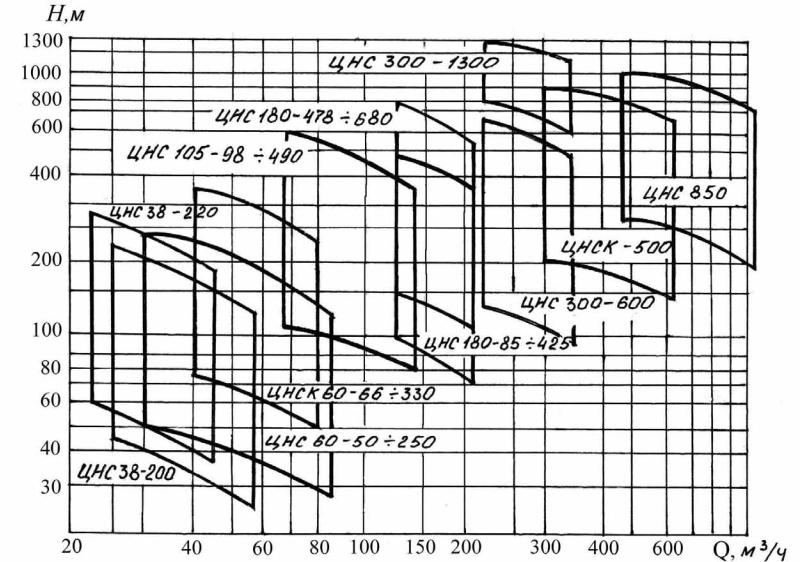
Основная линейка насосов ЦНС-105 представленная ниже : ЦНС 105-490 ЦНС 105-441 ЦНС 105-392 ЦНС 105-343 ЦНС 105-294 ЦНС 105-245 ЦНС 105-196 ЦНС 105-147 ЦНС 105-98
Насос ЦНС 105-98 Чернигов, Нежин, Конотоп, Коростень, Прилуки, Киев, Лубны, Черкассы, Кременчуг, Харьков, Днепропетровск, Луганск, Мариуполь, Запорожье, Кривой Рог, Николаев, Винница, Львов, Полтава, Поти, Тбилиси, Габала, Губа , Шемаха, Ширван, Баку, Ащхабад, Туркменабад, Бухара, Киши, Душанбе, Караганда, Астана, Алматы, Кишинёв и другие города Украины, Грузии, Азербайджана, Киргызстана, Казахстана, Молдовы
Насос ЦНС 105-147 центробежный, секционный в Украине
Насос ЦНС 105-147 используется при перекачивании холодной или горячей чистой воды, а также жидкостей схожих по свойствам с чистой водой.
Допустимый режим работы ЦНС 105-147.
| Температура | до +45 оС ЦНС, до +105оС ЦНСГ |
| Давление на входе в насос | от 0,06 до 0,014 мПа |
| Размер твердых частиц | до 0,21мм |
| Примеси по массе от общей массы жидкости | до 0,5% |
Основные характеристики насосов ЦНС 105-147.
| Производительность, м3/ч | 105 |
| Напор, метров | 147 |
| Кавит. запас, метров | 5,5 |
| Электродвигатель кВт/об. в минуту | 75/3000 |
| Масса агрегата, кг | 1310 |
| Диаметр рабочего колеса, мм | 225 |
Области применения ЦНС 105-147
– жилищно-коммунальное хозяйство;
– сельскохозяйственные и животноводческие предприятия;
– шахты и горнодобывающие предприятия;
-теплоэнергетика;
– другие области.
Производители – «Ясногорский насосный завод» г. Ясногорск Россия, «Димитровградхиммаш» г. Димитровград Россия, «Катайский насосный завод» г. Катайск Россия, «Насосэнергомаш» г. Сумы Украина, «Ливгидромаш» г. Ливны Россия, и другие.
Конструкция насоса ЦНС 105-147
Основой конструкции является корпус, в котором расположены секции. Внутри секций установлены рабочие колеса, посаженные на вал.
Габариты ЦНС 105-147
Где купить насос ЦНС 105-147 в Украине?
Компания «Элнас», обладает мощностями по хранению, ревизии и ремонту ЦНСГ 105-147. Вы можете приобрести интересующий Вас ЦНС105-147, заказав его из любого города Украины или ближнего зарубежья. Поставка ЦНС105-147 осуществляется любым транспортным перевозчиком или собственным автотранспортом.
Все оборудование перед отправкой проверяется и при необходимости ревизируется. На все поставляемые насосы предоставляется гарантия 1 год. Специалисты компании «Элнас» гарантируют качество поставляемых насосов ЦНС105-147.
Специалисты компании «Элнас» гарантируют качество поставляемых насосов ЦНС105-147.
Насос ЦНС 105-147 секционный центробежный для воды ЦНСг
ООО «МонтажЭнергоИнвест» предлагает широкий ассортимент оборудования для систем водоснабжения, отопления, дренажа и канализации, водоочистки и фильтрации. Насосы консольные, двухстороннего входа, центробежные, винтовые, шестерные, маслонасосы, дренажные, самовсасывающие, грунтовые, шламовые, погружные, артезианские, вихревые, топливные, питательные, конденсатные, нефтяные, химические, фекальные, водяные. Насосные агрегаты. Широкий ассортимент запасных частей и комплектующих к насосам. Сальниковая набивка, торцевые уплотнения, пальцы полумуфт.
Насос ЦНС 105-147 относят к разновидности промышленных агрегатов, применяемых для перекачивания чистой холодной и горячей воды – в модификации ЦНСг 105-147
Насос ЦНС 105-147 Украина благодаря своим техническим характеристикам незаменим в отраслях ЖКХ, водного хозяйства, теплоэнергетики, промышленных систем, а также схожих по направлению отраслях. Особого внимания стоит использование насосов ЦНС 105-147 на шахтах.
Особого внимания стоит использование насосов ЦНС 105-147 на шахтах.
Насос ЦНС 105-147 купить обладает достаточно компактным размером, позволяющим устанавливать его в стационарных условиях на любом предприятии. Агрегат ЦНС 105-147 Украина – АЦНС 105-147 монтируется на фундамент, в его комплектацию входит голова, закрепленный на сварной раме, а также соединенный с ним приводной электродвигатель.
Насос ЦНСГ 105-147 – Конструкция насоса ЦНС 105-147 и чертеж насоса.
Назначение и конструкция:
В многоступенчатых насосах поток перекачиваемой жидкости перемещается последовательно несколькими рабочими колесами, смонтированными на одном валу, в одном корпусе.
Корпус многоступечатого секционного насоса состоит из отдельных секций (камеража), число которых равно числу рабочих колес минус один, так как одно колесо расположено в передней крышке. Секционная конструкция корпуса насоса позволяет увеличить или уменьшить напор, не изменяя подачи. Напор будет равен сумме напоров, создаваемых каждым рабочим колесом.
Используются в гидросистемах для создания большого давления. Допускается изготовление с торцовым уплотнением.
Перекачиваемая жидкость:
Насосы ЦНС предназначены для перекачивания воды с водородным показателем pH 7-8,5, с масовой долей механических примесей не более 0,1% и размером твердых частиц не более 0,1мм и температурой от 1 до 45°С. Допускается изготовления насоса с торцовым уплотнением.
Насос ЦНС 105-147 купить
Насос ЦНС 105-147 цена
Насос ЦНС 105-147 Украина
БЕСПЛАТНЫЙ ПОДБОР ИЛИ ДЕФЕКТОВКА ОБОРУДОВАНИЯ. УЧИТЫВАЕМ ВСЕ ВАШИ ИНТЕРЕСЫ. КУПИМ ВАШИ НЕИСПРАВНЫЕ ИЛИ Б/У НАСОСЫ.
| Возможность подачи предложений по части позиций Участники могут подавать предложение по отдельным позициям закупки : | Предусмотрена |
| Запрет подачи цены выше поданной ранее Организатором установлено ограничение подачи ценового предложения выше ранее поданного участником. | Да |
| Подгрузка документации к заявке обязательна Организатор не будет рассматривать заявки, которые не были подкреплены документацией. : | Да |
| Присвоение ранга предложениям В ходе процедуры ценовым предложениям участников присваивается ранг (место) по цене. | Да |
| Минимальный шаг Организатором установлен минимальный шаг изменения участником цены за единицу по каждой позиции от своего ранее поданного предложения. : | 0,01% от цены за единицу |
| Автоматическое продление Если в последние 30 минут до момента окончания срока подачи заявок поступит предложение от одного из участников, то срок окончания подачи заявок в рамках данной процедуры будет автоматически продлён еще на 30 минут с момента поступления последней заявки. | 30 минут |
| Условия оплаты: | 100% постоплата, 30 календарых дней |
| Условия поставки: | DDP Акмолинская область, Бурабайский район, Успеноюрьевский с/о (70 км от г.Щучинск) месторождение Райгородок |
| Адрес места поставки товара, проведения работ или оказания услуг: | Акмолинская область, Бурабайский район, Успеноюрьевский сельский округ, близ села Райгородок. |
| Комментарии: Прошу прекрепить КП, учредительные документы, реквизиты | |
| Место проведения процедуры: | Данная процедура проводится в электронной форме, предложения участников подаются только через функционал ЭТП ETS-Tender. |
| по всем вопросам обращаться:: | 8 771 040 66 02 Арман или написать arman.zheximbayev@rggold. kz kz |
 Изменить цену в сторону увеличения после отправки заявки будет невозможно.
Изменить цену в сторону увеличения после отправки заявки будет невозможно.
Влияние на пластичность, прорастание и регенерацию аксонов после травмы спинного мозга
По мере созревания нервной системы внеклеточная среда центральной нервной системы (ЦНС) становится высоко структурированной и организованной. Внеклеточное пространство ЦНС вместе с его субдоменами играет решающую роль в функционировании и стабильности ЦНС. В этом обзоре мы сосредоточились на двух компонентах внеклеточной среды нейронов, которые важны для регуляции пластичности ЦНС, включая внеклеточный матрикс (ЕСМ) и миелин.ЕСМ состоит из протеогликанов хондроитинсульфата (CSPG) и тенасцинов, которые организованы в уникальные структуры, называемые перинейрональными сетями (PNN). PNN связаны с телом нейронных клеток и проксимальными дендритами преимущественно парвальбумин-положительных интернейронов, образуя прочную решетчатую структуру. Эти регулируемые в процессе развития структуры поддерживаются в ЦНС взрослых и повышают синаптическую стабильность. Однако после травмы CSPG и тенасцины вносят вклад в структуру тормозящего глиального рубца, который активно предотвращает регенерацию аксонов.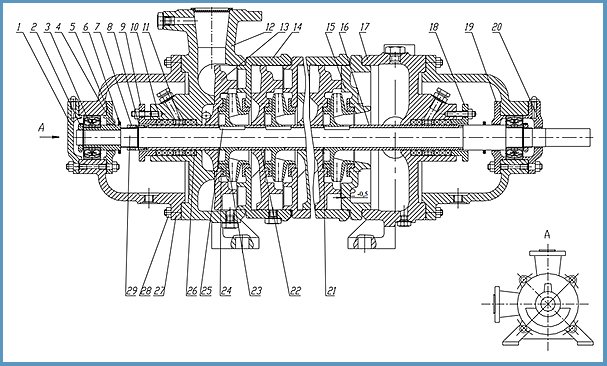 Миелиновые оболочки и зрелые олигодендроциты взрослых, несмотря на их важную роль в передаче сигналов в зрелых аксонах ЦНС, вносят вклад в тормозную среду, существующую после повреждения. Таким образом, в отличие от периферической нервной системы, ЦНС неспособна вернуться в «состояние развития», чтобы способствовать восстановлению нейронов. Однако было показано, что модуляция этих внешних факторов способствует росту, регенерации и функциональной пластичности после травмы. В этом обзоре будут освещены некоторые факторы, которые способствуют или предотвращают пластичность, прорастание и регенерацию аксонов после травмы спинного мозга.
Миелиновые оболочки и зрелые олигодендроциты взрослых, несмотря на их важную роль в передаче сигналов в зрелых аксонах ЦНС, вносят вклад в тормозную среду, существующую после повреждения. Таким образом, в отличие от периферической нервной системы, ЦНС неспособна вернуться в «состояние развития», чтобы способствовать восстановлению нейронов. Однако было показано, что модуляция этих внешних факторов способствует росту, регенерации и функциональной пластичности после травмы. В этом обзоре будут освещены некоторые факторы, которые способствуют или предотвращают пластичность, прорастание и регенерацию аксонов после травмы спинного мозга.
1. Введение
Восстановление центральной нервной системы (ЦНС) после травм — одна из самых больших проблем, с которыми сегодня сталкиваются нейробиологи. Наиболее частые причины травматического повреждения спинного мозга (ТСМ) можно предотвратить и включают дорожно-транспортные происшествия, падения, насилие и контактные виды спорта, в результате которых люди часто страдают различными типами сенсорных и / или двигательных нарушений, многие из которых теряют независимость. Травматическая ТСМ чаще встречается у мужчин, чем у женщин, и чаще всего возникает в молодом зрелом возрасте (15–29 лет), а также в пожилом возрасте (старше 60 лет) [1].Лечение ТСМ ограничено и часто направлено на предотвращение дальнейшего повреждения, при этом в настоящее время в клинике используются вмешательства, включающие реабилитацию [2].
Травматическая ТСМ чаще встречается у мужчин, чем у женщин, и чаще всего возникает в молодом зрелом возрасте (15–29 лет), а также в пожилом возрасте (старше 60 лет) [1].Лечение ТСМ ограничено и часто направлено на предотвращение дальнейшего повреждения, при этом в настоящее время в клинике используются вмешательства, включающие реабилитацию [2].
1.1. ECM и патофизиология после травмы ЦНС / аксонов
ЦНС не восстанавливается после травмы из-за множества тормозящих факторов. По сути, центральные нейроны взрослых ограничены в своей способности вызывать регенеративный ответ, частично из-за тормозящей среды в месте повреждения.Несколько исследователей продемонстрировали, что после травмы возникает начальная реакция роста; однако, как только аксоны сталкиваются с тормозной средой внутри поражения, рост останавливается, оставляя на их месте дистрофические концевые луковицы аксонов [3–5]. В нормальной ЦНС клетки окружены внеклеточным матриксом (ЕСМ), состоящим из сложной и интерактивной сети гликопротеинов и протеогликанов [6]. При различных обстоятельствах эти молекулы могут либо способствовать росту нейритов, например, во время развития нейронов [7], либо отталкивать его, например, после травмы [8–20] или во время болезней / дегенеративных состояний [21] (Рисунок 1).
При различных обстоятельствах эти молекулы могут либо способствовать росту нейритов, например, во время развития нейронов [7], либо отталкивать его, например, после травмы [8–20] или во время болезней / дегенеративных состояний [21] (Рисунок 1).
Кроме того, одна из основных причин, по которой спинной мозг не поддается восстановлению, связана со сложной и характерной патофизиологией, которая возникает после травмы. Биология, лежащая в основе SCI, состоит из первичной и вторичной фаз. Первичная фаза — это первоначальная травма, полученная при ударе. Это может включать давление на позвоночник, внутреннее кровотечение, а также разрыв аксонов и нервов. В зависимости от типа повреждения гематоэнцефалический барьер (ГЭБ) может быть нарушен, что приведет к инфильтрации лейкоцитов [22, 23], а также к нарушению ионного гомеостаза, что приведет к значительным изменениям кальция (Ca 2+ ) и калия ( K + ) в месте повреждения [24, 25].Кроме того, нарушение кровоснабжения места повреждения может привести к недостаточному снабжению пораженной тканью кислородом и питательными веществами.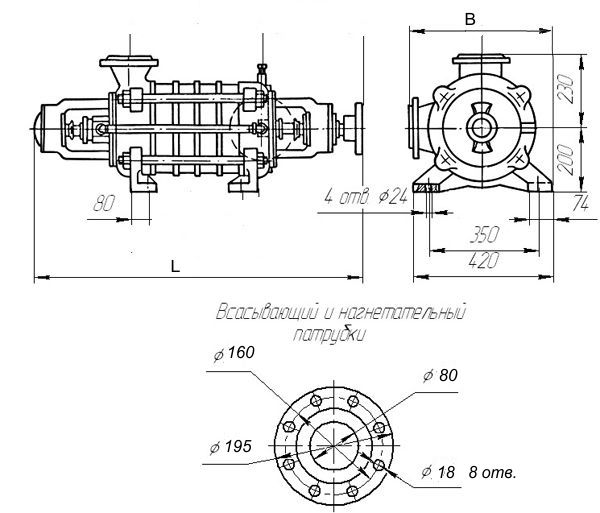 На этом этапе требуется неотложная помощь, чтобы предотвратить дальнейшее повреждение. Неотложное лечение включает стабилизацию позвоночника, за которой часто следует хирургическая декомпрессия, чтобы уменьшить воспалительную реакцию и предотвратить дополнительное кровотечение. В последующие недели и месяцы травма переходит во вторичную фазу, что приводит к прогрессирующей нейродегенерации. Эта фаза связана со вторым набором симптомов, включая воспаление, нейрональный и глиальный апоптоз, эксайтотоксичность глутамата, потенциальное нарушение ГЭБ, демиелинизацию аксонов и реактивный астроглиоз, приводящие к образованию глиального рубца.Наряду с образованием рубцов демиелинизация аксонов и дегенерация Валлера приводят к ретракции аксонов [26]. Ретракция аксонов происходит в две фазы: (1) внутренняя фаза раннего аксона, связанная с цитоскелетом, в которой Ca 2+ -зависимая активация протеаз кальпаина приводит к разрушению цитоскелета [27] и (2) фаза, зависимая от макрофагов, в котором инфильтрация фагоцитарных макрофагов вызывает / коррелирует с ретракцией дистрофических аксонов [28].
На этом этапе требуется неотложная помощь, чтобы предотвратить дальнейшее повреждение. Неотложное лечение включает стабилизацию позвоночника, за которой часто следует хирургическая декомпрессия, чтобы уменьшить воспалительную реакцию и предотвратить дополнительное кровотечение. В последующие недели и месяцы травма переходит во вторичную фазу, что приводит к прогрессирующей нейродегенерации. Эта фаза связана со вторым набором симптомов, включая воспаление, нейрональный и глиальный апоптоз, эксайтотоксичность глутамата, потенциальное нарушение ГЭБ, демиелинизацию аксонов и реактивный астроглиоз, приводящие к образованию глиального рубца.Наряду с образованием рубцов демиелинизация аксонов и дегенерация Валлера приводят к ретракции аксонов [26]. Ретракция аксонов происходит в две фазы: (1) внутренняя фаза раннего аксона, связанная с цитоскелетом, в которой Ca 2+ -зависимая активация протеаз кальпаина приводит к разрушению цитоскелета [27] и (2) фаза, зависимая от макрофагов, в котором инфильтрация фагоцитарных макрофагов вызывает / коррелирует с ретракцией дистрофических аксонов [28]. Наряду с этим происходит активация ряда ингибирующих белков, включая протеогликаны хондроитинсульфата (CSPG), миелин-ассоциированные ингибиторы (MAI) и хеморепеллентные белки [29].Кроме того, иммунная система реагирует после травмы сопутствующим и устойчивым глиозом, состоящим из активации и активации микроглии, макрофагов и астроцитов. Высвобождение цитокинов и белков ЕСМ, связанных с этим глиозом, может модулировать иммунный ответ и повреждение тканей [29]. Было показано, что активация микроглии после ТСМ способствует устойчивой хронической боли, часто испытываемой людьми после травм [30, 31]. Вместе этот биологический каскад событий не только создает тормозную среду, но также эффективно снижает пластичность аксонов, что приводит к неудачной репарации.
Наряду с этим происходит активация ряда ингибирующих белков, включая протеогликаны хондроитинсульфата (CSPG), миелин-ассоциированные ингибиторы (MAI) и хеморепеллентные белки [29].Кроме того, иммунная система реагирует после травмы сопутствующим и устойчивым глиозом, состоящим из активации и активации микроглии, макрофагов и астроцитов. Высвобождение цитокинов и белков ЕСМ, связанных с этим глиозом, может модулировать иммунный ответ и повреждение тканей [29]. Было показано, что активация микроглии после ТСМ способствует устойчивой хронической боли, часто испытываемой людьми после травм [30, 31]. Вместе этот биологический каскад событий не только создает тормозную среду, но также эффективно снижает пластичность аксонов, что приводит к неудачной репарации.
Большая часть текущих исследований SCI сосредоточена на нейропротекции и / или лечении этих вторичных механизмов. Таким образом, терапевтические стратегии, направленные на удаление или деградацию миелиновых остатков и CSPG, показали большие перспективы для восстановления ЦНС. В этом обзоре мы обсудим некоторые из основных факторов, способствующих предотвращению пластичности, прорастания и регенерации аксонов после травмы спинного мозга.
В этом обзоре мы обсудим некоторые из основных факторов, способствующих предотвращению пластичности, прорастания и регенерации аксонов после травмы спинного мозга.
2. Миелин в нервной системе
Миелинизация — существенная и уникальная особенность нервной системы взрослого человека, обеспечивающая электрическую изоляцию и быстрое распространение потенциалов действия на большие расстояния, а также метаболическую поддержку [32].Олигодендроциты и шванновские клетки представляют собой высокоспециализированные миелинпродуцирующие клетки ЦНС и периферической нервной системы (ПНС) соответственно. Олигодендроциты испускают до 60 проекций из одного тела клетки. Для сравнения, шванновские клетки образуют единую миелиновую междоузлия вокруг одного аксона. В ЦНС каждый выступ олигодендроцита сильно обвивается вокруг короткого аксонального сегмента с образованием миелиновых междоузлий [33]. Это особенно важно для длинных аксонов, которые могут находиться на расстоянии нескольких сантиметров или даже метров от тела клетки. Физиологическая функция и динамика олигодендроцитов и миелина подробно рассмотрены Saab и Nave [34].
Физиологическая функция и динамика олигодендроцитов и миелина подробно рассмотрены Saab и Nave [34].
Миелинизация головного мозга человека продолжается в течение нескольких десятилетий с максимальным объемом белого вещества по сравнению с общим объемом мозга, достигающим пика в пятом десятилетии жизни [35]. Интересно, что центры более высокого порядка, такие как кора головного мозга, миелинизируются позже, чем менее сложные области, участвующие в основных неврологических и гомеостатических функциях, такие как ствол мозга.
Миелин не является статической структурой, а вместо этого медленно и непрерывно модифицируется / ремоделируется с течением времени в нервной системе здорового взрослого человека.В зрительной коре нормальных стареющих макак-резусов длина междоузлий уменьшается, что указывает на то, что активная ремиелинизация происходит на протяжении всей жизни [36]. В самом деле, более короткие миелиновые междоузлия также наблюдаются у здоровых взрослых мышей во время нормального старения [37]. Небольшие изменения, такие как межузловая длина, могут приводить к изменениям скорости проводимости и синхронизации сигнала, что имеет решающее значение для правильного функционирования интегральных нейронных цепей [38, 39]. Подобно тому, как активность нейронов, связанная с когнитивными и моторными задачами, стимулирует синаптическую пластичность, сообщалось, что сходная активность стимулирует усиление регионально-зависимой миелинизации [40].Эти наблюдения подтверждают пластический характер функционального взаимодействия нейроглии.
Небольшие изменения, такие как межузловая длина, могут приводить к изменениям скорости проводимости и синхронизации сигнала, что имеет решающее значение для правильного функционирования интегральных нейронных цепей [38, 39]. Подобно тому, как активность нейронов, связанная с когнитивными и моторными задачами, стимулирует синаптическую пластичность, сообщалось, что сходная активность стимулирует усиление регионально-зависимой миелинизации [40].Эти наблюдения подтверждают пластический характер функционального взаимодействия нейроглии.
Перекрестная связь нейронов (аксонов) и олигодендроцитов (миелин) также необходима для поддержания правильной метаболической функции аксонов. Эта коммуникация модулирует многие факторы, включая локализацию ионных каналов, функцию цитоскелета и аксональный транспорт [41–44], и недавно была тщательно рассмотрена Micu и его коллегами в отношении аксомиелиновой нейротрансмиссии [45]. Миелинизация начинается во время развития, и поэтому миелинизированные аксоны становятся зависимыми от поддержки, обеспечиваемой этими глиальными клетками [46, 47]. Эта зависимость была продемонстрирована на мышах, дефицитных по миелин-специфическому протеолипидному белку (PLP / DM20). Набухание аксонов, связанное преимущественно с миелинизированными аксонами малого диаметра, наблюдалось у этих мышей в возрасте от 6 до 8 недель [46]. Сходным образом, мыши, лишенные 2,3-циклической нуклеотидной фосфодиэстеразы (CNP) в олигодендроцитах, не обнаруживают структурных аномалий в миелине, но развивают набухание аксонов, ведущее к дегенерации во взрослом возрасте (от 3,5 месяцев и старше) [47].
Эта зависимость была продемонстрирована на мышах, дефицитных по миелин-специфическому протеолипидному белку (PLP / DM20). Набухание аксонов, связанное преимущественно с миелинизированными аксонами малого диаметра, наблюдалось у этих мышей в возрасте от 6 до 8 недель [46]. Сходным образом, мыши, лишенные 2,3-циклической нуклеотидной фосфодиэстеразы (CNP) в олигодендроцитах, не обнаруживают структурных аномалий в миелине, но развивают набухание аксонов, ведущее к дегенерации во взрослом возрасте (от 3,5 месяцев и старше) [47].
Миелинизация не только увеличивает скорость проводимости, но также контролирует временное разрешение нейронных цепей.Эта функция имеет решающее значение для точного контроля моторных, сенсорных и когнитивных функций. Таким образом, нейронные цепи способны адаптироваться и реагировать на различные стимулы и события, поддерживая обучение и память. Однако в ЦНС взрослого человека задействованы тормозные механизмы и молекулы, такие как миелин-ассоциированные ингибиторы (MAI), для предотвращения или минимизации общей структурной пластичности и ремоделирования, которые могут быть вредными для физиологической функции цепи. Хотя приоритет этих «защитных» механизмов понятен для сохранения данной сети, он налагает ограничения на пластичность и регенерацию после повреждения ЦНС.Интересно, что в ПНС ситуация несколько иная. Долгое время считалось, что аксоны в ПНС способны регенерироваться после повреждения периферических нервов [48]. В отличие от ЦНС, MAI либо не обнаруживаются, либо быстро выводятся глиальными клетками PNS. Таким образом, как изолированные шванновские клетки, так и трансплантаты периферических нервов, как было показано, создают благоприятную для роста среду после травмы [49–51], подчеркивая фундаментальные различия, лежащие в основе способности ПНС стимулировать рост и восстановление, обеспечивая при этом стимулирующую среду по сравнению с в тормозную среду ЦНС.
Хотя приоритет этих «защитных» механизмов понятен для сохранения данной сети, он налагает ограничения на пластичность и регенерацию после повреждения ЦНС.Интересно, что в ПНС ситуация несколько иная. Долгое время считалось, что аксоны в ПНС способны регенерироваться после повреждения периферических нервов [48]. В отличие от ЦНС, MAI либо не обнаруживаются, либо быстро выводятся глиальными клетками PNS. Таким образом, как изолированные шванновские клетки, так и трансплантаты периферических нервов, как было показано, создают благоприятную для роста среду после травмы [49–51], подчеркивая фундаментальные различия, лежащие в основе способности ПНС стимулировать рост и восстановление, обеспечивая при этом стимулирующую среду по сравнению с в тормозную среду ЦНС.
2.1. Миелин-ассоциированные ингибиторы (MAI)
Nogo, миелиновый гликопротеин олигодендроцитов (OMgp) и миелин-ассоциированный гликопротеин (MAG) были идентифицированы как MAI, которые способны разрушать конусы роста аксонов и ингибировать рост нейритов [52-56] ( Рисунок 1) (Таблица 1). Nogo-A, ассоциированный с мембраной белок, принадлежащий к семейству ретикулонов [57], экспрессируется преимущественно в телах, отростках олигодендроцитов и зрелом миелине. Он также экспрессируется в нескольких типах нейронов в нервной системе развивающихся и взрослых [58].Nogo-A был идентифицирован как ингибитор роста нейритов в 1980-х годах [59, 60]. Это ингибирование было связано по крайней мере с двумя различными доменами, ингибирующими рост, Nogo-66 [61, 62] и Nogo-A-∆20 [63]. Доказательства ингибирующего действия Nogo-A были получены в исследованиях in vitro, в которых было показано, что воздействие Nogo-A на нейроны ганглия сетчатки курицы и ганглия задних корешков крысы (DRG) ингибирует рост нейритов и вызывает коллапс конуса роста [54, 64, 65]. Нейтрализация Nogo-A позволила аксональному росту совместно культивированных сенсорных и симпатических нейронов в эксплантаты зрительного нерва [60] и индуцировала отрастание аксонов Пуркинье взрослых крыс in vivo [66].
Nogo-A, ассоциированный с мембраной белок, принадлежащий к семейству ретикулонов [57], экспрессируется преимущественно в телах, отростках олигодендроцитов и зрелом миелине. Он также экспрессируется в нескольких типах нейронов в нервной системе развивающихся и взрослых [58].Nogo-A был идентифицирован как ингибитор роста нейритов в 1980-х годах [59, 60]. Это ингибирование было связано по крайней мере с двумя различными доменами, ингибирующими рост, Nogo-66 [61, 62] и Nogo-A-∆20 [63]. Доказательства ингибирующего действия Nogo-A были получены в исследованиях in vitro, в которых было показано, что воздействие Nogo-A на нейроны ганглия сетчатки курицы и ганглия задних корешков крысы (DRG) ингибирует рост нейритов и вызывает коллапс конуса роста [54, 64, 65]. Нейтрализация Nogo-A позволила аксональному росту совместно культивированных сенсорных и симпатических нейронов в эксплантаты зрительного нерва [60] и индуцировала отрастание аксонов Пуркинье взрослых крыс in vivo [66].
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
MAI OMgp также экспрессируется в олигодендроцитах и в нескольких типах нейронов ЦНС, таких как пирамидные клетки в гиппокампе и клетки Пуркинье в мозжечке, среди прочих [67]. Хотя о OMgp известно меньше по сравнению с Nogo-A и MAG, как и его аналоги, также было показано, что он является мощным ингибитором роста нейритов во многих клеточных линиях и первичных культурах нейронов [56, 68, 69]. Интересно, что первоначально сообщалось, что OMgp локализуется в узлах Ранвье в ЦНС, а также было показано, что он ингибирует коллатеральное разрастание [69]. У мышей с нокаутом OMgp наблюдались нарушения узлов ЦНС, приводящие к большей длине узлов и большему отрастанию из этих узлов по сравнению с животными дикого типа, указывая тем самым, что включение узлов в оболочку с помощью OMgp важно для развития миелина ЦНС и пластичности аксонов [69].Однако в более недавнем исследовании было показано, что антисыворотка против OMgp, ранее использовавшаяся для определения функции OMgp в узлах, неспецифична [70]. Антисыворотка обнаруживает сильную перекрестную реактивность с CSPG versican V2 и не обнаруживает дефектов в параузлах или повышенного коллатерального разрастания узлов у OMgp-нулевых мышей, потенциально опровергая участие OMgp в узле сборки или поддержания Ranvier [70].
Хотя о OMgp известно меньше по сравнению с Nogo-A и MAG, как и его аналоги, также было показано, что он является мощным ингибитором роста нейритов во многих клеточных линиях и первичных культурах нейронов [56, 68, 69]. Интересно, что первоначально сообщалось, что OMgp локализуется в узлах Ранвье в ЦНС, а также было показано, что он ингибирует коллатеральное разрастание [69]. У мышей с нокаутом OMgp наблюдались нарушения узлов ЦНС, приводящие к большей длине узлов и большему отрастанию из этих узлов по сравнению с животными дикого типа, указывая тем самым, что включение узлов в оболочку с помощью OMgp важно для развития миелина ЦНС и пластичности аксонов [69].Однако в более недавнем исследовании было показано, что антисыворотка против OMgp, ранее использовавшаяся для определения функции OMgp в узлах, неспецифична [70]. Антисыворотка обнаруживает сильную перекрестную реактивность с CSPG versican V2 и не обнаруживает дефектов в параузлах или повышенного коллатерального разрастания узлов у OMgp-нулевых мышей, потенциально опровергая участие OMgp в узле сборки или поддержания Ranvier [70]. Несмотря на эти противоречивые сообщения, важность OMgp в развитии и созревании миелинизации ЦНС была показана у мышей с нокаутом, у которых наблюдалась гипомиелинизация и нарушение скорости проводимости в их спинном мозге [71].
Несмотря на эти противоречивые сообщения, важность OMgp в развитии и созревании миелинизации ЦНС была показана у мышей с нокаутом, у которых наблюдалась гипомиелинизация и нарушение скорости проводимости в их спинном мозге [71].
MAG, другой MAI [52, 72, 73], является второстепенным компонентом зрелого компактного миелина, обогащенного периаксональной мембраной миелиновой оболочки и экспрессируется олигодендроцитами и шванновскими клетками. Первоначально считалось, что МАГ способствует росту нейритов [74]; однако в исследованиях, посвященных его взаимодействию с первичными нейронами, наблюдался противоположный эффект. Таким образом, было обнаружено, что очищенный рекомбинантный МАГ блокирует рост нейритов, а также вызывает сокращение конуса роста [52, 75]. Кроме того, было обнаружено, что клетки яичника китайского хомячка (СНО), экспрессирующие MAG на своей поверхности, совместно культивированные с нейронами мозжечка 7-дневных крыс и DRG взрослых, ингибируют рост нейритов [53].Напротив, когда эмбриональные нейроны DRG выращивали на клетках MAG-CHO, они удлиняли длинные нейриты. Эти результаты подчеркивают онтогенетическую регуляцию роста нейритов и переключение на ингибирование роста, которое происходит во взрослых / постнатальных нейронах [53]. Ингибирующие свойства MAG были дополнительно подтверждены демонстрацией того, что миелин мышей с нокаутом MAG не подавлял рост нейронов DRG in vitro по сравнению с миелином нормальных мышей [76]. Более того, подавление роста нейритов было полностью отменено иммунным истощением MAG из растворимой фракции кондиционированной миелином среды [77].Эти наблюдения предполагают, что в дополнение к остаткам миелина растворимые MAI, вероятно, высвобождаемые после повреждения, могут влиять на способность нейронов и аксонов к росту.
Эти результаты подчеркивают онтогенетическую регуляцию роста нейритов и переключение на ингибирование роста, которое происходит во взрослых / постнатальных нейронах [53]. Ингибирующие свойства MAG были дополнительно подтверждены демонстрацией того, что миелин мышей с нокаутом MAG не подавлял рост нейронов DRG in vitro по сравнению с миелином нормальных мышей [76]. Более того, подавление роста нейритов было полностью отменено иммунным истощением MAG из растворимой фракции кондиционированной миелином среды [77].Эти наблюдения предполагают, что в дополнение к остаткам миелина растворимые MAI, вероятно, высвобождаемые после повреждения, могут влиять на способность нейронов и аксонов к росту.
2.2. Миелин-ассоциированная пластичность
Помимо ингибирующей роли MAI в росте нейритов, эти молекулы имеют значительное влияние на контроль пластичности в ЦНС. В развивающейся нервной системе экспрессия Nogo-A высока во многих нейронах. Таким образом, Nogo-A, как было показано, быстро модулирует динамику актинового цитоскелета, чтобы контролировать структурную пластичность дендритных шипов пирамидных нейронов CA3 в гиппокампе in vivo [78]. Также было показано, что Nogo-A ограничивает вставку рецептора α -амино-3-гидрокси-5-метил-4-изоксазолепропионовой кислоты (AMPA), обеспечивая механизм, с помощью которого он может регулировать силу возбуждающей синаптической передачи [78]. В мозжечке экспрессия Nogo-A коррелирует с утончением и развитием синапсов, а также, как было показано, негативно регулирует морфологию дендритов и синаптическую передачу [79]. Также было показано, что нейронный Nogo-A в гиппокампе взрослых ограничивает физиологическую синаптическую активность.Это было продемонстрировано посредством функционального ингибирования антителами Nogo-A или его рецептора, рецептора 1 Nogo-66 (NgR1), а также у мышей с нокаутом Nogo-A [80]. Нейтрализация Nogo-A и NgR1 приводила к увеличению долговременной потенциации (LTP) без изменения долговременной депрессии (LTD) в острых срезах гиппокампа взрослых мышей [80]. Экспрессия NgR1 необходима для подавления синаптического обмена в нервной системе взрослых и для поддержания и стабилизации дендритных шипов и варикозного расширения аксонов в ответ на Nogo-A и другие лиганды.
Также было показано, что Nogo-A ограничивает вставку рецептора α -амино-3-гидрокси-5-метил-4-изоксазолепропионовой кислоты (AMPA), обеспечивая механизм, с помощью которого он может регулировать силу возбуждающей синаптической передачи [78]. В мозжечке экспрессия Nogo-A коррелирует с утончением и развитием синапсов, а также, как было показано, негативно регулирует морфологию дендритов и синаптическую передачу [79]. Также было показано, что нейронный Nogo-A в гиппокампе взрослых ограничивает физиологическую синаптическую активность.Это было продемонстрировано посредством функционального ингибирования антителами Nogo-A или его рецептора, рецептора 1 Nogo-66 (NgR1), а также у мышей с нокаутом Nogo-A [80]. Нейтрализация Nogo-A и NgR1 приводила к увеличению долговременной потенциации (LTP) без изменения долговременной депрессии (LTD) в острых срезах гиппокампа взрослых мышей [80]. Экспрессия NgR1 необходима для подавления синаптического обмена в нервной системе взрослых и для поддержания и стабилизации дендритных шипов и варикозного расширения аксонов в ответ на Nogo-A и другие лиганды. Покадровая визуализация in vivo у мышей с нокаутом NgR1 выявила усиление динамики позвоночника без изменения общей плотности позвоночника в соматосенсорной коре [81].
Покадровая визуализация in vivo у мышей с нокаутом NgR1 выявила усиление динамики позвоночника без изменения общей плотности позвоночника в соматосенсорной коре [81].
Генетическая абляция NgR1, как было показано, увеличивает синаптическую плотность в ювенильном гиппокампе, а также увеличивает плотность шипов в первичных нейронах гиппокампа [82]. Более того, в более раннем исследовании с использованием животных с нокаутом NgR1, NMDA-рецептор-зависимая LTD ослаблялась в коллатеральной цепи Шаффера (CA3-CA1) гиппокампа [83].Напротив, в срезах гиппокампа взрослых мышей обработка растворимым Nogo-A (Nogo66) или OMgp подавляла LTP NgR1-зависимым образом [84]. Обработка антителами против Nogo-A или NgR1 срезов коронарного мозга взрослых крыс, содержащих первичную моторную кору, увеличивала LTP через NgR1 [85]. Применение антитела против Nogo-A in vivo привело к большей плотности дендритных шипов и улучшенной двигательной способности при достижении передних конечностей по сравнению с контрольными животными, получавшими антитело [85]. Важно отметить, что лечение антителами против NogoA также продемонстрировало, что в здоровой системе ингибирование пути Nogo вызывает разрастание волокон CA3 [86].В исследовании Zemmar et al., Линии мышей, у которых Nogo-A был нокаутирован либо в олигодендроцитах, либо в нейронах, выявили усиленное ветвление дендритов и образование шипов, подтверждая, что оба источника Nogo-A вносят вклад в развитие синапсов и пластичность [87].
Важно отметить, что лечение антителами против NogoA также продемонстрировало, что в здоровой системе ингибирование пути Nogo вызывает разрастание волокон CA3 [86].В исследовании Zemmar et al., Линии мышей, у которых Nogo-A был нокаутирован либо в олигодендроцитах, либо в нейронах, выявили усиленное ветвление дендритов и образование шипов, подтверждая, что оба источника Nogo-A вносят вклад в развитие синапсов и пластичность [87].
Недавно был открыт рецептор, связанный с G-белком для Nogo-A-Δ20, сфинголипид-1-фосфатный рецептор 2 (S1PR2). Было показано, что Nogo-A-Δ20 связывает S1PR2, чтобы опосредовать его эффекты ингибирования роста путем передачи сигналов через G-белок G 13 , лейкоз-ассоциированный фактор обмена Rhoguanine (LARG) и RhoA, чтобы опосредовать дестабилизацию актина и коллапс конуса роста [ 88].Кроме того, подобно стратегиям нейтрализации Nogo-A с использованием антител или парадигм нокдауна, ингибирование S1PR2 блокировало Nogo-A-Δ20, тем самым противодействуя ингибированию Nogo-A-Δ20 роста нейритов и распространения клеток. Ингибирование S1PR2 также увеличивает гиппокампальный и кортикальный LTP, таким образом модулируя синаптическую пластичность [88]. Эти результаты демонстрируют, что MAI и их рецепторы способны противодействовать путям, способствующим росту, чтобы модулировать синаптическую силу и передачу, влияя на синаптическую пластичность.
Ингибирование S1PR2 также увеличивает гиппокампальный и кортикальный LTP, таким образом модулируя синаптическую пластичность [88]. Эти результаты демонстрируют, что MAI и их рецепторы способны противодействовать путям, способствующим росту, чтобы модулировать синаптическую силу и передачу, влияя на синаптическую пластичность.
Nogo-A и NgR1, как было показано, также важны для регуляции формирования памяти и регулирования выполнения различных поведенческих задач [81, 85, 89, 90]. Напр., Karlsson и его коллеги продемонстрировали нарушение опорно-двигательной функции и распознавание новых объектов у мышей, лишенных NgR1 [90]. Избыточная экспрессия NgR1 нарушает пространственную обучающую память с уменьшением плотности и сложности зрелого позвоночника [90]. Кроме того, тест пространственного обучения в водном лабиринте Морриса также показал, что мыши с нокаутом Nogo-A могли находить скрытую платформу быстрее, чем животные дикого типа, и обладали лучшим долгосрочным сохранением памяти. Это улучшение успеваемости коррелирует с увеличением плотности дендритных шипов в апикальных дендритах CA3 [91].
Это улучшение успеваемости коррелирует с увеличением плотности дендритных шипов в апикальных дендритах CA3 [91].
Участие MAIs в ограничении пластичности нейронов также элегантно продемонстрировано в зрительной системе грызунов. В мозге подростка наступают критические периоды (ЦП) пластичности, основанной на опыте. Это важно для нормального развития первичной зрительной коры. Таким образом, визуальные переживания во время КП улучшают нейронную сеть зрительной системы, обеспечивая формирование столбцов окулярного доминирования в коре головного мозга.В конце CP зрелые сети, которые были сформированы, активно поддерживаются, тем самым ограничивая пластичность [92]. У NgR1 и Nogo-A мутантных мышей, хотя пластичность происходит во время CP, также было показано, что она продолжается за пределами нормальной длины CP [93, 94]. Как следствие, стойкая пластичность окулярного доминирования, связанная с монокулярной депривацией, сохраняется на протяжении всей взрослой жизни [93, 94]. Сходным образом пластичность окулярного доминирования увеличивалась при нокауте у мышей рецептора MAI, известного как парный иммуноглобулиноподобный рецептор B (PirB) [95]. Эти исследования демонстрируют, что MAI и их рецепторы стабилизируют нейронную цепь и ограничивают пластичность нейронов, обусловленную опытом. Таким образом, MAI, как и многие другие белки, участвуют в консолидации и закрытии CP посредством стабилизации зрелых, высоконастроенных и сложных нейронных сетей.
Эти исследования демонстрируют, что MAI и их рецепторы стабилизируют нейронную цепь и ограничивают пластичность нейронов, обусловленную опытом. Таким образом, MAI, как и многие другие белки, участвуют в консолидации и закрытии CP посредством стабилизации зрелых, высоконастроенных и сложных нейронных сетей.
2.3. Борьба с ингибированием миелина для ускорения роста нейритов и регенерации аксонов
Как обсуждалось, миелин содержит несколько молекул, ингибирующих рост, которые ограничивают как пластичность, так и регенерацию аксонов в поврежденной ЦНС млекопитающих (таблица 1), некоторые из которых были подробно рассмотрены в других работах [34, 96, 97].Ряд стратегий противодействия и нейтрализации эффектов MAI, начиная с полной демиелинизации, показали улучшения в регенерации аксонов и функциональном восстановлении. Исследования Кейрстеда и его коллег в 1990-х годах показали, что после полной или частичной демиелинизации в месте поражения и проксимально от него происходит регенерация аксонов с обширным разрастанием нейритов [98, 99]. В частности, после латеральной гемисекции спинного мозга взрослой крысы было показано, что временное разрушение зрелого миелина за счет внутриоспинального слияния белков комплемента сыворотки и миелин-специфических антител (антигалактоцереброзид, главный сфинголипид в миелине) способствует регенерации аксонов руброспинальной аксоны в хвостовой отдел поясничного отдела спинного мозга [98].Используя аналогичный антимиелиновый подход, Кейрстед и его коллеги продемонстрировали, что демиелинизация, вызванная антигалактоцереброзидом и комплементом, в сочетании с трансплантацией шванновских клеток усиливает регенерацию аксонов после травмы [99]. Аналогичным образом, в исследованиях, направленных на блокирование активности Nogo-A с помощью антител против Nogo-A, было значительно увеличено регенерация и прорастание аксонов перерезанного кортикоспинального тракта (CST) у крыс после поражения спинного мозга по сравнению с контрольной группой [100, 101]. Кроме того, лечение анти-Nogo-A способствовало прорастанию компенсаторных волокон и иннервации от неповрежденной стороны к денервированной стороне [100].
В частности, после латеральной гемисекции спинного мозга взрослой крысы было показано, что временное разрушение зрелого миелина за счет внутриоспинального слияния белков комплемента сыворотки и миелин-специфических антител (антигалактоцереброзид, главный сфинголипид в миелине) способствует регенерации аксонов руброспинальной аксоны в хвостовой отдел поясничного отдела спинного мозга [98].Используя аналогичный антимиелиновый подход, Кейрстед и его коллеги продемонстрировали, что демиелинизация, вызванная антигалактоцереброзидом и комплементом, в сочетании с трансплантацией шванновских клеток усиливает регенерацию аксонов после травмы [99]. Аналогичным образом, в исследованиях, направленных на блокирование активности Nogo-A с помощью антител против Nogo-A, было значительно увеличено регенерация и прорастание аксонов перерезанного кортикоспинального тракта (CST) у крыс после поражения спинного мозга по сравнению с контрольной группой [100, 101]. Кроме того, лечение анти-Nogo-A способствовало прорастанию компенсаторных волокон и иннервации от неповрежденной стороны к денервированной стороне [100].
В исследованиях, направленных на рецепторы MAI, было проведено несколько многообещающих исследований по усилению разрастания нейритов и регенерации аксонов при модуляции этих рецепторов. В рецепторе NgR1 отсутствует трансмембранный домен и, следовательно, он образует комплекс с другими трансмембранными корецепторами, такими как LINGO-1 (LRR и Ig-содержащий белок, взаимодействующий с рецептором Nogo) и p75NTR (рецептор нейтротрофина p75) или TROY (суперсемейство рецепторов фактора некроза опухоли). член). При этом он может передавать сигнал ингибирования миелина аксону, таким образом опосредуя коллапс конуса роста и остановку роста нейритов [102].Примечательно, что все три MAI связываются с одним и тем же рецепторным комплексом NgR1 и активируют его [61, 68, 103]. Все три MAI также связываются с рецептором PirB [103]. Блокирование рецептора NgR1 или PirB частично ослабляет коллапс конуса роста и разрастание нейритов в DRG и нейронах гранул мозжечка в присутствии MAI или миелина. Однако в NgR1 нулевых нейронах мозжечка опосредованное антителами ингибирование PirB подавляло ингибирующие эффекты MAIs на рост нейритов [103]. Сообщается, что in vivo PirB играет ключевую роль в пластичности коры [95].В отличие от наблюдений in vitro, регенерация CST не усиливалась у PirB-нокаутных мышей после SCI [104]. Ряд других исследований генетических делеций / мутаций MAI и их рецепторов предоставили дополнительные доказательства ингибирующего действия этих белков на рост. Однако эффекты после повреждения у мышей с нокаутом по NgR1 или Nogo-A, MAG и OMgp были разными [105–107]. У мышей с тройными мутантами (Nogo-A, MAG и OMgp) больший рост аксонов и отрастание аксонов CST и швовидного тракта и улучшенная локомоция наблюдались после SCI по сравнению с одиночным или двойным нокаутом [108].Однако Ли и его коллеги наблюдали ограниченное прорастание аксонов и регенерацию аксонов CST после SCI без синергетических эффектов у животных с тройным нокаутом [107]. Эта вариабельность, вероятно, может быть связана с локализацией и типом повреждения, а также с компенсацией и синергизмом между MAI и другими факторами, ингибирующими рост, у животных с единичным нокаутом.
Однако в NgR1 нулевых нейронах мозжечка опосредованное антителами ингибирование PirB подавляло ингибирующие эффекты MAIs на рост нейритов [103]. Сообщается, что in vivo PirB играет ключевую роль в пластичности коры [95].В отличие от наблюдений in vitro, регенерация CST не усиливалась у PirB-нокаутных мышей после SCI [104]. Ряд других исследований генетических делеций / мутаций MAI и их рецепторов предоставили дополнительные доказательства ингибирующего действия этих белков на рост. Однако эффекты после повреждения у мышей с нокаутом по NgR1 или Nogo-A, MAG и OMgp были разными [105–107]. У мышей с тройными мутантами (Nogo-A, MAG и OMgp) больший рост аксонов и отрастание аксонов CST и швовидного тракта и улучшенная локомоция наблюдались после SCI по сравнению с одиночным или двойным нокаутом [108].Однако Ли и его коллеги наблюдали ограниченное прорастание аксонов и регенерацию аксонов CST после SCI без синергетических эффектов у животных с тройным нокаутом [107]. Эта вариабельность, вероятно, может быть связана с локализацией и типом повреждения, а также с компенсацией и синергизмом между MAI и другими факторами, ингибирующими рост, у животных с единичным нокаутом. Действительно, ингибирующие рост молекулы, такие как EphrinA3, передающие сигнал через рецептор EphrinA4, активируются в спинном мозге мышей с нокаутом Nogo-A, что позволяет предположить, что компенсаторные механизмы опосредуют остаточное ингибирование, наблюдаемое у этих животных, чтобы ограничить рост и восстановление нейритов [109 ].
Действительно, ингибирующие рост молекулы, такие как EphrinA3, передающие сигнал через рецептор EphrinA4, активируются в спинном мозге мышей с нокаутом Nogo-A, что позволяет предположить, что компенсаторные механизмы опосредуют остаточное ингибирование, наблюдаемое у этих животных, чтобы ограничить рост и восстановление нейритов [109 ].
Борьба с ингибированием миелина с помощью других стратегий лечения, включая вакцины, моноклональные антитела (включая антитело против Nogo A) и пептидные лиганды, которые конкурируют за связывание с рецептором [62, 110], показали значительный потенциал восстановления ЦНС [111]. После острой или хронической ТСМ у грызунов лечение растворимым белком-приманкой NgR1 (NgR1 (310) ecto-Fc), который нейтрализует эффекты всех трех MAI (Nogo-A, MAG и OMgp), дало многообещающие результаты в стимулировании аксональной прорастание и функциональное восстановление [112–114].Интересно, что после травмы спинного мозга у взрослых крыс включение либо прекондиционирующего поражения периферического седалищного нерва и применение лечения белком-приманкой NgR1, либо лечение хондроитиназой ABC (ChABC — фермент, который переваривает гликозаминогликановые сахарные цепи CSPG) привело к большему увеличению регенерация аксонов [115]. Комбинация двух из этих процедур больше не усиливала регенерацию; однако объединение всех трех продвинутых аксонов для регенерации в миллиметрах от места поражения [115].
Комбинация двух из этих процедур больше не усиливала регенерацию; однако объединение всех трех продвинутых аксонов для регенерации в миллиметрах от места поражения [115].
Было показано, что острая блокада антител только к Nogo-A усиливает разрастание аксонов в поврежденном спинном мозге, а также улучшает поведенческие исходы [100, 101, 116]. В модели инсульта на крысах лечение антителами против Nogo-A увеличивало ветвление дендритов и плотность шипов [117]. Подобные методы лечения также привели к увеличению прорастания и регенерации волокон после повреждения в органотипических культурах срезов гиппокампа крыс [86]. Антитела против Nogo-A, доставляемые посредством инъекции гибридомных клеток, продуцирующих антитело IN-1 (повышенное против ингибирующей фракции миелина), в лобно-теменную кору за 7-10 дней до поражения спинного мозга, способствовали регенерации аксонов размером до 7-11 мм в перерезанные аксоны CST в течение 2–3 недель после травмы [116].В дополнительном исследовании у взрослых крыс, которым имплантировали гибридомы, секретирующие IN-1, в гиппокамп сразу после дорсальной гипергемисекции спинного мозга, наблюдалось большее улучшение опорно-двигательной функции [101]. Исследование, в котором использовалось интратекальное введение антител против Nogo-A в субдуральное пространство, также индуцировало регенерацию аксонов CST наряду с улучшением поведения при передвижении в открытом поле, ходьбе по лестнице, ходьбе с узким лучом, а также улучшением реакции на тепло и появление волосков фон Фрея [ 100].Благодаря успешной регенерации, наблюдаемой на экспериментальных моделях травм, обработанных антителами против Nogo-A, как описано выше, в настоящее время проводятся клинические испытания с использованием этих антител для лечения инсульта и SCI [111, 118].
Исследование, в котором использовалось интратекальное введение антител против Nogo-A в субдуральное пространство, также индуцировало регенерацию аксонов CST наряду с улучшением поведения при передвижении в открытом поле, ходьбе по лестнице, ходьбе с узким лучом, а также улучшением реакции на тепло и появление волосков фон Фрея [ 100].Благодаря успешной регенерации, наблюдаемой на экспериментальных моделях травм, обработанных антителами против Nogo-A, как описано выше, в настоящее время проводятся клинические испытания с использованием этих антител для лечения инсульта и SCI [111, 118].
2.4. Ремиелинизация после травмы
Несмотря на множество тормозящих факторов, связанных с миелином, замещение олигодендроцитов и миелина вокруг демиелинизированных аксонов считается ключевым механизмом, способствующим регенерации ЦНС с целью восстановления перекрестного взаимодействия нейронов и глии и функционального восстановления.Зрелые олигодендроциты, как и нейроны, являются постмитотическими клетками и поэтому не участвуют в ремиелинизации после повреждения. Ремиелинизация устанавливается преимущественно пролиферирующими клетками-предшественниками олигодендроцитов (OPC), экспрессирующими рецептор тромбоцитарного фактора роста A, клетки нейрон-глиального антигена 2 (NG2), а также нервные стволовые клетки и клетки-предшественники (NSPC). Эти клетки мигрируют в место поражения, где они дифференцируются и дают зародыши, миелинизирующие олигодендроциты после повреждения [119–122].Этот устойчивый олигодендрогенез происходит в течение первых нескольких месяцев после травмы, что частично компенсирует потерю зрелых олигодендроцитов и миелина [123]. Было показано, что вновь образованные олигодендроциты генерируют миелиновые оболочки за короткий промежуток времени [124]. Таким образом, возможны пластические изменения, связанные с де- и ремиелинизацией. В самом деле, генетическое удаление новых миелин-образующих олигодендроцитов ухудшает выполнение сложной моторной задачи у мышей [125].
Ремиелинизация устанавливается преимущественно пролиферирующими клетками-предшественниками олигодендроцитов (OPC), экспрессирующими рецептор тромбоцитарного фактора роста A, клетки нейрон-глиального антигена 2 (NG2), а также нервные стволовые клетки и клетки-предшественники (NSPC). Эти клетки мигрируют в место поражения, где они дифференцируются и дают зародыши, миелинизирующие олигодендроциты после повреждения [119–122].Этот устойчивый олигодендрогенез происходит в течение первых нескольких месяцев после травмы, что частично компенсирует потерю зрелых олигодендроцитов и миелина [123]. Было показано, что вновь образованные олигодендроциты генерируют миелиновые оболочки за короткий промежуток времени [124]. Таким образом, возможны пластические изменения, связанные с де- и ремиелинизацией. В самом деле, генетическое удаление новых миелин-образующих олигодендроцитов ухудшает выполнение сложной моторной задачи у мышей [125].
MAI и другие ограничивающие рост белки важны для поддержания физиологической пластичности и стабильности нейронной сети.![]() К сожалению, это эффективно ограничивает / предотвращает восстановление функциональной связи выживших нейронов / сетей после травмы или заболевания. Важность миелинизации для функции ЦНС была описана ранее, и поэтому понятно, что после травмы, когда происходит дегенерация аксонов и демиелинизация, также происходит спонтанная ремиелинизация для стабилизации уязвимых аксонов [126, 127]. Однако, подобно неадекватной регенерации аксонов, которая происходит после травмы, степень и качество ремиелинизации также считаются недостаточными, что приводит к неполной, аномально тонкой миелиновой оболочке и продолжающейся дисфункции аксонов [128, 129].
К сожалению, это эффективно ограничивает / предотвращает восстановление функциональной связи выживших нейронов / сетей после травмы или заболевания. Важность миелинизации для функции ЦНС была описана ранее, и поэтому понятно, что после травмы, когда происходит дегенерация аксонов и демиелинизация, также происходит спонтанная ремиелинизация для стабилизации уязвимых аксонов [126, 127]. Однако, подобно неадекватной регенерации аксонов, которая происходит после травмы, степень и качество ремиелинизации также считаются недостаточными, что приводит к неполной, аномально тонкой миелиновой оболочке и продолжающейся дисфункции аксонов [128, 129].
Во многих исследованиях сообщается о высокой распространенности очень тонких миелиновых зон, окружающих зоны поражения, даже через годы после ТСМ. У взрослых крыс неполная миелинизация наблюдалась в течение 450 дней после контузивной травмы спинного мозга [130]. При посмертном хроническом повреждении спинного мозга человека через 1-2 года после травмы иммуногистохимический анализ также продемонстрировал некоторую демиелинизацию аксонов даже через десять лет после травмы [131].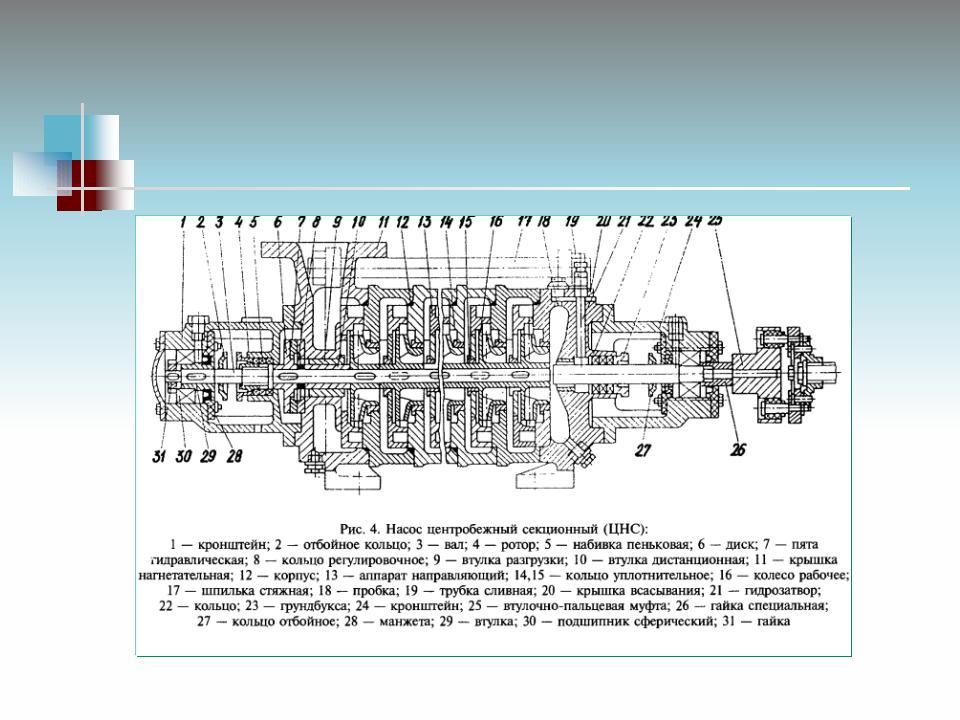
Экспериментально после хронической контузивной травмы спинного мозга большинство сохранившихся руброспинальных аксонов демиелинизируются у крыс [132] и мышей [133].Путем измерения сохраненных руброспинальных аксонов, которые пересекают место повреждения с укороченными межузловыми промежутками (что свидетельствует о ремиелинизации), было обнаружено, что большинство сохраненных аксонов ремиелинизировались через 3 месяца после SCI [132]. Hesp и его коллеги также продемонстрировали ремиелинизацию сохраненных аксонов через 2–3 месяца после контузивной травмы спинного мозга, хотя стереотипно она короче и тоньше, что позволяет предположить, что ремиелинизация протекает постоянно и хронически после травмы спинного мозга [123]. В последующем исследовании Пауэрс и его коллеги измерили спонтанно регенерированный миелин in vivo [120].В соответствии с предыдущими исследованиями они обнаружили, что вскоре после травмы средняя длина регенерированных миелиновых оболочек была значительно короче, чем у неповрежденного миелина; однако через 6 месяцев после травмы вновь сформированные миелиновые оболочки и междоузлия были аналогичны контрольным [120].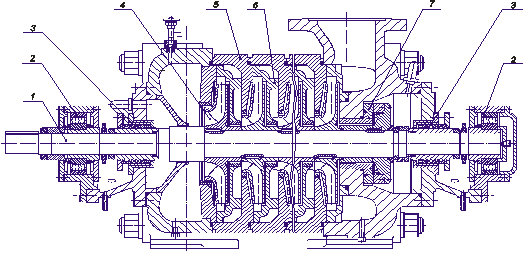 Эти исследования показывают, что аномально тонкие и короткие миелиновые междоузлия, наблюдаемые через много месяцев после травмы / патологии, могут быть связаны с продолжающейся демиелинизацией, а не с неполной регенерацией миелина в интактных сохранившихся аксонах.
Эти исследования показывают, что аномально тонкие и короткие миелиновые междоузлия, наблюдаемые через много месяцев после травмы / патологии, могут быть связаны с продолжающейся демиелинизацией, а не с неполной регенерацией миелина в интактных сохранившихся аксонах.
Спонтанная ремиелинизация после ТСМ протекает медленнее и менее эффективно, чем ремиелинизация после химически индуцированной очаговой демиелинизации.У мышей после фокальной демиелинизации, опосредованной лизолецитином, остатки миелина быстро очищаются в течение первых 4 дней [134], ремиелинизация наблюдается через 7 дней и завершается через 3 недели [135]. Скорость ремиелинизации после SCI, вероятно, обусловлена ингибирующей средой и возникающей в результате патологией, включая миелиновый дебрис и ассоциированные MAI (NogoA, MAG и OMgp), которые сами по себе ингибируют дифференцировку OPC [136, 137]. Ясно, что взаимодействие демиелинизации и компенсаторной ремиелинизации после травмы важно для функционального восстановления. Несколько источников миелинизирующих клеток были исследованы как основанные на трансплантации методы лечения ТСМ [4, 138, 139]. Такое лечение привело к улучшению восстановления поведения после ушиба ТСМ [140].
Несколько источников миелинизирующих клеток были исследованы как основанные на трансплантации методы лечения ТСМ [4, 138, 139]. Такое лечение привело к улучшению восстановления поведения после ушиба ТСМ [140].
Подобно OPC, нервные клетки-предшественники также активируются и мигрируют к месту повреждения. Однако они преимущественно дифференцируются в реактивные астроциты [121, 122, 129]. Эта миграция клеток вносит вклад в тормозную среду, продуцируемую MAI в глиальном рубце и вокруг него после SCI, тем самым препятствуя регенерации аксонов [141].В дополнение к нацеливанию на MAI в качестве барьера для нейрорегенерации после повреждения, другие молекулы, такие как компоненты ECM, которые работают синергетически для поддержания стабильной, преимущественно ингибирующей рост среды, также должны быть рассмотрены и обсуждаются в следующих разделах.
3. CSPG и перинейрональные сети
ECM ЦНС богат на CSPG, некоторые из которых существуют во внеклеточной среде, а другие связаны со специфическими структурами.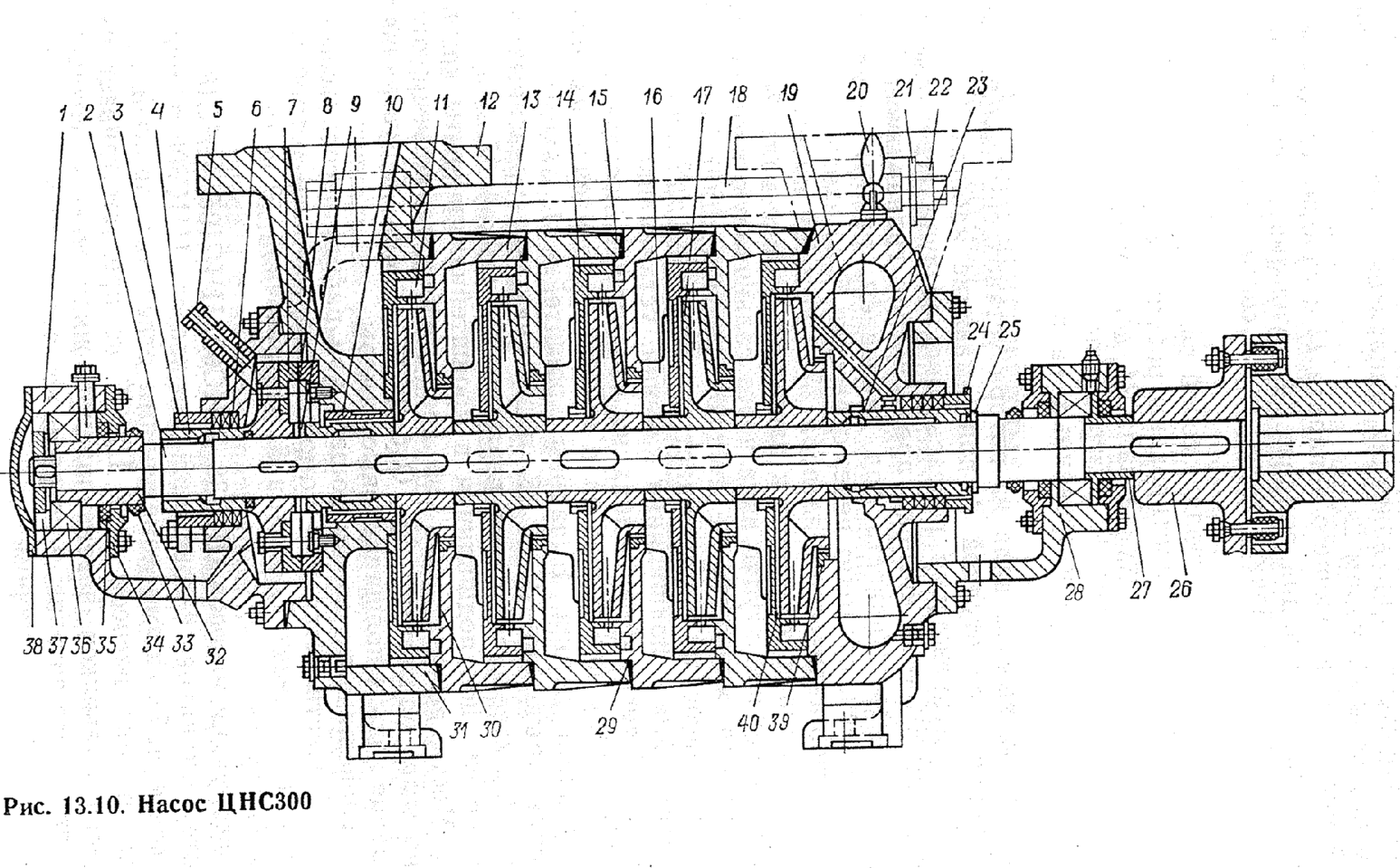 Внутри ЦНС CSPG могут ассоциироваться в специализированные структуры, называемые перинейрональными сетями (PNN), которые окружают сомы и дендриты зрелых нейронов.PNN состоят из белков ECM, включая гиалуронан, связывающих белков [например, связывающего белка 1 хряща (Crtl1 / Hapln1) и специфического для мозга связывающего белка (Bral2 / Hapln4)], CSPG и тенасцина-R (TN-R) [142] , с одним дополнительным исследованием, также идентифицирующим тенасцин-C (TN-C) с помощью иммуноблоттинга в PNNs [143] (Рисунок 1) (Таблица 1). Существует также ряд CSPG, включая бревикан, нейрокан, аггрекан, фосфакан и версикан, которые связываются с гиалуроновой основой PNN (Таблица 1) [144]. Гиалуронан синтезируется гиалуронансинтазами (HAS).Его взаимодействие с CSPG происходит через связывающие белки (например, Hapln1). Молекулы TN-R действуют для перекрестного связывания CSPG с образованием организованного остова PNN. Кроме того, TN-R, как было показано, взаимодействует с интегринами, такими как β 1 интегрины, contactin / F3 / F11, фосфакан и MAG [145–147].
Внутри ЦНС CSPG могут ассоциироваться в специализированные структуры, называемые перинейрональными сетями (PNN), которые окружают сомы и дендриты зрелых нейронов.PNN состоят из белков ECM, включая гиалуронан, связывающих белков [например, связывающего белка 1 хряща (Crtl1 / Hapln1) и специфического для мозга связывающего белка (Bral2 / Hapln4)], CSPG и тенасцина-R (TN-R) [142] , с одним дополнительным исследованием, также идентифицирующим тенасцин-C (TN-C) с помощью иммуноблоттинга в PNNs [143] (Рисунок 1) (Таблица 1). Существует также ряд CSPG, включая бревикан, нейрокан, аггрекан, фосфакан и версикан, которые связываются с гиалуроновой основой PNN (Таблица 1) [144]. Гиалуронан синтезируется гиалуронансинтазами (HAS).Его взаимодействие с CSPG происходит через связывающие белки (например, Hapln1). Молекулы TN-R действуют для перекрестного связывания CSPG с образованием организованного остова PNN. Кроме того, TN-R, как было показано, взаимодействует с интегринами, такими как β 1 интегрины, contactin / F3 / F11, фосфакан и MAG [145–147]. Недавно было обнаружено, что семафорин 3A (sema3A) и 3B (sema3B) (обсуждаемые ниже) также связаны с CSPGs в структуре PNN [148].
Недавно было обнаружено, что семафорин 3A (sema3A) и 3B (sema3B) (обсуждаемые ниже) также связаны с CSPGs в структуре PNN [148].
Хотя PNN локализованы в некоторых глутаматергических нейронах в ЦНС [149], они преимущественно связаны с телом нейронных клеток и проксимальными дендритами парвальбумин-положительных ГАМКергических интернейронов, образуя прочную решетчатую структуру [144, 150 ].Поддержание этой специализированной структуры важно для стабилизации синапсов и сетей и гомеостаза. В частности, PNN функционируют для стабилизации зрелых нейронов за счет снижения пластичности дендритных шипов [151], формирования каркаса для синаптических ингибирующих молекул [152], а также ограничения движения рецепторов в синапсе [153]. Формирование и созревание PNN происходит одновременно с развитием и созреванием нервной системы во время ХП. Ферментативное переваривание этой специализированной структуры ECM с ChABC продлевает CP в зрительной системе [154].Когда удаляются определенные компоненты PNN, такие как линк белок 1 (Crtl1), образование PNN снижается, а пластичность нейронов повышается [155]. Более того, первичные нейрональные и астроцитарные культуры от мышей с четырехкратным нокаутом, у которых отсутствуют нейрокан CSPG, бревикан, а также TN-C и TN-R, снижают образование PNN. В этих случаях у мышей обнаруживаются изменения синаптической пластичности, нарушение синаптогенеза, стабильность синапсов и измененная синаптическая активность [156].
Более того, первичные нейрональные и астроцитарные культуры от мышей с четырехкратным нокаутом, у которых отсутствуют нейрокан CSPG, бревикан, а также TN-C и TN-R, снижают образование PNN. В этих случаях у мышей обнаруживаются изменения синаптической пластичности, нарушение синаптогенеза, стабильность синапсов и измененная синаптическая активность [156].
3.1. Глиальный рубец и связанные с ним ингибирующие белки
Как обсуждалось выше, в отличие от ПНС, ЦНС взрослых млекопитающих имеет пониженную способность к регенерации.Частично это является результатом образования характерного глиального рубца в месте повреждения, который окружает поражение реактивными астроцитами и фибробластами и тормозит регенерацию аксонов [29]. Более того, Дэвис и его коллеги показали, что когда глиальные рубцы сведены к минимуму, сильная регенерация ЦНС может происходить даже в областях дегенерированного белого вещества в спинном мозге [157, 158].
CSPG представляют собой основное семейство ингибирующих белков, связанных с глиальным рубцом и повреждением аксонов [8–10]. К ним относятся подгруппа лектиканов (включающая аггрекан, бревикан, версикан и нейрокан), малые богатые лейцином протеогликаны (включая бигликан и декорин), фосфакан / белок-тирозинфосфатаза рецепторного типа β (RPTP β ) и другие протеогликаны, включая NG2 и нейрогликан-C [159]. После повреждения CSPG активно секретируются в ECM, в основном реактивными астроцитами и нейронами [160], но с незначительным компонентом, также поступающим из секреции макрофагами и олигодендроцитами [145, 161, 162].Это приводит к обилию CSPG в месте повреждения, добавляя к ингибирующей среде. Основной особенностью, способствующей ингибирующей природе CSPG, является их структура, поскольку они имеют ряд боковых цепей гликозаминогликанов (GAG). Каждый CSPG различается по количеству имеющихся у них боковых цепей; например, аггрекан содержит более 100 боковых цепей ГАГ, в то время как другие содержат менее 10 [159]. Количество цепей и статус сульфатирования CSPG соответствуют функциональному влиянию CSPG на рост ЦНС [10, 163–165].
К ним относятся подгруппа лектиканов (включающая аггрекан, бревикан, версикан и нейрокан), малые богатые лейцином протеогликаны (включая бигликан и декорин), фосфакан / белок-тирозинфосфатаза рецепторного типа β (RPTP β ) и другие протеогликаны, включая NG2 и нейрогликан-C [159]. После повреждения CSPG активно секретируются в ECM, в основном реактивными астроцитами и нейронами [160], но с незначительным компонентом, также поступающим из секреции макрофагами и олигодендроцитами [145, 161, 162].Это приводит к обилию CSPG в месте повреждения, добавляя к ингибирующей среде. Основной особенностью, способствующей ингибирующей природе CSPG, является их структура, поскольку они имеют ряд боковых цепей гликозаминогликанов (GAG). Каждый CSPG различается по количеству имеющихся у них боковых цепей; например, аггрекан содержит более 100 боковых цепей ГАГ, в то время как другие содержат менее 10 [159]. Количество цепей и статус сульфатирования CSPG соответствуют функциональному влиянию CSPG на рост ЦНС [10, 163–165].
CSPG оказывают ингибирующее действие как на олигодендроциты, так и на нейроны. Было показано, что это опосредуется сигма-рецептором протеинтирозинфосфатазы (PTP σ ). Когда CSPG связываются с рецепторами PTP σ , они инициируют активацию сигнального пути GTPase Rho / ROCK. В нейронах это приводит к ингибированию роста аксонов, переводя ростовые конусы в дистрофическое состояние [13-15]. Используя новый пептидный миметик PTP σ , Ланг и его коллеги смогли заблокировать опосредованное CSPG ингибирование in vitro, что позволило взрослым сенсорным нейронам распространять аксоны через градиент CSPG [15].Это было эквивалентно обработке ChABC, которая удаляет сахарные цепи GAG, содержащие лиганд PTP σ . Введение пептидного миметика PTP σ после контузивной травмы спинного мозга также способствовало функциональному восстановлению как опорно-двигательной, так и мочевыводящей систем, что предполагает восстановление функциональных цепей [15]. Когда рецептор PTP σ отсутствует, животные модели SCI показывают повышенную регенерацию аксонов CST [166]. Об этом свидетельствует подавление РНКи-опосредованной регуляции PTP σ , что сводит на нет ингибирующие эффекты CSPG на рост отростков олигодендроцитов и миелинизацию.Такой же эффект наблюдался в олигодендроцитах, выделенных от мышей с нокаутом PTP σ [167]. Вместе эти результаты подчеркивают ингибирующую роль CSPGs в миграции, созревании и миелинизации OPC, ограничивая их репаративный потенциал после повреждения.
Об этом свидетельствует подавление РНКи-опосредованной регуляции PTP σ , что сводит на нет ингибирующие эффекты CSPG на рост отростков олигодендроцитов и миелинизацию.Такой же эффект наблюдался в олигодендроцитах, выделенных от мышей с нокаутом PTP σ [167]. Вместе эти результаты подчеркивают ингибирующую роль CSPGs в миграции, созревании и миелинизации OPC, ограничивая их репаративный потенциал после повреждения.
Интересно и вопреки широко распространенной демонстрации подавления регенерации аксонов, опосредованного глиальным рубцом, недавние данные лаборатории Софронева показали, что глиальный рубец оказывает благотворное влияние на отрастание и не только служит барьером для предотвращения дальнейшего повреждения, но и может быть выгодным или, по крайней мере, необходимым для процесса ремонта.В частности, в исследовании 2016 года, проведенном Андерсоном и его коллегами, изучающих восстановление спинного мозга на двух разных моделях трансгенных мышей с уменьшенным образованием глиальных рубцов либо путем делеции рубцовых астроцитов (TK + GCV; тимидинкиназа + ганцикловир), либо путем условного нокаута STAT3 ( преобразователь сигнала и активатор транскрипции 3), особенно в астроцитах, они не обнаружили увеличения регенерации, а в некоторых аксональных трактах (CST и восходящие сенсорные тракты) наблюдалось увеличение отмирания аксонов после травмы [168]. Несмотря на то, что он действует как физический барьер для возобновления роста аксонов, результаты этого исследования показывают, что глиальный рубец модулирует воспалительную реакцию после травмы и что без присутствия глиального рубца этот воспалительный ответ может быть эквивалентным барьером для возобновления роста [168]. Тем не менее, недавнее исследование, характеризующее различные фенотипы астроцитов, особенно в отношении участка поражения, было опубликовано в 2017 году Хара и соавторами [169]. В этом исследовании были охарактеризованы три различных подтипа астроцитов, связанных с глиальным рубцом: наивные астроциты, присутствующие до поражения, реактивные астроциты, которые первыми проникают в среду поражения и классифицируются как «переходные» с потенциальными нейропротективными и иммунорегулирующими. роли и рубцовые астроциты, которые составляют астроцитарный рубец, ингибирующий рост на поздней стадии [169].Интересно, что когда «переходные» (реактивные) астроциты были трансплантированы в наивный спинной мозг, они превратились в наивные астроциты; аналогично, они превращаются в астроциты, образующие рубцы, при трансплантации в место повреждения, демонстрируя, что окружающая среда диктует фенотип астроцитов и, следовательно, ингибирование, опосредованное глиальным рубцом [169].
Несмотря на то, что он действует как физический барьер для возобновления роста аксонов, результаты этого исследования показывают, что глиальный рубец модулирует воспалительную реакцию после травмы и что без присутствия глиального рубца этот воспалительный ответ может быть эквивалентным барьером для возобновления роста [168]. Тем не менее, недавнее исследование, характеризующее различные фенотипы астроцитов, особенно в отношении участка поражения, было опубликовано в 2017 году Хара и соавторами [169]. В этом исследовании были охарактеризованы три различных подтипа астроцитов, связанных с глиальным рубцом: наивные астроциты, присутствующие до поражения, реактивные астроциты, которые первыми проникают в среду поражения и классифицируются как «переходные» с потенциальными нейропротективными и иммунорегулирующими. роли и рубцовые астроциты, которые составляют астроцитарный рубец, ингибирующий рост на поздней стадии [169].Интересно, что когда «переходные» (реактивные) астроциты были трансплантированы в наивный спинной мозг, они превратились в наивные астроциты; аналогично, они превращаются в астроциты, образующие рубцы, при трансплантации в место повреждения, демонстрируя, что окружающая среда диктует фенотип астроцитов и, следовательно, ингибирование, опосредованное глиальным рубцом [169]. Было обнаружено, что это фенотипическое изменение в значительной степени контролируется коллагеном типа I, который обнаруживается в высоких уровнях в поражении и, как было показано, зависит от интегрин-N-кадгерина [169].Без вмешательства нормальная судьба астроцитов в непосредственной близости от поражения ЦНС направлена на формирование рубцового астроцитарного фенотипа, который действительно вреден для регенерации аксонов. Недавний комментарий Сильвера о ингибировании, опосредованном глиальным рубцом, и неспособности поражений млекопитающих образовывать стимулирующие рост астроглиальные мосты или иным образом усиливать опосредованное глиальным возобновление роста артикулирует эту проблему и дополнительно разъясняет необходимость лечения для противодействия глиальному рубцу [170]. например, с ChABC или другими.
Было обнаружено, что это фенотипическое изменение в значительной степени контролируется коллагеном типа I, который обнаруживается в высоких уровнях в поражении и, как было показано, зависит от интегрин-N-кадгерина [169].Без вмешательства нормальная судьба астроцитов в непосредственной близости от поражения ЦНС направлена на формирование рубцового астроцитарного фенотипа, который действительно вреден для регенерации аксонов. Недавний комментарий Сильвера о ингибировании, опосредованном глиальным рубцом, и неспособности поражений млекопитающих образовывать стимулирующие рост астроглиальные мосты или иным образом усиливать опосредованное глиальным возобновление роста артикулирует эту проблему и дополнительно разъясняет необходимость лечения для противодействия глиальному рубцу [170]. например, с ChABC или другими.
Ферментативная инактивация CSPG с помощью ChABC делает их неспособными взаимодействовать со своими рецепторами. Было показано, что это эффективно способствует пластичности и регенерации аксонов во многих моделях SCI. В частности, после поражения шейного позвоночника и лечения ChABC рост дорсального столба и волокон CST выходил за пределы повреждения по сравнению с контрольными, необработанными группами, в которых аксоны отводились от поражения [171]. В этом исследовании ChABC также способствовал восстановлению сенсорного и проприоцептивного поведения после травмы [171].Более того, в исследованиях, сочетающих лечение ChABC с реабилитацией передних конечностей после повреждения спинного столба, значительная регенерация аксонов и восстановление поведения также приводили при лечении во время травмы [172] или при отсроченном лечении через 4 недели после травмы [113]. В модели компрессионного ТСМ у крыс обработка ChABC способствовала дифференцировке олигодендроцитов и миелинизации эндогенных и трансплантированных клеток-предшественников нейронов [173, 174]. Лечение ChABC также способствовало миграции OPCs в место поражения и вокруг него после ушиба позвоночника [175].Ингибирующее действие CSPG на дифференцировку OPC и миелинизацию также нейтрализуется небольшими молекулами, такими как протамин, антагонист гепарина [176].
В частности, после поражения шейного позвоночника и лечения ChABC рост дорсального столба и волокон CST выходил за пределы повреждения по сравнению с контрольными, необработанными группами, в которых аксоны отводились от поражения [171]. В этом исследовании ChABC также способствовал восстановлению сенсорного и проприоцептивного поведения после травмы [171].Более того, в исследованиях, сочетающих лечение ChABC с реабилитацией передних конечностей после повреждения спинного столба, значительная регенерация аксонов и восстановление поведения также приводили при лечении во время травмы [172] или при отсроченном лечении через 4 недели после травмы [113]. В модели компрессионного ТСМ у крыс обработка ChABC способствовала дифференцировке олигодендроцитов и миелинизации эндогенных и трансплантированных клеток-предшественников нейронов [173, 174]. Лечение ChABC также способствовало миграции OPCs в место поражения и вокруг него после ушиба позвоночника [175].Ингибирующее действие CSPG на дифференцировку OPC и миелинизацию также нейтрализуется небольшими молекулами, такими как протамин, антагонист гепарина [176].
Было обнаружено, что присутствие CSPG в области поражения и вокруг него после ТСМ, а также при рассеянном склерозе отрицательно коррелирует с успешной ремиелинизацией. Наблюдения in vivo и in vitro показывают, что CSPG ограничивают миграцию и созревание OPC. В моделях демиелинизации индуцированная поражением активация CSPG снижает дифференцировку и развитие процесса OPCs.Удаление CSPG при лечении ксилозидом после индуцированной лизолецитином демиелинизации не только увеличивало количество OPC и олигодендроцитов, присутствующих в месте поражения, но также значительно улучшало ремиелинизацию [177]. Повышенная регуляция NG2, CSPG, обнаруженного на поверхности OPC, также наблюдалась после индуцированной лизолецитином демиелинизации. Ремиелинизация и репарация происходят только после удаления отложений NG2 с участка поражения, опосредованного матриксной металлопротеиназой-9 [178].
Поскольку ChABC является ферментом, у него есть свои ограничения; в первую очередь деградация с течением времени [163] и поэтому требует повторного дозирования для поддержания адекватной ферментативной активности, способствующей восстановлению. В ответ на это требование, долговременная экспрессия ChABC была достигнута в моделях SCI у крыс с использованием лентивируса или аденоассоциированного вируса [179], способствующего регенерации аксонов CST [180]. Однако неясно, может ли конститутивная экспрессия ChABC в течение длительного периода времени приводить к нежелательной пластичности в ЦНС.
В ответ на это требование, долговременная экспрессия ChABC была достигнута в моделях SCI у крыс с использованием лентивируса или аденоассоциированного вируса [179], способствующего регенерации аксонов CST [180]. Однако неясно, может ли конститутивная экспрессия ChABC в течение длительного периода времени приводить к нежелательной пластичности в ЦНС.
3.2. Опосредованное тенасцином С ингибирование пластичности ЦНС и регенерации аксонов
Помимо повышенных уровней CSPG, существует ряд онтогенетических белков, которые активируются после повреждения аксонов в зрелой ЦНС, включая тенасцины, семафорины, эфрины и нетрины [181] .В этом обзоре мы обсудим роль тенасцинов и семафоринов в росте и регенерации аксонов. Тенасцины — это внеклеточные гликопротеины, которые составляют составную часть ECM [182]. TN-C — это самый распространенный гликопротеин в ECM ЦНС, который высоко экспрессируется во время развития, но подавляется по мере созревания ЦНС млекопитающих. TN-C играет совершенно противоположные роли как в развивающейся, так и в зрелой ЦНС, действуя как молекула, способствующая росту в одних обстоятельствах, и молекула, ингибирующая рост в других. Например, в развивающейся ЦНС эти белки, особенно TN-C, способствуют пролиферации и миграции нервных предшественников наряду с удлинением аксонов, направлением и образованием конусов роста [7]. С другой стороны, характер иммуноокрашивания TN-C в развивающейся ЦНС демонстрирует, что он также может ингибировать рост, поскольку он способствует формированию границ между нейроанатомическими подразделениями, показывая его хеморепульсивную роль [183].
Например, в развивающейся ЦНС эти белки, особенно TN-C, способствуют пролиферации и миграции нервных предшественников наряду с удлинением аксонов, направлением и образованием конусов роста [7]. С другой стороны, характер иммуноокрашивания TN-C в развивающейся ЦНС демонстрирует, что он также может ингибировать рост, поскольку он способствует формированию границ между нейроанатомическими подразделениями, показывая его хеморепульсивную роль [183].
При повреждении зрелой ЦНС TN-C активируется и откладывается в ECM реактивными астроцитами, внося вклад в тормозящий глиальный рубец, который формируется после SCI (Table 1) [11, 12, 16-20].Напротив, уровни TN-R повышаются по мере созревания ЦНС млекопитающих из-за повышенного образования PNN, как обсуждалось выше [184]. Обычно TN-C оказывает ингибирующее действие на зрелые аксоны; однако, когда он связан со специфическим гетеродимерным рецептором интегрина, интегрином альфа9 бета1 ( α 9 β 1), который также подавляется в зрелой ЦНС, TN-C может способствовать росту нейритов и регенерации аксонов [17, 19]. К сожалению, в зрелой ЦНС сопутствующей активации рецептора интегрина α 9 β 1 не происходит.Интегрины, такие как гетеродимер α 9 β 1, высоко экспрессируются во время развития ЦНС на поверхности аксональных клеток, что способствует образованию конуса роста и удлинению аксонов. Было показано, что несбалансированная повышающая регуляция TN-C после повреждения без экспрессии его рецептора, способствующего росту, вносит существенный вклад в внутреннюю недостаточность регенерации аксонов. Подобно ситуации во время развития ЦНС, TN-C (в частности, связывающий домен фибронектина III, fnD) также играет противоположную роль, поскольку, как было показано, он необходим для регенерации [18].В частности, компрессионное поражение спинного мозга у взрослых мышей привело к более выраженному отмиранию отрубленных аксонов CST у мышей с нокаутом TN-C, что было спасено при замене TN-C посредством вирусно-опосредованной сверхэкспрессии fnD [18].
К сожалению, в зрелой ЦНС сопутствующей активации рецептора интегрина α 9 β 1 не происходит.Интегрины, такие как гетеродимер α 9 β 1, высоко экспрессируются во время развития ЦНС на поверхности аксональных клеток, что способствует образованию конуса роста и удлинению аксонов. Было показано, что несбалансированная повышающая регуляция TN-C после повреждения без экспрессии его рецептора, способствующего росту, вносит существенный вклад в внутреннюю недостаточность регенерации аксонов. Подобно ситуации во время развития ЦНС, TN-C (в частности, связывающий домен фибронектина III, fnD) также играет противоположную роль, поскольку, как было показано, он необходим для регенерации [18].В частности, компрессионное поражение спинного мозга у взрослых мышей привело к более выраженному отмиранию отрубленных аксонов CST у мышей с нокаутом TN-C, что было спасено при замене TN-C посредством вирусно-опосредованной сверхэкспрессии fnD [18].
Как стимулирующие рост, так и ингибирующие эффекты TN-C и TN-R были выяснены в ряде исследований. Интересно, что TN-C и TN-R проявляют противоположные функции при повреждении лицевого нерва. В частности, восстановление вибриссальных движений было хуже у мышей с дефицитом TN-C и лучше у мышей с дефицитом TN-R по сравнению с контрольной группой [185].Исследования одиночного нокаута TN-R и TN-C показывают нарушение LTP и пластичности in vivo и in vitro [186, 187]. У животных с дефицитом TN-C, подвергнутых гемисекции в поясничном отделе спинного мозга, было обнаружено повышенное количество нейрофиламент-положительных волокон вдоль гемисектированных краев спинного мозга [188]. Напротив, дефицит TN-R улучшает функциональное восстановление мышей после компрессии грудного отдела спинного мозга [189] с дальнейшими исследованиями с использованием антител против TN-R, показывающими улучшение восстановления после SCI [190].
Интересно, что TN-C и TN-R проявляют противоположные функции при повреждении лицевого нерва. В частности, восстановление вибриссальных движений было хуже у мышей с дефицитом TN-C и лучше у мышей с дефицитом TN-R по сравнению с контрольной группой [185].Исследования одиночного нокаута TN-R и TN-C показывают нарушение LTP и пластичности in vivo и in vitro [186, 187]. У животных с дефицитом TN-C, подвергнутых гемисекции в поясничном отделе спинного мозга, было обнаружено повышенное количество нейрофиламент-положительных волокон вдоль гемисектированных краев спинного мозга [188]. Напротив, дефицит TN-R улучшает функциональное восстановление мышей после компрессии грудного отдела спинного мозга [189] с дальнейшими исследованиями с использованием антител против TN-R, показывающими улучшение восстановления после SCI [190].
3.3. Семафорин-опосредованное ингибирование пластичности ЦНС и регенерации аксонов
Семафорины представляют собой класс мембранно-ассоциированных белков, участвующих в росте аксонов, управлении и формировании новых синаптических связей во время эмбрионального развития [191]. Семафорины также экспрессируются в ЦНС взрослого человека. Кроме того, исследования PNNs продемонстрировали, что семафорины sema3A и 3B связаны с CSPGs в структурах PNN, указывая тем самым, что эти семафорины вносят вклад в ингибирование и / или хеморепульсию роста аксонов, частично за счет своего вклада в PNN [148].В частности, sema3A был локализован в Wisteria floribunda agglutinin — (WFA-) -положительных PNN, ассоциация, которая была устранена после лечения ChABC [148]. Семафорины также были обнаружены в глиальных рубцах после травм (Таблица 1) [192–194]. В частности, было обнаружено, что Sema3A активируется после повреждения ЦНС [192] и аналогичным образом связан с предотвращением регенерации после SCI [195, 196]. Кроме того, было показано, что ингибитор sema3A, SM-216289, способствует регенерации аксонов, усиливает миграцию шванновских клеток к месту поражения, ингибирует апоптоз и значительно улучшает функциональное восстановление задней конечности после перерезки спинного мозга у взрослых крыс [ 195].
Семафорины также экспрессируются в ЦНС взрослого человека. Кроме того, исследования PNNs продемонстрировали, что семафорины sema3A и 3B связаны с CSPGs в структурах PNN, указывая тем самым, что эти семафорины вносят вклад в ингибирование и / или хеморепульсию роста аксонов, частично за счет своего вклада в PNN [148].В частности, sema3A был локализован в Wisteria floribunda agglutinin — (WFA-) -положительных PNN, ассоциация, которая была устранена после лечения ChABC [148]. Семафорины также были обнаружены в глиальных рубцах после травм (Таблица 1) [192–194]. В частности, было обнаружено, что Sema3A активируется после повреждения ЦНС [192] и аналогичным образом связан с предотвращением регенерации после SCI [195, 196]. Кроме того, было показано, что ингибитор sema3A, SM-216289, способствует регенерации аксонов, усиливает миграцию шванновских клеток к месту поражения, ингибирует апоптоз и значительно улучшает функциональное восстановление задней конечности после перерезки спинного мозга у взрослых крыс [ 195]. Интересно, что определенные подтипы семафорина, такие как sema4D, также были обнаружены в мембране миелинизирующих олигодендроцитов [197]. Более того, после повреждения ЦНС, sema4D активируется и может предотвращать рост аксонов in vitro, предполагая, что он может вносить вклад в миелин-ассоциированную ингибирующую среду в месте поражения in vivo [197].
Интересно, что определенные подтипы семафорина, такие как sema4D, также были обнаружены в мембране миелинизирующих олигодендроцитов [197]. Более того, после повреждения ЦНС, sema4D активируется и может предотвращать рост аксонов in vitro, предполагая, что он может вносить вклад в миелин-ассоциированную ингибирующую среду в месте поражения in vivo [197].
4. Резюме
Понимание физиологической регуляции роста аксонов и их пластичности в нервной системе взрослого человека имеет важное значение для разработки терапевтических стратегий.Пластичность и регенерация в ЦНС или их отсутствие в значительной степени регулируются образованием внеклеточного матрикса. По мере созревания ЦНС, приводящего к возникновению миелинизированных аксонов, PNN и ассоциированных белков, разумно понять, почему снижается способность к пластическим изменениям и восстановлению аксонов (суммировано на Рисунке 1). Эти экстранейрональные компоненты являются ключевыми для поддержания схемы, необходимой для нормального функционирования нервной системы, как на уровне сомы клеток и дендритов, так и на уровне аксонов. Таким образом, ЦНС имеет защитные меры для предотвращения или ограничения роста новых аксонов и / или связей, так что зрелые сети не подвергаются риску.Поэтому для ЦНС вредно позволять аксональному росту и пластичности (на грубом уровне) продолжаться в течение всей взрослой жизни, за пределами нормальной синаптической пластичности. Однако из литературы ясно, что ЦНС способна вызывать регенеративный ответ, если ингибирующие факторы (а) удаляются, создавая среду, способствующую росту, или (б) регулируются во времени, обеспечивая достаточное окно для регенерации. Продолжая понимать изменения, которые способствуют пластичности и восстановлению ПНС, а также те, которые препятствуют восстановлению ЦНС, мы, следовательно, можем оценить ключевые цели для терапевтического вмешательства.В будущем маловероятно, что единая терапевтическая стратегия будет успешной в полном содействии как регенерации аксонов, так и поведенческому / функциональному восстановлению после травмы спинного мозга. Подходы, которые решают проблемы как с внеклеточной средой ЦНС, так и с внутренней средой нейрона, имеют наилучшие возможности для успешного восстановления.
Таким образом, ЦНС имеет защитные меры для предотвращения или ограничения роста новых аксонов и / или связей, так что зрелые сети не подвергаются риску.Поэтому для ЦНС вредно позволять аксональному росту и пластичности (на грубом уровне) продолжаться в течение всей взрослой жизни, за пределами нормальной синаптической пластичности. Однако из литературы ясно, что ЦНС способна вызывать регенеративный ответ, если ингибирующие факторы (а) удаляются, создавая среду, способствующую росту, или (б) регулируются во времени, обеспечивая достаточное окно для регенерации. Продолжая понимать изменения, которые способствуют пластичности и восстановлению ПНС, а также те, которые препятствуют восстановлению ЦНС, мы, следовательно, можем оценить ключевые цели для терапевтического вмешательства.В будущем маловероятно, что единая терапевтическая стратегия будет успешной в полном содействии как регенерации аксонов, так и поведенческому / функциональному восстановлению после травмы спинного мозга. Подходы, которые решают проблемы как с внеклеточной средой ЦНС, так и с внутренней средой нейрона, имеют наилучшие возможности для успешного восстановления.
Сокращения
| α 9 β 1: | Alpha9 beta1 интегрин | |||
| AMPA: | α -амино-3-гидрокси-5-метил-4-изоксазолпропионовая кислота | |||
| BBB: | Гематоэнцефалический барьер | |||
| Bral2 / Hapln4: | Специфический для мозга связывающий белок 2 | |||
| Ca 2+ : | Кальций | |||
| ChABC: | Хондроитиназа ABC | |||
| CHO: | Клетки яичников китайского хомячка | |||
| CNP: | 2, 3-циклическая нуклеотидфосфодиэстераза | |||
| ЦНС: | Центральная нервная система | |||
| CP: | Критический период | |||
| Crtl1 / Hapln1: | Хрящевой связывающий белок 1 | |||
| CSPGs: | Хондроитинсульфат протеогликаны | |||
| CST: 90 047 | Кортикоспинальный тракт | |||
| DRG: | Ганглии задних корней (ганглии) | |||
| ECM: | Внеклеточный матрикс | |||
| FnD: | Связывающий домен фибронектина III | |||
| GAGcos цепи | ||||
| HAS: | Гиалуронансинтаза | |||
| K + : | Калий | |||
| LARG: | Связанный с лейкемией фактор обмена ро-гуанина | |||
| LINGO-1: | Лейцин- богатый повтор и белок-1, содержащий иммуноглобиноподобный домен | |||
| LTD: | Долгосрочная депрессия | |||
| LTP: | Долгосрочная потенциация | |||
| MAG: | Миелин-ассоциированный гликопротеин | |||
| MAI: | Миелин-ассоциированные ингибиторы | |||
| NG2: | Нейрон-глиальный антиген 2 | |||
| NgR1: | Рецептор Nogo-66 1 | |||
| NMDA: | N-метил-D-аспартат | |||
| NSPCs: | Нервные стволовые клетки и клетки-предшественники | |||
| OMgp: | Олигодендроцитарный миелиновый гликопротеин | |||
| OPCs: | Клетки-предшественники олигодендроцитов | |||
| p75NTR: | p75 рецептор нейротрофина | |||
| PirB | Парный иммуноглобальный рецептор :(миелинспецифический) протеолипидный белок | |||
| PNN: | Перинейрональные сети | |||
| PNS: | Периферическая нервная система | |||
| PTP σ : | Белковая тирозинфосфатаза сигма | RP β : | Фосфакан / протеин-тирозинфосфатаза рецепторного типа β | |
| SCI: | Повреждение спинного мозга | |||
| Sema3A: | Семафорин 3A | |||
| Sema4D: | Семафорин 4D | |||
| SIPR2: | Сфинголипид 1-фосфатный рецептор | ST48 | : | Преобразователь сигнала и активатор транскрипции — 3 |
| TK + GCV: | Тимидинкиназа + ганцикловир | |||
| TN-C: | Tenascin C | |||
| TN-R: | Tenascin R | |||
| TROY: | Член суперсемейства рецепторов фактора некроза опухоли. |
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов в отношении публикации этой статьи.
Вклад авторов
Шмма Курайш и Линдси Х. Форбс внесли равный вклад в эту работу.
Благодарности
Мелисса Р. Эндрюс и Шмма Курайш поддержаны исследовательским грантом Исследовательского совета биотехнологии и биологических наук (BBSRC).
Раков | Бесплатный полнотекстовый | Клеточная и молекулярная прогрессия рака простаты: модели для фундаментальных и доклинических исследований
1.Введение
Рак предстательной железы (РПЖ) является наиболее часто диагностируемым злокачественным новообразованием и второй по значимости причиной смерти от рака у мужчин в Соединенных Штатах. По оценкам, только в Соединенных Штатах в этом году от РПЖ пострадает около 191 930 мужчин и около 33 330 смертей [1]. Примечательно, что заболеваемость РПЖ и связанная с ним смертность почти на две трети и более чем в два раза выше, соответственно, у афроамериканцев (AA) по сравнению с их коллегами европейского происхождения (CA) [2,3].РПЖ следует определенному паттерну клеточной прогрессии, но проявляет разнообразную молекулярную патобиологию, что делает его одним из наиболее гетерогенных видов рака [4,5]. Тест на простатоспецифический антиген (ПСА) является основным инструментом выявления при скрининге РПЖ. Однако из-за отсутствия точности и специфичности полезность ПСА для диагностики РПЖ была поставлена под сомнение [6,7,8]. Большинство пациентов с РПЖ обычно подвергаются локальной радикальной простатэктомии, лучевой терапии, протонно-лучевой терапии и криохирургии после постановки первоначального диагноза [9,10,11].Однако для пациентов с метастатическим заболеванием или рецидивирующим раком с локально-регионарными и отдаленными метастазами основным методом лечения считается андрогенная депривация (АДТ) или кастрационная терапия [12]. К сожалению, несмотря на выдающийся первоначальный терапевтический ответ, у большинства пациентов с РПЖ, получавших АДТ, в конечном итоге наблюдается рецидив РПЖ в очень агрессивной и устойчивой к терапии форме, что приводит к плохим клиническим исходам [13,14].Для решения проблем, связанных с клиническим лечением рака простаты, исследовательские лаборатории по всему миру неустанно работают над пониманием лежащего в основе молекулярного разнообразия и биологии РПЖ.Эти усилия привели к созданию новых методов лечения, которые в настоящее время используются в клиниках, в то время как исследователи продолжают собирать новые идеи для решения новых проблем и неудач, с которыми сталкиваются в клинических условиях. Эти успехи стали возможны благодаря разработке нескольких исследовательских моделей in vitro и in vivo, в то время как новые модели продолжают разрабатываться для решения генетических и биологических сложностей, связанных с РПЖ. В этом обзоре мы обсуждаем клеточную и молекулярную прогрессию РПЖ, а также доступные модели in vitro и in vivo для исследования РПЖ.Мы считаем, что представленная здесь информация будет полезна исследователям, особенно новичкам в этой области, в понимании молекулярной патобиологии РПЖ и поможет им в выборе правильной модели (моделей) для их лабораторных и доклинических исследований.
2. Клеточное и молекулярное развитие рака простаты
Простата человека — это железистый орган размером с грецкий орех, который развивается из урогенитального синуса эмбриона [15]. Его основная функция — производить семенную жидкость, содержащую цинк, лимонную кислоту и различные ферменты, включая протеазу, называемую простатоспецифическим антигеном (ПСА).Гистологически простату можно разделить на центральную, периферическую и переходную зоны, состоящие из секреторной протоково-ацинарной структуры, расположенной внутри фиброзно-мышечной стромы [16,17]. Протоково-ацинарная структура образована высокими столбчатыми секреторными клетками просвета, уплощенным базальным эпителием, прикрепленным к базальной мембране, и разбросанными нейроэндокринными клетками (рис. 1). Клетки люминального эпителия экспрессируют цитокератины (CK) 8 и 18, NKX3.1, рецептор андрогенов (AR) и PSA, тогда как базальные эпителиальные клетки экспрессируют CK5, CK14, глутатион-S-трансферазу Pi 1 (GSTP1), p63 и низкие уровни AR [18,19].Клеточное происхождение рака простаты не очень ясно, отчасти из-за отсутствия хорошо охарактеризованных эпителиальных клонов простаты [20,21,22]. РПЖ развивается из нормального эпителия предстательной железы в результате многоступенчатого процесса гистологической трансформации, который регулируется различными лежащими в основе молекулярными изменениями [23] (Рисунок 2). Интраэпителиальная неоплазия простаты (PIN) низкой и высокой степени злокачественности развивается из нормального эпителия предстательной железы в результате потери фосфатазы и гомолога тензина (PTEN), NK3 Homeobox 1 (NKX3.1), сверхэкспрессия протоонкогена MYC, B-клеточной лимфомы 2 (BCL-2) и гена глутатион-S-трансферазы pi 1 (GSTP1) в сочетании с мутацией белка Speckle Type BTB / POZ (SPOP) и трансмембранной сериновой протеазой. 2-ETS-родственный ген (TMPRSS2-ERG) слияние [24,25,26,27,28,29,30,31,32,33,34,35,36]. Дальнейшая потеря белка ретинобластомы (RB1), наряду с активацией теломеразы и частой мутацией Forkhead Box A1 (FOXA1), приводит к развитию аденокарциномы простаты из распространенного поражения PIN [37,38,39,40,41,42,43 ].Дальнейшие молекулярные аберрации, включая потерю члена семейства SMAD 4 (SMAD4), корепрессоров AR, мутации в AR, FOXA1, BRCA1 / 2, ATM, ATR и RAD51, сопровождаемые усилением функции коактиватора AR, CXCL12, CXCR4, RANK -RANKL, EMT, BAI1 и EZh3 приводят к развитию метастатического рака простаты [44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59] . Как видно из модели прогрессирования РПЖ (рис. 2), инактивация PTEN, по-видимому, является критическим событием в канцерогенезе РПЖ и связана с агрессивным проявлением болезни.Изменения PTEN происходят различными путями при раке простаты, такими как делеция и перестройка генома, внутригенный разрыв или транслокация. Потеря PTEN связана с активацией передачи сигналов PI3K / AKT / mTOR, которая регулирует выживание, пролиферацию и энергетический метаболизм клеток [60,61]. Другой важной детерминантой онкогенеза РПЖ является SMAD4, ген-супрессор опухоли (18q21.1), который опосредует сигнальный путь трансформирующего фактора роста β (TGF-β) и подавляет рост эпителиальных клеток.Анализ транскриптома выявил значительно более низкие уровни SMAD4 в тканях РПЖ по сравнению с соседними доброкачественными тканями [46]. Следует отметить, что в модели на мышах специфическое удаление Smad4 и Pten из предстательной железы приводит к развитию инвазивного и метастатического потенциала РПЖ (обсуждается ниже) [45]. В каскаде инициации и прогрессирования РПЖ опухолевый супрессор NKX3.1 (8p21 ) играет ключевую роль и часто теряется из-за потери гетерозиготности (LOH) [62,63]. Следует отметить, что LOH на 8p21, по-видимому, является ранним событием в онкогенезе РПЖ [63,64,65].Таким образом, вероятно, что гены, которые находятся в этих часто удаленных областях, связаны с инициацией РПЖ. В нормальных условиях NKX3.1 оказывает подавляющее рост и дифференцирующее действие на эпителий простаты [66]. У гетерозиготных мышей Nkx3.1 развивается аномальная морфология простаты с диспластическим эпителием [67,68]. Важно отметить, что Nkx3.1-нулевые мыши обнаруживают изменения в морфологии эпителия простаты с тяжелой дисплазией [67]. Kim et al. продемонстрировали, что потеря функции Pten и Nkx3.1 у мышей участвовал в развитии РПЖ. Важно отметить, что сложные мутантные мыши по Pten; Nkx3.1 показали более высокую частоту интраэпителиальной неоплазии простаты высокой степени (HGPIN) [69]. В дополнение к критическим генам-супрессорам опухолей, описанным выше, протоонкоген MYC также амплифицируется при РПЖ [70,71,72]. MYC кодирует фактор транскрипции, который регулирует экспрессию нескольких генов, участвующих в пролиферации клеток, метаболизме, функции митохондрий и обновлении стволовых клеток [73,74,75]. Несколько исследований предполагают, что MYC активируется за счет сверхэкспрессии, амплификации, перестройки, активации пути Wnt / β-катенина, вариации промотора MYC зародышевой линии и потери FOXP3 при РПЖ [76,77,78,79], и является движущей силой критического онкогенного события. Инициирование и прогрессирование РПЖ [71,80].Помимо MYC, слияние генов TMPRSS2: ERG, возникающее в результате хромосомной перестройки, также встречается примерно в 45% случаев РПЖ. Это изменение приводит к экспрессии усеченного белка ERG под контролем промотора контрольного андроген-чувствительного гена TMPRSS2 [81,82,83,84,85]. ERG принадлежит к семейству транскрипционных факторов ETS (ERG, ETV1 и ETV4), и его активация связана с прогрессированием РПЖ как на ранних, так и на поздних стадиях [82,83,86]. MYB, другой ген, кодирующий фактор транскрипции, также, как сообщается, амплифицируется при РПЖ и демонстрирует повышенную частоту амплификации при РПЖ, устойчивом к кастрации (CRPC) [87].Исследования нашей лаборатории показали, что MYB играет жизненно важную роль в росте РПЖ, злокачественном поведении и устойчивости к истощению андрогенов [56].Генная терапия в ЦНС
Bowers W, Howard D, Federoff H. Стратегии генной терапии для нейрозащиты: последствия для болезни Паркинсона Exp Neurol 1997 144 : 58–68
CAS PubMed Google Scholar
Раймон Х, Тод С, Гейдж Ф.Применение генной терапии ex vivo в лечении болезни Паркинсона Exp Neurol 1997 144 : 82–91
CAS PubMed Google Scholar
Картер Б.С., Зервас Н.Т., Чиокка Е.А. Нейрогенетическая хирургия: текущие ограничения и перспективы генной и вирусной терапии Clin Neurosurg 1999 45 : 226–246
CAS PubMed Google Scholar
Фуэйо Дж., Гомес-Манзано К., Юнг В.К., Кирицис А.П.Таргетинг в генной терапии глиом Arch Neurol 1999 56 : 445–448
CAS PubMed Google Scholar
Boviatsis EJ et al . Долгосрочная выживаемость крыс с новообразованиями головного мозга, получавших ганцикловир и вектор вируса простого герпеса, который сохраняет интактный ген тимидинкиназы Cancer Res 1994 54 : 5745–5751
CAS Google Scholar
Хак Н, Исаксон О.Антисмысловая генная терапия нейродегенеративного заболевания? Exp Neurol 1997 144 : 139–146
CAS PubMed Google Scholar
Chirmule N и др. . Иммунные ответы на аденовирус и аденоассоциированный вирус у людей Генная терапия 1999 6 : 1574–1583
CAS Google Scholar
Парр М.Дж. и др. .Параметры иммунитета, влияющие на генную терапию аденовирусным вектором в головном мозге J Нейровирус 1998 4 : 194–203
CAS Google Scholar
Ян В., Мейсон В., Саммерс Дж. Ковалентно замкнутая кольцевая вирусная ДНК, образованная из двух типов линейной ДНК в печени, инфицированной вирусом гепатита сурков J Virol 1996 70 : 4567–4575
CAS PubMed PubMed Central Google Scholar
Ho DY, McLaughlin JR, Sapolsky RM.Индуцируемая экспрессия генов из дефектных векторов вируса простого герпеса с использованием системы промотора, чувствительной к тетрациклину Mol Brain Res 1996 41 : 200–209
CAS PubMed PubMed Central Google Scholar
Лу Б, Федерофф Х.Дж. Векторы ампликона вируса простого герпеса типа 1 с экспрессией индуцируемого глюкокортикоидами гена Hum Gene Ther 1995 6 : 419–428
CAS PubMed Google Scholar
Manome Y и др. .Экспрессия трансгена в злокачественной глиоме с использованием дефектного по репликации аденовирусного вектора, содержащего промотор Egr-1: активация ионизирующим излучением или поглощение радиоактивного йододезоксиуридина Hum Gene Ther 1998 9 : 1409–1417
CAS Google Scholar
Oligino T и др. . Экспрессия трансгена, индуцируемая лекарственными средствами, в головном мозге с использованием вектора вируса простого герпеса Генная терапия 1998 5 : 491–496
CAS PubMed Google Scholar
Ye X и др. .Регулируемая доставка терапевтических белков после in vivo переноса гена соматических клеток Science 1999 283 : 88–91
CAS PubMed PubMed Central Google Scholar
Каплитт М.Г. и др. . Долгосрочная экспрессия генов и фенотипическая коррекция с использованием аденоассоциированных вирусных векторов в мозге млекопитающих Nat Genet 1994 8 : 148–154
CAS PubMed PubMed Central Google Scholar
Klein RL и др. .Нейрон-специфическая трансдукция в эптогиппокампе или нигростриатном пути крыс с помощью рекомбинантных аденоассоциированных вирусных векторов Exp Neurol 1998 150 : 183–194
CAS PubMed Google Scholar
Peel AL и др. . Эффективная трансдукция зеленого флуоресцентного белка в нейронах спинного мозга с использованием аденоассоциированных вирусных векторов, содержащих промоторы, специфичные для определенного типа клеток. Gene Therapy 1997 4 : 16–24
CAS PubMed Google Scholar
Song S et al .Вектор HSV-1, содержащий промотор тирозингидроксилазы крысы, усиливает как долгосрочную, так и специфичную для клеточного типа экспрессию в среднем мозге J Neurochem 1997 68 : 1792–1803
CAS PubMed Google Scholar
Биланг-Блеуэл А и др. . Интрастриатальная инъекция аденовирусного вектора, экспрессирующего нейротрофический фактор линии глиальных клеток, предотвращает дегенерацию дофаминергических нейронов и нарушение поведения на модели болезни Паркинсона на крысах Proc Natl Acad Sci USA 1997 94 : 8818–8823
CAS PubMed Google Scholar
Davar G et al .Сравнительная эффективность экспрессии генов, доставленных в сенсорные нейроны мыши с векторами вируса герпеса J Comp Neurol 1994 339 : 3–11
CAS PubMed Google Scholar
Maidment NT et al . Экспрессия репортерного гена lacZ в базальном переднем мозге, гиппокампе и нигростриатном пути крыс с использованием нереплицирующегося вируса простого герпеса Exp Neurol 1996 139 : 107–114
CAS PubMed Google Scholar
Herrlinger U et al .Векторы HSV-1 для терапии экспериментальных опухолей ЦНС. В: Методы в молекулярной медицине, протоколы генной терапии Human Press Inc: Totowa, NJ (в печати)
Dewey RA et al . Хроническое воспаление головного мозга и стойкая экспрессия HSV1-TK у лиц, переживших сингенную глиому, пролеченных аденовирусной генной терапией: значение для клинических испытаний Nature Med 1999 5 : 1256–1263
CAS PubMed Google Scholar
Ройзман Б., Сирс А.Е.Вирусы простого герпеса и их размножение. В: Fields BN, Knipe DM, Howley PM (eds) Fields Virology Lippincott-Raven: Philadelphia 1996 pp 2231–2296
Google Scholar
Sodeik B, Ebersold MW, Helenius A. Опосредованный микротрубочками транспорт поступающих капсидов вируса простого герпеса 1 в ядро J Cell Biol 1997 136 : 1007–1021
CAS PubMed PubMed Central Google Scholar
Держатель EL и др. .Аксоплазма кальмаров поддерживает ретроградный аксональный транспорт вируса простого герпеса Biol Bull 1999 197 : 257–258
CAS PubMed PubMed Central Google Scholar
Якобс А и др. . Функциональная совместная экспрессия тимидинкиназы HSV-1 и зеленого флуоресцентного белка (TKGFP): значение для визуализации экспрессии терапевтического гена Неоплазия (в печати)
Glorioso JC et al .Вирус простого герпеса как вектор доставки генов для центральной нервной системы. В: Viral Vectors 1–23 Academic Press: New York 1995
Krisky DM et al . Делеция нескольких немедленных и ранних генов вируса простого герпеса снижает цитотоксичность и обеспечивает долгосрочную экспрессию генов в нейронах. Генная терапия 1998 5 : 1593–1603
CAS Google Scholar
Wu P, Phillips MI, Bui J, Terwilliger EF.Интеграция трансгена, опосредованная аденоассоциированным вирусом, в нейроны и другие неделящиеся клетки-мишени J Virol 1998 72 : 5919–5926
CAS PubMed PubMed Central Google Scholar
Samaniego LA, Neiderhiser L, DeLuca NA. Персистенция и экспрессия генома вируса простого герпеса в отсутствие немедленных ранних белков J Virol 1998 72 : 3307–3320
CAS PubMed PubMed Central Google Scholar
Ho DY, Mocarski ED.Бета-галактозидаза как маркер в периферических и нервных тканях мышей, инфицированных вирусом простого герпеса Virology 1988 167 : 279–283
CAS PubMed Google Scholar
Гоинс Б., Рудольф А.С., Лиглер Ф.С. Гемоглобин, инкапсулированный в липосомы: историческое развитие кровезаменителя Biotech 1991 19 : 117–125
CAS Google Scholar
Dobson AT и др. .Латентный, непатогенный вектор, производный от HSV-1, стабильно экспрессирует бета-галактозидазу в нейронах мыши Neuron 1990 5 : 353–360
CAS PubMed Google Scholar
Спаете Р., Френкель Н. Ампликон вируса герпеса: новый вектор клонирования-амплификации эукариотического дефектного вируса Cell 1982 30 : 295–304
CAS PubMed PubMed Central Google Scholar
Fraefel C et al .Безвирусный перенос плазмидных векторов вируса простого герпеса типа 1 в нервные клетки J Virol 1996 70 : 7190–7197
CAS PubMed PubMed Central Google Scholar
Saeki Y et al . ДНК вируса простого герпеса типа 1, амплифицированная как бактериальная искусственная хромосома в Escherichia coli : спасение репликационно-компетентного потомства вируса и упаковка ампликонных векторов Hum Gene Ther 1998 9 : 2787–2794
CAS PubMed Google Scholar
Ставропулос Т.А., Стратди, Калифорния.Усовершенствованная система упаковки для векторов вируса простого герпеса, зависимых от помощников J Virol 1998 72 : 7173–7143
Google Scholar
Константини L и др. . Перенос генов в нигростриатную систему с помощью векторов ампликонов гибридного вируса простого герпеса / аденоассоциированного вируса Hum Gene Ther 1999 10 : 2481–2494
Google Scholar
Wang AU и др. .Векторы ампликона HSV-1 представляют собой высокоэффективную систему доставки генов для миобластов и мышечных трубок скелетных мышц. Am J Phys (в печати)
Fraefel C, Jacoby DR, Breakefield XO. Последние разработки в области векторных систем ампликона на основе вируса простого герпеса типа 1. In: Advances in Virus Research Academic Press: New York (в печати)
Muzyczka N. Использование аденоассоциированного вируса в качестве общего вектора трансдукции для клеток млекопитающих Curr Topics Microbiol / Immunol 1992 148 : 97–129
Google Scholar
Сяо X и др. .Перенос антисмыслового гена вектора аденоассоциированного вируса (AAV) in vivo снижает рецепторы, содержащие ГАМК (A) альфа 1, и увеличивает чувствительность к припадкам нижних колликулярных узлов Brain Res 1997 756 : 76–83
CAS PubMed Google Scholar
Котин Р.М., Линден Р.М., Бернс К.И. Характеристика предпочтительного сайта на хромосоме 19q человека для интеграции ДНК аденоассоциированного вируса путем негомологичной рекомбинации EMBO J 1992 11 : 5071–5078
CAS PubMed PubMed Central Google Scholar
Balague C, Kalla M, Zhang WW.Белок Rep78 аденоассоциированного вируса и концевые повторы усиливают интеграцию последовательностей ДНК в клеточный геном J Virol 1997 71 : 3299–3306
CAS PubMed PubMed Central Google Scholar
Walker SL, Wondreling RS, Owens RA. Мутационный анализ белка Rep68 аденоассоциированного вируса: идентификация критических остатков, необходимых для сайт-специфической эндонуклеазной активности J Virol 1997 71 : 2722–2730
CAS PubMed PubMed Central Google Scholar
Weitzman MD, Kyostio SR, Kotin RM, Owens RA.Белки Rep аденоассоциированного вируса (AAV) опосредуют образование комплекса между ДНК AAV и местом ее интеграции в ДНК человека Proc Natl Acad Sci USA 1994 91 : 5808–5812
CAS PubMed Google Scholar
Ян CC и др. . Пути клеточной рекомбинации и структуры шпилечных концевых повторов вируса достаточны для интеграции аденоассоциированного вируса in vivo и in vitro J Virol 1997 71 : 9231–9247
CAS PubMed PubMed Central Google Scholar
Ferrari FK, Xiao X, McCarty D, Samulski RJ.Новые разработки в области создания векторов для генной терапии rAAV с высоким титром без Ad Nature Med 1997 3 : 1295–1297
CAS PubMed Google Scholar
Сяо Х, Ли Дж., Самульски Р.Дж. Получение рекомбинантных аденоассоциированных вирусных векторов с высоким титром в отсутствие аденовируса-помощника J Virol 1998 72 : 2224–2232
CAS PubMed PubMed Central Google Scholar
Summerford C, Samulski R.Связанный с мембраной гепарансульфат протеогликан является рецептором для сборки аденоассоциированного вируса типа 2 J Virol 1998 72 : 1438–1445
CAS PubMed PubMed Central Google Scholar
Гримм Д., Керн А., Риттнер К., Кляйншмидт Дж. Новые инструменты для производства и очистки рекомбинантных векторов аденоассоциированного вируса Hum Gene Ther 1998 9 : 2745–2760
CAS PubMed PubMed Central Google Scholar
Bartlett JS, Samulski RJ, McCown T.Селективное и быстрое проникновение аденоассоциированного вируса типа 2 в мозг Hum Gene Ther 1998 9 : 1181–1186
CAS PubMed PubMed Central Google Scholar
Каплитт М.Г. и др. . Промотор проэнкефалина обеспечивает регионально-специфическую и долгосрочную экспрессию во взрослом мозге после in vivo переноса гена через дефектный вирусный вектор простого герпеса Proc Natl Acad Sci USA 1994 91 : 8979–8983
CAS PubMed Google Scholar
Мандель Р.Дж. и др. .Характеристика интрастриатального рекомбинантного аденоассоциированного вируса-опосредованного переноса генов тирозингидроксилазы человека и GTP-циклогидролазы I человека на модели болезни Паркинсона у крыс J Neurosci 1998 18 : 4271–4284
CAS PubMed Google Scholar
Lo W и др. . Перенос гена в мозг, опосредованный аденоассоциированным вирусом: продолжительность и модуляция экспрессии Hum Gene Ther 1999 10 : 201–213
CAS PubMed Google Scholar
McCown TJ и др. .Дифференциальные и устойчивые паттерны экспрессии переноса гена ЦНС вектором аденоассоциированного вируса (AAV) Brain Res 1996 713 : 99–107
CAS PubMed Google Scholar
Бартлетт Дж., Кляйншмидт Дж., Бушер Р., Самульски Р. Направленная аденоассоциированная вирусная векторная трансдукция непермиссивных клеток, опосредованная биспецифическим антителом F (ab’gamma) 2 Nat Biotechnol 1999 17 : 181–186
CAS PubMed PubMed Central Google Scholar
Girod A и др. .Модификация генетического капсида обеспечивает эффективное перенацеливание аденоассоциированного вируса типа 2 Nature Med 1999 5 : 1052–1056
CAS PubMed Google Scholar
Хаберман Р., Маккаун Т., Самульски Р. Индуцируемая долговременная экспрессия генов в головном мозге с переносом генов аденоассоциированного вируса Генная терапия 1998 5 : 1604–1611
CAS PubMed PubMed Central Google Scholar
Rivera V и др. .Долгосрочная регулируемая экспрессия гормона роста у мышей после внутримышечного переноса гена Proc Natl Acad Sci USA 1999 96 : 8657–8662
CAS PubMed PubMed Central Google Scholar
Ян Y и др. . Клеточный иммунитет к вирусным антигенам ограничивает аденовирусы с делецией E1 для генной терапии Proc Natl Acad Sci USA 1994 91 : 4407–4411
CAS PubMed PubMed Central Google Scholar
Dai Y и др. .Клеточные и гуморальные иммунные ответы на аденовирусные векторы, содержащие ген фактора IX: толеризация фактора IX и векторных антигенов обеспечивает долгосрочную экспрессию Proc Natl Acad Sci USA 1995 92 : 1401–1405
CAS PubMed PubMed Central Google Scholar
Zhou H, O’Neal W., Morral N, Beaudet AL. Разработка линии комплементарных клеток и системы для конструирования аденовирусных векторов с удаленными E1 и E2a J Virol 1996 70 : 7030–7038
CAS PubMed PubMed Central Google Scholar
Gao GP, Yang Y, Wilson JM.Биология аденовирусных векторов с делециями E1 и E4 для генной терапии, направленной на печень J Virol 1996 70 : 8934–8943
CAS PubMed PubMed Central Google Scholar
Armentano D и др. . Необходимость E40RF3 для достижения долговременной экспрессии трансгена из промотора цитомегаловируса в аденовирусных векторах J Virol 1999 8 : 7031–7034
Google Scholar
Илан Y и др. .Вставка аденовирусной области E3 в рекомбинантный вирусный вектор предотвращает противовирусные гуморальные и клеточные иммунные ответы и обеспечивает долгосрочную экспрессию гена Proc Natl Acad Sci USA 1997 94 : 2587–2592
CAS PubMed PubMed Central Google Scholar
Энгельгардт Дж. Ф., Литцки Л., Уилсон Дж. М.. Длительная экспрессия трансгена в легких хлопковой крысы с рекомбинантными аденовирусами, дефектными по E2a Hum Gene Ther 1994 10 : 1217–1229
Google Scholar
Ян Y и др. .Инактивация E2a в рекомбинантных аденовирусах улучшает перспективы генной терапии муковисцидоза Nat Genet 1994 7 : 362–369
CAS PubMed Google Scholar
Wang Q и др. . Устойчивая экспрессия трансгена в печени мыши после переноса гена in vivo с помощью аденовирусного вектора дельта E1 / дельта E4 Генная терапия 1997 4 : 393–400
CAS PubMed Google Scholar
Fang B и др. .Отсутствие персистенции E1-рекомбинантных аденовирусных векторов, содержащих чувствительную к температуре мутацию E2A, у иммунокомпетентных мышей и собак с гемофилией B Генная терапия 1996 3 : 217–222
CAS PubMed Google Scholar
Фишер К.Дж. и др. . Рекомбинантный аденовирус с делецией всех вирусных генов для генной терапии муковисцидоза Virology 1996 217 : 11–22
CAS Google Scholar
Кочанек С и др. .Новый аденовирусный вектор: замена всех вирусных кодирующих последовательностей на 28 т.п.н. ДНК, независимо экспрессирующих как полноразмерный дистрофин, так и бета-галактозидазу. Proc Natl Acad Sci USA 1996 93 : 5731–5736
CAS Google Scholar
Харди S и др. . Конструирование аденовирусных векторов посредством рекомбинации Cre-lox J Virol 1997 71 : 1842–1849
CAS PubMed PubMed Central Google Scholar
Морси М.А. и др. .Аденовирусный вектор, удаленный для всех вирусных кодирующих последовательностей, приводит к повышенной безопасности и расширенной экспрессии трансгена лептина Proc Natl Acad Sci USA 1998 95 : 7866–7871
CAS Google Scholar
Кумар-Сингх Р., Фарбер ДБ. Инкапсидированная аденовирусная мини-хромосома-опосредованная доставка генов к сетчатке: применение для спасения дегенерации фоторецепторов Hum Mol Genet 1998 7 : 1893–1900
CAS PubMed Google Scholar
Либер А., Хе К.Ю., Кей М.А.Аденовирусный претерминальный белок стабилизирует мини-аденовирусные геномы in vitro и in vivo Nat Biotechnol 1997 15 : 1383–1387
CAS PubMed Google Scholar
Thomas CE et al . Периферическая инфекция аденовирусом вызывает неожиданное длительное воспаление головного мозга у животных, которым внутричерепно вводили аденовирусные векторы первого поколения, но не высокопроизводительные: к реалистичной долгосрочной неврологической генной терапии хронических заболеваний (представлено)
Harui A , Сузуки С, Кочанек С, Митани К.Частота и стабильность хромосомной интеграции аденовирусных векторов J Virol 1999 73 : 6141–6146
CAS PubMed PubMed Central Google Scholar
Overturf K и др. . Аденовирусная генная терапия на мышиной модели наследственной тирозинемии типа I Hum Gene Ther 1997 8 : 513–521
CAS PubMed Google Scholar
Horwitz MS.Аденовирусы. В: Филдс Б.Н., Книп Д.М., Хоули П.М. (ред.). Fields Virology , 3-е изд. Lippincott Raven Publishers: Philadelphia 1996 2149–2173
Mulligan RC. Основы генной терапии Наука 1993 260 : 926–932
CAS PubMed PubMed Central Google Scholar
Миллер Д.Г., Адам М.А., Миллер А.Д. Перенос генов ретровирусными векторами происходит только в клетках, которые активно реплицируются во время инфекции Mol Cell Biol 1990 10 : 4239–4242
CAS PubMed PubMed Central Google Scholar
Cepko CL и др. .Анализ происхождения с использованием ретровирусных векторов Curr Top Dev Biol 1998 36 : 51–74
CAS PubMed Google Scholar
Bankiewicz KS и др. . Практические аспекты разработки ex vivo и in vivo генной терапии болезни Паркинсона Exp Neurol 1997 114 : 147–156
Google Scholar
Martinez-Serrano A, Bjorklund A.Иммортализованные клетки-предшественники нейронов для переноса и восстановления генов ЦНС Trends in Neurosci 1997 20 : 530–538
CAS Google Scholar
Fisher L и др. . Выживание и функция фибробластов, трансплантированных интрастриатально, генетически модифицированных для продуцирования L-допа Neuron 1990 6 : 371–380
Google Scholar
Джонстон К. и др. .Гибридные ампликонные векторы HSV / AAV увеличивают экспрессию трансгена в клетках глиомы человека Hum Gene Ther 1997 8 : 359–370
CAS PubMed PubMed Central Google Scholar
Sena-Esteves M и др. . Одностадийное преобразование клеток в продуценты ретровирусных векторов с помощью гибридных ампликонов HSV / EBV J Virol 1999 73 : 10426–10439
CAS PubMed PubMed Central Google Scholar
Palombo F и др. .Сайт-специфическая интеграция в клетках млекопитающих, опосредованная новым вектором гибридного бакуловируса и аденоассоциированного вируса J Virol 1998 72 : 5025–5034
CAS PubMed PubMed Central Google Scholar
Ван С., Вос Дж.М. Гибридный инфекционный вектор вируса герпеса на основе вируса Эпштейна-Барра и вируса простого герпеса типа 1 для переноса генов в клетки человека in vitro и in vivo J Virol 1996 70 : 8422–8430
CAS PubMed PubMed Central Google Scholar
Йейтс Дж., Уоррен Н., Сагден Б.Стабильная репликация плазмид, полученных из вируса Эпштейна-Барра, в различных клетках млекопитающих Nature 1985 313 : 812–815
CAS PubMed PubMed Central Google Scholar
Recchia A и др. . Сайт-специфическая интеграция, опосредованная вектором гибридного аденовируса и аденоассоциированного вируса Proc Natl Acad Sci USA 1999 96 : 2615–2620
CAS PubMed Google Scholar
Фишер К. и др. .Новый гибридный вектор аденовируса-аденоассоциированного вируса, который демонстрирует эффективное восстановление и доставку генома AAV. Hum Gene Ther 1996 7 : 2079–2087
CAS PubMed PubMed Central Google Scholar
Бильбао G и др. . Химеры с аденовирусными / ретровирусными векторами: новая стратегия для достижения высокоэффективной стабильной трансдукции in vivo FASEBJ 1997 11 : 624–634
CAS Google Scholar
Feng M et al .Стабильная трансдукция гена in vivo с помощью нового аденовирусного / ретровирусного химерного вектора Nat Biotechnol 1997 15 : 866–870
CAS PubMed Google Scholar
Naldini L и др. . Доставка in vivo и стабильная трансдукция неделящихся клеток лентивирусным вектором Science 1996 272 : 263–267
CAS Google Scholar
Кафри Т. и др. .Устойчивая экспрессия доставки генов непосредственно в печень и мышцы лентивирусными векторами Nat Genet 1997 17 : 314–317
CAS PubMed Google Scholar
Naldini L и др. . Эффективный перенос, интеграция и устойчивая долгосрочная экспрессия трансгена в мозге взрослых крыс, которым вводили лентивирусный вектор Proc Natl Acad Sci USA 1996 93 : 11382–11388
CAS PubMed Google Scholar
Zufferey R et al .Множественно ослабленный лентивирусный вектор обеспечивает эффективную доставку гена in vivo Nat Biotechnol 1997 15 : 871–875
CAS Google Scholar
Кафри Т. и др. . Линия упаковочных клеток для лентивирусных векторов J Virol 1999 73 : 576–584
CAS PubMed PubMed Central Google Scholar
Miyoshi H и др. .Разработка самоинактивирующегося лентивирусного вектора J Virol 1998 72 : 8150–8157
CAS PubMed PubMed Central Google Scholar
Ивакума Т, Цуй Й, Чанг Л. Самоинактивирующиеся лентивирусные векторы с модификациями U3 и U5 Вирол 1999 26 : 120–132
Google Scholar
Белый S и др. .Лентивирусные векторы с использованием элементов вируса иммунодефицита человека и обезьян 1 J Virol 1999 73 : 2832–2840
CAS PubMed PubMed Central Google Scholar
Бломер У. и др. . Высокоэффективный и устойчивый перенос генов во взрослых нейронах с лентивирусным вектором J Virol 1997 71 : 6641–6649
PubMed PubMed Central Google Scholar
Цзяо С., Ченг Л., Вольф Дж. А., Ян Н. С..Перенос и экспрессия генов, опосредованные бомбардировкой частиц, в тканях мозга крыс Biotech 1993 11 : 497–502
CAS Google Scholar
Wolff JA et al . Прямой перенос гена в мышцу мыши in vivo Наука 1990 247 : 1465–1468
CAS Google Scholar
Wagner E et al .Связывание аденовируса с комплексами трансферрин-полилизин / ДНК значительно усиливает опосредованную рецептором доставку и экспрессию трансфицированных генов Proc Natl Acad Sci USA 1992 89 : 6099–7103
CAS PubMed PubMed Central Google Scholar
Huckett B, Ariatti M, Hawtrey AO. Доказательства адресного переноса генов посредством рецептор-опосредованного эндоцитоза. Стабильная экспрессия после инсулино-направленного проникновения NEO в клетки HepG2 Biochem Pharmacol 1990 40 : 253–263
CAS PubMed Google Scholar
Wagner E, Cotten M, Foisner R, Birnstiel ML.Комплексы трансферрин-поликатион-ДНК: влияние поликатионов на структуру комплекса и доставку ДНК в клетки Proc Natl Acad Sci USA 1991 88 : 4255–4259
CAS Google Scholar
Saeki Y и др. . Разработка и характеристика катионных липосом, конъюгированных с HVJ (вирус Сендай): обратный эффект катионного липида для in vitro и in vivo Перенос гена Hum Gene Ther 1997 8 : 2133–2141
CAS PubMed Google Scholar
Aronsohn AI, Hughes JA.Сигнальные пептиды ядерной локализации усиливают генную терапию, опосредованную катионными липосомами J Drug Target 1998 5 : 163–169
CAS PubMed Google Scholar
Намики Ю., Такахаши Т., Оно Т. Генная трансдукция диссеминированной внутрибрюшинной опухоли с использованием катионных липосом, содержащих белки негистонового хроматина: катионная липосомная генная терапия карциноматоза Генная терапия 1998 5 : 240–246
CAS PubMed Google Scholar
Канеда Й, Саеки Й, Моришита Р.Генная терапия с использованием липосом HVJ: лучшее из обоих миров? Mol Med Сегодня 1999 5 : 298–303
CAS PubMed Google Scholar
Schwartz B и др. . Перенос генов с помощью голой ДНК в мозг взрослой мыши Генная терапия 1996 3 : 405–411
CAS PubMed Google Scholar
Брукс А.И. и др. .Воспроизводимая и эффективная доставка гена ЦНС мыши с использованием инжектора, управляемого микропроцессором. J Neurosci Meth 1998 80 : 137–147
CAS Google Scholar
Райнов Н.Г., Брейкфилд ХО, Крамм СМ. Способы применения вектора для генной терапии опухолей головного мозга Генная терапия и биология молей (в печати)
Muldoon LL et al . Доставка лечебных средств к опухолям головного мозга и внутримозговых опухолей.В: Chiocca EA, Breakefield XO (ред.). Генная терапия неврологических заболеваний Humana Press: Boston, MA 1998 295–311
Google Scholar
Короткий MP и др. . Доставка гена в клетки глиомы в головном мозге крысы путем трансплантации ретровируса, упаковывающего клеточную линию J Neurosci Res 1990 27 : 427–439
CAS PubMed PubMed Central Google Scholar
Goldsmith KT, Curiel DT, Engler JA, Garver RIJ. Транс Комплементация аденовируса с удаленным E1A с кодированными последовательностями E1A для получения рекомбинантных клеток-продуцентов аденовирусов Hum Gene Ther 1994 5 : 1341–1348
CAS PubMed Google Scholar
Boviatsis EJ et al . Перенос генов в экспериментальные опухоли головного мозга, опосредованный аденовирусом, вирусом простого герпеса (HSV) и ретровирусными векторами Hum Gene Ther 1994 5 : 183–191
CAS PubMed Google Scholar
Aboody KS и др. .Новая платформа для генной терапии опухолей головного мозга: чужеродный ген, экспрессирующий нервные стволовые клетки, проявляет тропизм к внутричерепным глиомам (представлено)
Gage FH. Стволовые клетки центральной нервной системы Curr Opin Neurobiol 1998 8 : 671–676
CAS PubMed Google Scholar
Лал Б и др. . Имплантация эндотелиальных клеток и выживаемость в экспериментальных глиомах Proc Natl Acad Sci USA 1994 91 : 9695–9699
CAS PubMed Google Scholar
Uteza Y и др. .Внутривитловая трансплантация инкапсулированных фибробластов, секретирующих фактор роста фибробластов человека 2, задерживает дегенерацию фоторецепторных клеток у крыс Королевского колледжа хирургов Proc Natl Acad Sci USA 1999 96 : 3126–3131
CAS PubMed Google Scholar
Крамм CM и др. . Опосредованная вектором герпеса доставка гена тимидинкиназы к диссеминированным опухолям ЦНС Hum Gene Ther 1996 7 : 291–300
CAS PubMed Google Scholar
Снайдер Е.Ю., Тейлор Р.М., Вулф Дж. Х.Приживление нервных клеток-предшественников корректирует лизосомные накопления в мозге мыши MPS VII Nature 1995 374 : 367–370
CAS PubMed Google Scholar
Muldoon LL и др. . Сравнение внутримозгового заражения и нарушения осмотического гематоэнцефалического барьера для доставки аденовируса, вируса герпеса и частиц оксида железа в нормальный мозг крысы Am J Pathol 1995 147 : 1840–1851
CAS PubMed PubMed Central Google Scholar
Nilaver G и др. .Доставка вируса герпеса и аденовируса к внутримозговым опухолям голых крыс после нарушения осмотического гематоэнцефалического барьера Proc Natl Acad Sci USA 1995 92 : 9829–9833
CAS PubMed Google Scholar
Райнов Н.Г. и др. . Селективное поглощение вирусных и монокристаллических частиц, доставленных внутриартериально к экспериментальным новообразованиям головного мозга Hum Gene Ther 1995 6 : 1543–1552
CAS PubMed Google Scholar
Райнов Н.Г. и др. .Долгосрочная выживаемость в модели опухоли головного мозга грызунов за счет усиленной брадикинином внутриартериальной доставки терапевтического вектора вируса простого герпеса Cancer Gene Ther 1998 5 : 158–162
CAS PubMed PubMed Central Google Scholar
Barnett FH и др. . Селективная доставка векторов вируса герпеса к экспериментальным опухолям головного мозга с использованием RMP-7 Cancer Gene Ther 1999 6 : 14–20
CAS PubMed Google Scholar
Inamura T, черный K.Брадикинин избирательно открывает гематологический барьер опухоли в экспериментальных опухолях головного мозга J Cereb Blood Flow Metab 1994 14 : 862–870
CAS PubMed Google Scholar
Миллер CR и др. . Дифференциальная восприимчивость первичных и установленных клеток глиомы человека к аденовирусной инфекции: нацеливание через рецептор эпидермального фактора роста обеспечивает перенос гена, не зависящий от волоконного рецептора Cancer Res 1998 58 : 5738–5748
CAS Google Scholar
Jin BK и др. .Продленная экспрессия гена in vivo , управляемая промотором тирозингидроксилазы в векторе ампликона дефектного вируса простого герпеса Hum Gene Ther 1996 7 : 2015–2024
CAS PubMed Google Scholar
Бреннер М. и др. . Промотор GFAP управляет астроцит-специфической экспрессией у трансгенных мышей J Neurosci 1994 14 : 1030–1037
CAS PubMed Google Scholar
Szklarczyk A, Kaczmarek L.Мозг как уникальная антисмысловая среда Antisense Nucleic Acid Drug Dev 1999 9 : 105–116
CAS PubMed Google Scholar
Цуй Дж., Сюй С, Лю П. Подавление экспрессии фактора роста нервов в постишемическом гиппокампе антисмысловым олигонуклеотидом c-fos J Neurosci 1999 19 : 1335–1344
CAS PubMed Google Scholar
Boado R, Tsukamoto H, Pardridge W.Доставка антисмысловых молекул в мозг для лечения болезни Альцгеймера и церебрального СПИДа J Pharm Sci 1998 87 : 1308–1315
CAS Google Scholar
Martres M и др. . Повышение и понижение экспрессии переносчика дофамина путем переноса плазмидной ДНК в головном мозге крысы Eur J Neurosci 1998 10 : 3607–3616
CAS PubMed Google Scholar
Dunnett S, Bjorklund A.Перспективы новых восстановительных и нейропротективных методов лечения болезни Паркинсона Nature 1999 399 : 32–39
Google Scholar
Во время М., Нэгеле Дж., О’Мэлли К., Геллер А. Долгосрочное восстановление поведения у паркинсонических крыс с помощью вектора HSV, экспрессирующего тирозингидроксилазу Science 1994 266 : 1399–1403
CAS PubMed PubMed Central Google Scholar
Во время MJ и др. .Экспрессия in vivo терапевтических человеческих генов для выработки дофамина в хвостатой части обезьян, получавших МРТР, с использованием вектора AAV. Генная терапия 1998 5 : 820–827
CAS PubMed Google Scholar
Хореллоу П. и др. . Прямой внутримозговый перенос гена аденовирусного вектора, экспрессирующего тирозингидроксилазу, на модели болезни Паркинсона на крысах Neuroreport 1994 6 : 49–53
CAS PubMed Google Scholar
Fitoussi N и др. .Обмен и метаболизм дофамина в полосатом теле крыс с паркинсонизмом, трансплантированных генетически модифицированными астроцитами человека Neuroscience 1998 85 : 405–413
CAS PubMed Google Scholar
Хореллоу П., Брундин П., Маллет Дж., Бьорклунд А. In vivo высвобождение допа и дофамина из генно-инженерных клеток, привитых к денервированному полосатому телу крысы Нейрон 1990 5 : 393–402
CAS PubMed Google Scholar
Lundberg C, Horellou P, Mallet J, Bjorklund A.Генерация допа-продуцирующих астроцитов ретровирусной трансдукцией гена тирозингидроксилазы человека — in vitro характеристика и in vivo эффекты в модели паркинсона крысы Exp Neurol 1996 139 : 39–53
CAS PubMed Google Scholar
Trejo F, Vergara P, Brenner M, Segovia J. Генная терапия на модели болезни Паркинсона на грызунах с использованием дифференцированных клеток C6, экспрессирующих трансген GFAP-тирозингидроксилазы Life Sci 1999 65 : 483–491
CAS PubMed Google Scholar
Вулф Дж., Фишер Л., Сюй Л.Прививка фибробластов, генетически модифицированных для продуцирования L-допа, на модели болезни Паркинсона на крысах Proc Natl Acad Sci USA 1989 86 : 9011–9014
Google Scholar
Канг У. и др. . Регулирование выработки дофамина генетически модифицированными первичными фибробластами J Neurosci 1993 13 : 5203–5211
CAS PubMed Google Scholar
Cao L и др. .Генная терапия модельной крысы с болезнью Паркинсона путем прямой инъекции плазмидной ДНК-липофектина Hum Gene Ther 1995 6 : 1497–1501
CAS PubMed Google Scholar
Имаока Т, Дата I, Омото Т, Нагацу Т. Значительное восстановление поведения в модели болезни Паркинсона путем прямого внутримозгового переноса гена с использованием непрерывной инъекции комплекса плазмидная ДНК-липосома Hum Gene Ther 1998 9 : 1093–1102
CAS PubMed Google Scholar
Сеговия Дж., Вергара П., Бреннер М.Специфическая для астроцитов экспрессия тирозингидроксилазы после внутримозгового переноса гена вызывает восстановление поведения при экспериментальном паркинсонизме Генная терапия 1998 5 : 1650–1655
CAS PubMed Google Scholar
Bencsics C и др. . Двойная трансдукция с помощью GTP циклогидролазы I и тирозингидроксилазы необходима для спонтанного синтеза L-допа первичными фибробластами J Neurosci 1996 16 : 4449–4456
CAS PubMed Google Scholar
Leff S и др. .Продукция L-допа in vivo генетически модифицированными первичными фибробластами крысы или трансплантатами клеток глиосаркомы 9L посредством коэкспрессии GTP циклогидролазы I с тирозингидроксилазой Exp Neurol 1998 151 : 249–264
CAS PubMed Google Scholar
Wachtel S, Bencsics C, Kang U. Роль ароматической декарбоксилазы L-аминокислот в замещении дофамина генетически модифицированными фибробластами в модели болезни Паркинсона на крысах J Neurochem 1997 69 : 2055–2063
CAS PubMed Google Scholar
Вентилятор D и др. .Восстановление поведения у крыс с поражением 6-гидроксидофамином путем контрансдукции полосатого тела генами тирозингидроксилазы и ароматической L-аминокислотной декарбоксилазы с использованием двух отдельных аденоассоциированных вирусных векторов Hum Gene Ther 1998 9 : 2527–2535
CAS PubMed Google Scholar
Ли У, Чанг Дж, Немет Н, Кан У. Везикулярный переносчик моноаминов-2 и декарбоксилаза ароматических L-аминокислот усиливают доставку дофамина после введения L-3,4-дигидроксифенилаланина крысам с паркинсонизмом J Neurosci 1999 19 : 3266–3274
CAS PubMed Google Scholar
Лефф С., Спратт С., Снайдер Р., Мандель Р.Долгосрочное восстановление активности декарбоксилазы L-ароматических аминокислот полосатого тела с использованием переноса гена рекомбинантного аденоассоциированного вирусного вектора на модели болезни Паркинсона на грызунах Neuroscience 1999 92 : 185–196
CAS PubMed Google Scholar
Исаксон О. Поведенческие эффекты и доставка генов на крысиной модели болезни Паркинсона; технические комментарии Наука 1995 269 : 856–857
CAS PubMed Google Scholar
Кирик Д., Розенблад С, Бьорклунд А.Характеристика поведенческих и нейродегенеративных изменений у крыс после частичных повреждений нигростриатальной дофаминовой системы, вызванных интрастриатальным введением 6-гидроксидофамина Exp Neurol 1998 152 : 259–277
CAS PubMed Google Scholar
Olsson M, Nikkhah G, Bentlage C, Bjorklund A. Акинезия передних конечностей в модели Паркинсона у крыс: дифференциальные эффекты агонистов дофамина и трансплантатов ниграла, оцененные с помощью нового шагового теста J Neurosci 1995 15 : 3863–3875
CAS PubMed Google Scholar
Чжоу Кью, Пальмитер Р.Мыши с дефицитом дофамина сильно гипокинетичны, адипсичны и афагичны. Cell 1995 83 : 1197–1209
CAS Google Scholar
Szczypka M et al . Доставка вирусного гена избирательно восстанавливает питание и предотвращает летальность мышей с дефицитом дофамина Neuron 199 22 : 167–178
CAS PubMed Google Scholar
Бон М.Комментарий к нейротрофическому фактору, производному линии глиальных клеток (GDNF). От молекулы, секретируемой глией, к генной терапии Biochem Pharmacol 1999 57 : 135–142
CAS PubMed Google Scholar
Choi-Lundberg DL и др. . Дофаминергические нейроны, защищенные от дегенерации с помощью генной терапии GDNF Science 1997 275 : 838–841
CAS PubMed Google Scholar
Лапчак П.А. и др. .Опосредованная аденовирусным вектором генная терапия GDNF в модели поражения грызунов на поздней стадии болезни Паркинсона Brain Res 1997 777 : 153–160
CAS PubMed Google Scholar
Мандель Р.Дж., Спратт СК, Снайдер РО, Лефф С.Е. Инъекция в средний мозг рекомбинантного аденоассоциированного вируса, кодирующего нейротрофический фактор линии глиальных клеток крыс, защищает нейроны черного мозга в модели прогрессирующей 6-гидроксидофаминовой дегенерации болезни Паркинсона у крыс Proc Natl Acad Sci USA 1997 9 : 14083–14088
Google Scholar
Galpern WR и др. .Клеточно-опосредованная доставка нейротрофического фактора головного мозга увеличивает уровни дофамина в модели дегенерации черной субстанции у крыс с MPP + Трансплантация клеток 1996 5 : 225–232
CAS PubMed Google Scholar
Левивье М., Прзедборски С., Бенчич С., Канг У. Интрастриатальная имплантация фибробластов, генетически сконструированных для производства нейротрофического фактора мозга, предотвращает дегенерацию дофаминергических нейронов в модели болезни Паркинсона у крыс J Neurosci 1995 15 : 7810–7829
CAS PubMed Google Scholar
Линднер М. и др. .Имплантация инкапсулированных клеток, продуцирующих катехоламин и GDNF, крысам с односторонним истощением дофамина и симптомами болезни Паркинсона Exp Neurol 1995 132 : 62–76
CAS PubMed Google Scholar
Yoshimoto Y и др. . Астроциты, ретровирусно трансдуцированные с помощью BDNF, улучшают поведение на крысиной модели болезни Паркинсона Brain Res 1995 691 : 25–36
CAS PubMed Google Scholar
Frim DM и др. .Имплантированные фибробласты, генетически сконструированные для производства нейротрофического фактора головного мозга, предотвращают токсичность 1-метил-4-фенилпиридиния для дофаминергических нейронов у крыс Proc Natl Acad Sci USA 1994 91 : 5104–5108
CAS PubMed Google Scholar
Ямада М. и др. . Экспрессия bcl-2 вируса простого герпеса, опосредованная вектором, предотвращает вызванную 6-гидроксидофамином дегенерацию нейронов черной субстанции in vivo Proc Natl Acad Sci USA 1999 96 : 4078-4083
CAS PubMed PubMed Central Google Scholar
Wexler N и др. .Гомозиготы по болезни Гентингтона Природа 1997 161 : 223–226
Google Scholar
Isacson O, Hague N. Генная терапия болезни Хантингтона. В: Breakefield XO (ed) Перенос генов и терапия неврологических расстройств The Humana Press: New Jersey 1988, стр. 423–440
Google Scholar
Спицы E. Нейрохимические изменения при хорее Хантингтона.Исследование ткани головного мозга после смерти Мозг 1980 103 : 179–210
CAS PubMed Google Scholar
Кордовер Дж., Исаксон О., Эмерих Д. Клеточная доставка трофических факторов для лечения болезни Хантингтона: возможна ли нейрозащита? Exp Neurol 1999 159 : 4–20
CAS PubMed Google Scholar
Schumacher J и др. .Внутримозговая имплантация фибробластов, продуцирующих фактор роста нервов, защищает полосатое тело от нейротоксических уровней возбуждающих аминокислот Neuroscience 1991 45 : 561–570
CAS PubMed Google Scholar
Kordower J и др. . Трансплантаты EGF-чувствительных нервных стволовых клеток, полученных от трансгенных мышей GFAP-gBGF: трофические и тропические эффекты в модели болезни Хантингтона на грызунах J Comp Neurol 1997 387 : 96–113
CAS PubMed Google Scholar
Emerich D et al .Защитный эффект инкапсулированных клеток, продуцирующих нейротрофический фактор CNTF, на модели обезьяны с болезнью Хантингтона Nature 1997 386 : 395–399
CAS PubMed Google Scholar
Она В. и др. . Ингибирование каспазы-1 замедляет прогрессирование заболевания на мышиной модели болезни Хантингтона. Природа 1999 399 : 263–267
CAS Google Scholar
Эндрюс Т., Брукс Д.Достижения в понимании ранней болезни Хантингтона с использованием методов функциональной визуализации ПЭТ и SPET Mol Med Today 1998 4 : 532–539
CAS PubMed Google Scholar
Кэмпбелл К., Олссон М., Бьорклунд А. Региональная инкорпорация и сайт-специфическая дифференцировка предшественников полосатого тела, трансплантированных в эмбриональный желудочек переднего мозга Neuron 1995 15 : 1259–1273
CAS PubMed Google Scholar
Olsson M и др. .Обширная миграция и иннервация мишеней предшественниками полосатого тела после трансплантации в полосатое тело новорожденного Neuroscience 1997 79 : 57–78
CAS PubMed Google Scholar
Bjorklund A et al . Функциональная способность трансплантатов полосатого тела на крысиной модели Хантингтона. В: Dunnett S, Bjorklund A (ред.). Функциональная трансплантация нейронов Raven Press: New York 1994 157–197
Isacson O et al .Функциональная замена нейронов привитыми нейронами полосатого тела в полосатом теле крысы, пораженном иботеновой кислотой Природа 1984 311 : 458–460
CAS PubMed Google Scholar
Tuszynski MH и др. . Генная терапия головного мозга взрослых приматов: интрапаренхиматозные трансплантаты клеток, генетически модифицированные для выработки фактора роста нервов, предотвращают холинергическую дегенерацию нейронов Генная терапия 1996 3 : 305–314
CAS PubMed Google Scholar
Тушинский и др. .Нацеленная внутрипаренхимная доставка человеческого NGF путем переноса гена в базальный передний мозг приматов в течение 3 месяцев не ускоряет отложение бета-амилоидных бляшек Exp Neurol 1998 154 : 573–582
CAS PubMed Google Scholar
Aebischer P и др. . Интратекальная доставка CNTF с использованием инкапсулированных генетически модифицированных ксеногенных клеток пациентам с боковым амиотрофическим склерозом Nature Med 1996 2 : 696–699
CAS PubMed Google Scholar
Haase G и др. .Терапевтический эффект цилиарного нейротрофического фактора при прогрессирующей моторной нейропатии зависит от пути доставки. Ann Neurol 1999 45 : 296–304
CAS PubMed Google Scholar
Gimenez y Ribotta M et al . Предотвращение гибели мотонейронов аденовирусными нейротрофическими факторами J Neurosci Res 1997 48 : 281–285
CAS PubMed Google Scholar
Mohajeri M, Figlewicz D, Bohn M.Внутримышечные трансплантаты миобластов, генетически модифицированные для секреции нейротрофического фактора линии глиальных клеток, предотвращают потерю мотонейронов и прогрессирование заболевания на мышиной модели семейного бокового амиотрофического склероза Hum Gene Ther 1999 10 : 1853–1866
CAS PubMed Google Scholar
Pechan P и др. . Генная терапия ишемического инсульта. В: Chiocca E, Breakefield XO (eds) Генная терапия неврологических заболеваний и опухолей головного мозга Human Press: Totowa, NJ 1998 397–407
Google Scholar
Линник М., Захос М., Гешвинд М., Федерофф Х.Экспрессия bcl-2 из дефектного вектора вируса простого герпеса-1 ограничивает гибель нейронов при очаговой церебральной ишемии Инсульт 1995 26 : 1670–1675
CAS PubMed Google Scholar
Лоуренс М.С. и др. . Вирусные векторы простого герпеса, экспрессирующие Bcl-2, являются нейропротективными при доставке после инсульта J Cereb Blood Flow Met 1997 17 : 740–744
CAS Google Scholar
Бец А, Ян Г, Дэвидсон Б.Уменьшение размера инсульта у крыс с использованием аденовирусного вектора для индукции сверхэкспрессии антагониста рецептора интерлейкина-1 в головном мозге J Cereb Blood Flow Met 1995 15 : 547–551
CAS Google Scholar
Ян Г. и др. . Ослабление ишемической воспалительной реакции в головном мозге мыши с использованием аденовирусного вектора для индукции сверхэкспрессии антагониста рецептора интерлейкина-1 J Cereb Blood Flow Met 1998 18 : 840–847
CAS Google Scholar
Сюй Д. и др. .Повышение нейрональной экспрессии NAIP снижает ишемическое повреждение гиппокампа крысы Nature Med 1997 3 : 997–1004
CAS PubMed Google Scholar
Yenari MA et al . Генная терапия HSP72 оказывает нейропротекторное действие на крысах с моделями инсульта и эпилепсии Ann Neurol 1988 44 : 584–591
Google Scholar
Мухонене М и др. .Перенос генов в сосуды головного мозга после субарахноидального кровоизлияния Инсульт 1997 28 : 822–828
Google Scholar
Pechan P и др. . Генетически модифицированные фибробласты, продуцирующие NGF, защищают нейроны гиппокампа после ишемии у крыс Neuroreport 1995 6 : 669–672
CAS PubMed Google Scholar
Крамм CM и др. .Генная терапия опухолей головного мозга Brain Pathol 1995 5 : 345–381
CAS PubMed PubMed Central Google Scholar
Weyerbrock A, Oldfield EH. Технологии переноса генов при злокачественных глиомах Curr Opin Oncol 1999 11 : 168–173
CAS PubMed Google Scholar
Moolten FL. Химиочувствительность опухоли, обусловленная встроенными генами тимидинкиназы герпеса: парадигма перспективной стратегии борьбы с раком Cancer Res 1986 56 : 5276–5281
Google Scholar
Mullen CA, Kilstrup M, Blaese RM.Перенос бактериального гена цитозиндезаминазы в клетки млекопитающих придает летальную чувствительность к 5-фторцитозину: система отрицательной селекции Proc Natl Acad Sci USA 1992 89 : 33–37
CAS PubMed PubMed Central Google Scholar
Aghi M и др. . Синергетические противораковые эффекты терапии гена ганцикловира / тимидинкиназы и 5-фторцитозин / цитозиндезаминазы J Natl Cancer Inst 1998 90 : 370–380
CAS Google Scholar
Wei MX и др. .Экспериментальная терапия опухолей у мышей с использованием активирующего циклофосфамид гена цитохрома P450 2B1 Hum Gene Ther 1994 5 : 969–978
CAS PubMed PubMed Central Google Scholar
Manome Y et al . Трансдукция вирусным вектором кДНК дезоксицитидинкиназы человека сенсибилизирует клетки глиомы к цитотоксическим эффектам цитозинарабинозида in vitro и in vivo Nature Med 1996 2 : 567–573
CAS PubMed Google Scholar
Maisonpierre PC и др. .Ангиопоэтин-2, естественный антагонист Tie2, который нарушает in vivo ангиогенез Science 1997 277 : 55–60
CAS PubMed PubMed Central Google Scholar
Milauer B et al . Рост глиобластомы ингибировал in vivo доминантно-отрицательным мутантом Flk-1 Nature 1994 367 : 576–578
Google Scholar
Паулюс В., Баур И., Бейтлер А.С., Ривз С.А.Для диффузной инвазии клеток глиомы в мозг необходимы интегрины бета-1 Lab Invest 1996 75 : 819–826
CAS Google Scholar
Weston BW и др. . Экспрессия антисмысловых последовательностей альфа (1,3) фукозилтрансферазы человека подавляет опосредованную селектином адгезию и метастазирование в печень клеток карциномы толстой кишки Cance Res 1999 59 : 2127–2135
CAS Google Scholar
Ким JYH и др. .опухолевые клетки, трансдуцированные макрофагным колониестимулирующим фактором Hum Gene Ther 1997 8 : 1065–1072
CAS PubMed PubMed Central Google Scholar
Herrlinger U et al . Вакцинация от экспериментальных глиом с использованием клеток глиомы, трансдуцированных GM-CSF Cancer Gene Ther 1997 4 : 345–352
CAS PubMed PubMed Central Google Scholar
Кикучи Т. и др. .Противоопухолевая активность опухолевых клеток, продуцирующих интерлейкин-2, и рекомбинантного интерлейкина 12 против клеток глиомы мыши, расположенных в центральной нервной системе Int J Cancer 1999 80 : 425–430
CAS PubMed Google Scholar
Bischoff JR и др. . Мутант аденовируса, который избирательно реплицируется в опухолевых клетках человека с дефицитом р53 Science 1996 274 : 373–376
CAS Google Scholar
Мартуза Р.Л. и др. .Экспериментальная терапия глиомы человека с помощью генно-инженерного мутанта вируса Science 1991 252 : 854–856
CAS PubMed PubMed Central Google Scholar
Walczak H и др. . Опухолевая активность лиганда, индуцирующего апоптоз, связанного с фактором некроза опухоли in vivo Nature Med 1999 5 : 157–163
CAS Google Scholar
Kondo S и др. .Таргетная терапия злокачественной глиомы человека на мышиной модели с помощью антисмысловой терапии 2-5A, направленной против теломеразной РНК Онкоген 1998 16 : 3323–3330
CAS Google Scholar
Сорочану Л., Гиллеспи Й, Хазаэли М.Б., Зонтхаймер Х. Использование хлортоксина для нацеливания на первичные опухоли головного мозга Cancer Res 1998 58 : 4871–4879
CAS PubMed Google Scholar
Бейтман А и др. .Фузогенные мембранные гликопротеины как новый класс терапевтических генов для генной терапии рака ASGT Meeting 1999 Abst. № 687
Birkenmeier EH и др. . Мукополисахаридоз мышей VII типа. Характеристика мышей с дефицитом бета-глюкуронидазы J Clin Invest 1989 83 : 1256–1258
Google Scholar
Wolfe JH, Deshmane SL, Fraser NW.Перенос гена вектора герпеса и экспрессия бета-глюкуронидазы в центральной нервной системе мышей MPS VII Nat Genet 1992 1 : 379–384
CAS PubMed Google Scholar
Ghodsi A и др. . Обширная активность бета-глюкуронидазы в центральной нервной системе мышей после переноса гена, опосредованного асденовирусом, в мозг Hum Gene Ther 1998 9 : 2331–2340
CAS PubMed Google Scholar
Watson GL и др. .Лечение лизосомного накопления в головном мозге мышей MPS VII, получавших интратекальное введение аденоассоциированного вируса Генная терапия 1998 5 : 1642–1649
CAS PubMed Google Scholar
Elliger S и др. . Устранение лизосомных накоплений в головном мозге мышей MPS VII, получавших интратекальное введение вектора аденоассоциированного вируса Генная терапия 1999 6 : 1175–1178
CAS PubMed Google Scholar
Finegold AA, Mannes AJ, Iadarola MJ.Паракринная парадигма in vivo генная терапия в центральной нервной системе: лечение хронической боли Hum Gene Ther 1999 10 : 1251–1257
CAS PubMed Google Scholar
Канг В. и др. . Опосредованный вирусом герпеса перенос гена препроэнкефалина в миндалину является антиноцицептивным Brain Res 1998 792 : 133–135
CAS PubMed Google Scholar
Wilson SP и др. .Антигипералгезические эффекты инфекции, вызванной вирусом простого герпеса, кодирующим препроэнкефалин Proc Natl Acad Sci USA 1999 96 : 3211–3216
CAS PubMed Google Scholar
McLaughlin ME, Ehrhart TL, Berson EL, Dryja TP. Спектр мутаций гена, кодирующего бета-субъединицу палочковой фосфодиэстеразы, у пациентов с аутосомно-рецессивным пигментным ретинитом Proc Natl Acad Sci USA 1995 92 : 3249–3253
CAS PubMed Google Scholar
Cayouette M, Gravel C.Опосредованный аденовирусом перенос гена цилиарного нейротрофического фактора может предотвратить дегенерацию фоторецепторов у мышей с дегенерацией сетчатки (rd) Hum Gene Ther 1997 8 : 423–430
CAS PubMed Google Scholar
Беннет Дж. и др. . Опосредованная аденовирусом доставка bcl-2, стимулированного родопсином, приводит к задержке гибели фоторецепторных клеток у мышей rd / rd Генная терапия 1998 5 : 1156–1164
CAS PubMed Google Scholar
Беннет Дж. и др. .Спасение фоторецепторных клеток у мышей с дегенерацией сетчатки (rd) с помощью in vivo генная терапия Nature Med 1996 2 : 649–654
CAS PubMed Google Scholar
Джомари К. и др. . Восстановление функции фоторецепторов с помощью AAV-опосредованного переноса гена на мышиной модели наследственной дегенерации сетчатки Генная терапия 1997 4 : 683–690
CAS PubMed PubMed Central Google Scholar
Кафри Т.Клеточный иммунный ответ на клетки, инфицированные аденовирусным вектором, не требует de novo экспрессия вирусного гена: значение для генной терапии Proc Natl Acad Sci USA 1998 95 : 11377–11382
CAS PubMed Google Scholar
Такахаши М., Миёси Х, Верма И.М., Гейдж ФХ. Спасение от дегенерации фоторецепторов у человека путем переноса генов вируса иммунодефицита человека, опосредованного вектором J Virol 1999 73 : 7812–7816
CAS PubMed PubMed Central Google Scholar
Ноебельс JL.Ориентация на гены эпилепсии Neuron 1996 16 : 241–244
CAS PubMed Google Scholar
Роберт Дж. Дж. и др. . Опосредованный аденовирусом перенос функционального гена GAD в нервные клетки: потенциал для лечения неврологических заболеваний Генная терапия 1997 4 : 1237–1245
CAS PubMed Google Scholar
Zhang LX и др. .Передача гена холецистокинина с помощью липофектина корректирует поведенческие аномалии крыс с аудиогенными припадками Neuroscience 1997 77 : 15–22
CAS PubMed Google Scholar
Geddes BJ, Harding TC, Lightman SL, Uney JB. Долгосрочная генная терапия в ЦНС: обращение вспять гипоталамического диабета у крыс Brattleboro с помощью аденовируса, экспрессирующего аргинин вазопрессин Nature Med 1997 3 : 1402–1405
CAS PubMed Google Scholar
Марголис ТП и др. .Пути экспрессии вирусных генов во время острой нейрональной инфекции HSV-1 Вирусология 1992 189 : 150–160
CAS PubMed Google Scholar
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Профилактика неврологических заболеваний в более позднем возрасте
Эта статья посвящена профилактике неврологических заболеваний в послеродовой период жизни и рассматривает как общественное здоровье, так и индивидуальные стратегии, которые использовались для первичной профилактики.Вторичная профилактика рассматривается при определенных состояниях, например, инсульте.
Для разработки первичных профилактических стратегий необходимы надежные эпидемиологические данные. Факторы, которые следует учитывать, — это определение случая, дизайн исследования и полученная уверенность, с которой может быть сделана интерпретация результатов. Для многих обсуждаемых неврологических состояний определение случая проблематично. Например, в случае инсульта в большинстве научных исследований используется клиническое определение. Следовательно, проблема скрытого инфаркта мозга не рассматривается.Технологические достижения, особенно в области визуализации, также потенциально могут привести к изменениям в диагностической проницательности. Для многих неврологических состояний есть подробные серии клинических случаев, но при оценке рисков большинство доказательств было основано на исследованиях случай-контроль с полученными ошибками.
В этом обзоре в первую очередь рассматривается инсульт, поскольку он является основной причиной неврологической смерти и инвалидности взрослых. При текущих знаниях факторов риска это состояние, при котором больше всего возможностей для первичной профилактики.Краткий обзор некоторых других основных неврологических заболеваний включен, чтобы проиллюстрировать области, в которых сообщалось или требуется работа. В этом обзоре представлена перспектива развитого мира.
Бремя неврологических заболеваний
Число смертей от неврологических заболеваний в Соединенном Королевстве снижается с детства до среднего возраста с примерно 10% у детей от 1 до 4 лет до 5% у лиц в возрасте от 25 до 34 лет, с небольшой долей в дальнейшем. Однако смертность от цереброваскулярных заболеваний увеличивается с 35 лет и составляет третью по частоте причина смерти среди всего населения.
Данные о смерти Управления переписей и обследований населения (OPCS) для 19931 детализируют случаи смерти, связанные с неврологическими расстройствами (таблица 1):
Таблица 1Смертей в Англии и Уэльсе 1993
Имеется мало данных о заболеваемости из рутинной статистики, но отчеты о заболеваемости из общей практики дают ключ к пониманию бремени симптомов (таблица 2) .2 В целом неврологические жалобы и жалобы со стороны органов чувств представляют четвертую наиболее частую причину появления, составляя 17% консультации.Это самый высокий показатель в возрастной группе до 5 лет и неуклонно увеличивается с возрастом. Только по неврологическим симптомам в 1991-1991 гг. Прошли консультации 1732/10 000 пациентов. Один процент консультаций общей практики приходится на мигрень.
Таблица 2Количество консультаций в общей практике 1995
Таблица 3 3 обобщены данные об инфекционных заболеваниях с неврологическими последствиями, о которых сообщалось в Службе статистики инфекционных заболеваний (CDC) OPCS в 1993 году в Англии и Уэльсе.
Таблица 3Уведомления об инфекционных заболеваниях
Заболевания сосудов: инсульт
Многие из факторов риска, выявленных для инсульта, также были связаны с ишемической болезнью сердца, которые были выявлены в обсервационных исследованиях, в основном с учетом исхода ишемической болезни сердца, таких как исследование множественных факторов риска и вмешательство4 и исследования Фрамингема.5 Существуют факторы риска, специфичные для инсульта, но, как правило, соответствующие доли риска не были точно оценены. В большинстве исследований рассматривались только факторы риска полного инсульта или ишемического инсульта, а относительные риски взвешиваются для описания факторов риска тромботических событий, которые, однако, составляют 76% инсультов. Снижение смертности от инсульта, хотя и обнадеживающее, остается в значительной степени необъяснимым, и недавние данные о заболеваемости не предполагают, что снижение продолжается.
КЛАССИФИКАЦИЯ ВИДОВ ФАКТОРОВ РИСКА (ТАБЛИЦА 4)
При рассмотрении факторов риска инсульта важно различать факторы, которые поддаются уменьшению, и те, которые нет. В первой группе будут представлены доказательства того, как эффективно снизить их распространенность. Обозначены широкие категории, в рамках которых можно рассматривать риск:
Таблица 4Факторы риска инсульта
Унаследованные биологические признаки
Сюда входят возраст, пол и генетическая предрасположенность.
Социальные характеристики
Сюда входят этническое происхождение с культурными особенностями и социально-экономическим статусом. Эти черты совпадают с поведенческими и биологическими факторами.
Факторы окружающей среды
Это такие факторы, как температура и другие климатические переменные, которые могут влиять на сезонные и географические вариации заболеваемости и смертности от инсульта.
Физиологические характеристики
К ним относятся артериальное давление, сывороточные факторы, такие как холестерин и глюкоза, и ожирение.
Поведенческие характеристики (таблица 5)
К ним относятся курение, употребление алкоголя, диетические факторы, а также использование оральных контрацептивов и заместительной гормональной терапии.
Таблица 5Поведенческие черты
Унаследованные биологические признаки
Возраст — И в Соединенных Штатах, и в Соединенном Королевстве риск инсульта удваивается каждое последующее десятилетие старше 55 лет. 7 Согласно демографическим прогнозам Соединенного Королевства, возрастное число людей будет возрастать и подвергаться повышенному риску инсульта.Мальмгрен и др. Однако по оценкам 8, это приведет к чистому увеличению только на 4% людей с умеренными или тяжелыми физическими недостатками. Показатели заболеваемости увеличиваются с 0,2 на 1000 населения в год в возрасте 45-54 лет до 10/1000 среди людей в возрасте старше 85,7 лет.
Пол — Фрамингемское исследование показало, что в целом заболеваемость мужчин увеличилась на 30%, что немного выше у лиц моложе 65 лет. В рамках Проекта по инсульту в Оксфордшире в Соединенном Королевстве вероятность того, что мужчина перенесет первый инсульт, была на 26% выше. чем у женщины.9
Влияние на плод и младенец — Некоторые группы приписывают неравенство в отношении здоровья, наблюдаемое в Великобритании, влиянию плода и младенца.10 Существуют разногласия по поводу ранних причин сердечно-сосудистых заболеваний.11 Инсульт был связан с более ранней материнской и неонатальной смертностью.12 Если этот сдвиг в парадигме происхождения сердечно-сосудистых и цереброваскулярных заболеваний верен, исследования и лечение внутриутробных заболеваний могут быть альтернативной стратегией деятельности по снижению факторов риска в среднем возрасте.Однако необходимы более подробные исследования этих влияний на факторы риска сердечно-сосудистых заболеваний и инсульта в детском и раннем взрослом возрасте.
Семейный анамнез — Семейный анамнез инсульта у любого родственника первой степени родства был независимым предиктором инсульта только у женщин из белой когорты среднего класса с относительным риском 2,3 после учета возраста, холестерина, артериального давления, курения сигарет и т. Д. и диабет.13 Welin et al 14 в шведском когортном исследовании мужчин среднего возраста показали, что у мужчин, чьи матери умерли от инсульта, риск инсульта увеличивался в три раза, а относительный риск для населения был оценен как 0.19. Исследования частоты инсульта среди близнецов показывают, что смертельные случаи от инсульта в 20 раз чаще, чем ожидалось, как у монозиготных, так и у дизиготных близнецов, что означает, что общая семейная среда является чрезвычайно сильным фактором риска инсульта15.
Бромберг и др. 16 подсчитали, что субарахноидальное кровотечение у родственников первой степени родства встречается почти в семь раз чаще, чем у родственников второй степени, и это первый отчет, в котором проводится различие между типом родственника и риском.Это означает, что пожизненный риск субарахноидального кровотечения составляет от 2% до 5% у родственников первой степени родства. Следовательно, скрининг на наличие неразорвавшейся аневризмы должен рассматриваться, по крайней мере, у родственников первой степени родства пациентов с субарахноидальным кровотечением.
Социальные характеристики
Социально-экономический статус (SEC) — в Соединенном Королевстве Ачесон и Сандерсон17 оценили относительный риск 1,6 в группе с самым низким (SEC V) по сравнению с группой с самым высоким (SEC 1). Также существует тенденция к тому, что SMR (стандартизованный коэффициент смертности) от инсульта будет выше в северных регионах Соединенного Королевства и в центральных районах города, где проживает доля SEC IV и V.18 Это может отражать множество факторов, таких как социальная депривация, экологические и географические различия, а также различия в питании. Британское региональное исследование сердца не обнаружило существенной разницы в риске инсульта между социально-экономическими группами после поправки на другие факторы риска19.
Этническая группа — Данные о смертности в Соединенных Штатах неизменно показывают более высокий уровень смертности от инсульта среди чернокожих сообществ, вероятно, отчасти из-за более высокой распространенности ишемической болезни сердца и гипертонии в этих сообществах.20-24 В США Kittner et al 25 документально подтвердили, что относительный риск инсульта составляет 1,4 для чернокожих женщин и 1,1 для чернокожих мужчин после поправки на возраст, гипертонию и сахарный диабет. Было высказано предположение о возможности того, что конкретный тип инсульта является причиной расовых различий26. Даже среди чернокожих сообществ существует связь между более темным цветом кожи и гипертонией в группах с более низким социально-экономическим статусом27.
Балараджан28 показал значительно различающиеся уровни смертности между этническими группами в Англии и Уэльсе.В целом, этнические группы, в которых смертность от ишемической болезни сердца в 1970-х годах была ниже, показали наибольшее улучшение в течение следующего десятилетия. Самые высокие показатели ишемической болезни сердца были обнаружены у лиц индийского происхождения. У рожденных в Карибском бассейне, Соединенных Штатах, старом Содружестве и Западной Европе были самые низкие показатели. Самые высокие показатели инсульта были у афрокарибских бобов, за которыми следовали африканцы, индейцы и ирландцы, и они показали снижение на 28% с 1970-2 по 1979-83 гг. Однако в этом анализе были проблемы с классификацией, и не было сделано никаких поправок на социальный класс или другие факторы.Эти данные относятся только к стране происхождения, а не к этнической группе, и поэтому не принимают во внимание этнические группы второго поколения. Самый высокий уровень смертности от инсульта у индийских мужчин, что на 53% выше среднего по стране, и на 25% у индийских женщин. Эти данные подтверждают необходимость установления распространенности более традиционных факторов риска инсульта в больших репрезентативных выборках этих сообществ. Круикшенк и др. сообщили о 76% превышении смертности среди карибских мужчин.29 Эти различия в смертности никогда не находили удовлетворительного объяснения.29 30
Исследование Чатурведи и др. 31 обнаружил, что медиана систолического артериального давления у мужчин афрокарибского региона на 6 мм рт. Ст. Выше, чем у мужчин из Европы, и на 17 мм рт. Ст. У женщин из афро-карибских стран, чем у европейских женщин. Haines и др. 32, однако, не обнаружили существенной разницы между уровнями артериального давления у белых и чернокожих испытуемых, посещающих центральный район Лондона, общей практики, но обнаружили, что больше чернокожих, чем белых людей лечили от гипертонии.
Сама по себе гипертония не может объяснить высокую частоту инсультов у чернокожих людей. Еще одним важным соображением является разница в распространенности сахарного диабета между этническими группами. Среди афрокарибов выше показатели смертности и госпитализации, чем среди других этнических групп.29 33 Миллер и др. 34 обследовали 209 мужчин (81% всех зарегистрированных в общей практике) и обнаружили, что гипергликемия присутствует у 29% афрокарибских бобов по сравнению с 9% европейцев.Гейнс и др. 24 обзора литературы о различиях между черным и белым в факторах риска инсульта пришли к выводу, что помимо гипертонии, курение, диабет и ожирение более распространены среди чернокожих.
В США, Говард и др. 35 подсчитали, что на социально-экономический статус приходилось менее 25% избыточной смертности мужчин от инсульта среди чернокожих людей в возрасте 45-64 лет, а среди чернокожих женщин он не значительно снижал оценочную избыточную смертность от инсульта.
Факторы окружающей среды
Многие группы сообщают о сезонных колебаниях смертности от инсульта.
Возможные механизмы могут быть связаны с влиянием температуры на кровяное давление и механизмы свертывания крови, и необходимы дальнейшие исследования среды, в которой произошел инсульт, в связи с кровяным давлением и гемостатическими факторами.
Физиологические характеристики
Гипертония: данные наблюдений и экологические данные — Все исследования показали, что артериальная гипертензия является единственным наиболее важным фактором риска инсульта, который, как было показано, в значительной степени снижается с помощью антигипертензивной терапии в условиях рандомизированного контролируемого исследования.Приписываемый фактор риска, однако, варьировался от 0,6 в исследовании Framingham40 до 0,75 в исследовании Whitehall41-43.
Мировые тенденции в области гипертонии и ее сердечно-сосудистых последствий имеют большое значение для общественного здравоохранения. Снижение смертности как от инсульта, так и от ишемической болезни сердца предшествовало эффективной антигипертензивной терапии, и остаются неясными причины вклада антигипертензивной терапии в это снижение (44).
Bonita и Beaglehole45 предположили, что по крайней мере три четверти снижения смертности от инсульта в Соединенных Штатах в период 1970-80 гг. Было вызвано другими факторами, а не антигипертензивным лечением.
Данные вмешательства в связи с гипертонией: метаанализ. — МакМахон и его коллеги46 провели метаанализ девяти основных проспективных обсервационных исследований с участием субъектов в возрасте 25–84 лет, 98% из которых — мужчины. После поправки на погрешность регрессионного разбавления продолжительные различия в обычном диастолическом артериальном давлении в 5, 7,5 и 10 мм рт. при обычном артериальном или нормотензивном артериальном давлении более низкое артериальное давление в конечном итоге должно снижать риск сосудистых заболеваний.Этот эффект наблюдался во всем диапазоне артериального давления, и не было никаких доказательств того, что какой-либо порог, ниже которого уровни диастолического артериального давления не были связаны с более низким риском инсульта. Следовательно, для групп населения, у которых часто встречаются инсульты, существенно более низкое диастолическое артериальное давление в конечном итоге должно приводить к существенно меньшему относительному риску сосудистых заболеваний. Абсолютные преимущества более низкого кровяного давления будут наибольшими для крупных стран с высокой частотой инсультов, таких как Китай.Они пришли к выводу, что все еще существует необходимость в лучшем понимании диетических и других детерминант уровней артериального давления у населения.
Коллинз и др. 47 проанализировали 14 рандомизированных исследований гипотензивных препаратов. Долгосрочные различия, наблюдаемые в обсервационных исследованиях, в пределах 5-6 мм рт.ст. в обычном диастолическом артериальном давлении были связаны с примерно на 35-40% меньшим количеством случаев инсульта. Число инсультов снизилось в испытаниях на 42 (стандартное отклонение 6)%, что позволяет предположить, что практически все эпидемиологически ожидаемое снижение инсульта происходит быстро.В отношении инсульта обзор предоставляет прямые и весьма важные доказательства того, что как смертельный, так и нефатальный инсульт можно предотвратить всего за несколько лет после снижения артериального давления, даже если диастолическое артериальное давление ниже 110 мм рт. Ст. В начале лечения. Пропорционально схожее снижение наблюдалось среди пациентов с легкой, умеренной и тяжелой артериальной гипертензией.47 Это говорит о том, что в рамках программы первичной медико-санитарной помощи пациенты с пограничным артериальным давлением 140-160 мм рт. быть включены для рассмотрения подходов к снижению артериального давления.
Болезни сердца — Считается, что после возраста, пола и гипертонии наиболее важным фактором риска инсульта является сердечное заболевание. В популяции пациентов с гипертонической болезнью Фрамингема соотношение инфаркта миокарда и инсульта составляло два у мужчин и один у женщин, тогда как оно составляло шесть у мужчин и три у женщин, не страдающих гипертонией.5 Исследование также показало у мужчин, что при наличии коронарной болезни болезни сердца: относительный риск инсульта составил 2,5 при наличии застойной сердечной недостаточности и 4.4 при гипертрофии левого желудочка. Риски были более выражены у женщин (3,2, 4,3 и 7,9 соответственно). Исследование округа Эванс26 нашло аналогичные, но более слабые доказательства этого. Напротив, Велин и его коллеги14 при многофакторном анализе не обнаружили, что ишемическая болезнь сердца или гипертрофия левого желудочка являются значимыми факторами.
Гипертрофия левого желудочка — Десять процентов людей в возрасте 30-62 лет могут ожидать изменений ЭКГ при гипертрофии левого желудочка в течение 10 лет48 между 15-57% и эхокардиографии до 98%.49 Гипертрофия левого желудочка на ЭКГ указала на высокий риск инсульта в исследовании Framingham (относительный риск 7,5 у мужчин и 8,6 у женщин). В некоторой степени это отражает тяжесть и продолжительность ассоциированной гипертензии, но остаточный эффект гипертрофии левого желудочка, показанный на ЭКГ, является значительным (от четырех до восьми раз) после корректировки на сопутствующее повышение артериального давления.48
Фибрилляция предсердий — Фибрилляция предсердий поражает 2% населения (0.От 5% в возрасте от 50 до 59 лет до 8,8% в возрасте от 80 до 89 лет), а в исследовании Framingham Study было установлено, что на фибрилляцию предсердий приходится от 7% до 31% всех инсультов у пациентов в возрасте до 60 лет, что в несколько раз увеличивает риск инсульта. из пяти (1,5% в пятом десятилетии до 23,5% в восьмом десятилетии) 50. 51 До 20% лиц с фибрилляцией предсердий, у которых был первый инсульт, имеют следующий эпизод в течение одного года52. Кроме того, фибрилляция предсердий связана с более высокой ранней смертностью у пациентов, госпитализированных с острым инсультом53, и с большим риском рецидива. инсульт54, хотя эти ассоциации не были обнаружены во Фрамингемском исследовании.55 Копецки и др. 56 предположили, что одиночная фибрилляция предсердий у пациентов младше 60 лет на момент постановки диагноза не была связана с повышенным риском инсульта, но когортное исследование не включало пациентов с сердечно-сосудистыми заболеваниями, гипертонией или диабетом в начале. У людей с фибрилляцией предсердий без ревматической болезни сердца вероятность первого инсульта или транзиторной ишемической атаки в пять-шесть раз выше, чем в контрольной группе того же возраста и артериального давления50; у лиц с фибрилляцией предсердий и ревматической болезнью сердца риск в 17 раз выше, чем в контрольной группе.Gustafsson и др. , 57 в экологическом анализе в Швеции предположили, что лечение антикоагулянтами или аспирином является экономически эффективным при условии, что риск серьезных осложнений кровотечения остается низким (экономия 2 миллиона фунтов стерлингов на миллион жителей в год).
Доказательства испытаний — Неревматическая фибрилляция предсердий увеличивает риск инсульта, и пять рандомизированных контролируемых испытаний использования варфарина для первичной профилактики инсульта показали последовательное снижение примерно на две трети при умеренной длительной антикоагуляции.58-64 Варфарин снижает частоту ишемического инсульта или эмболии с 4,5% до 1,4% в год (31 случай на 1000 пролеченных). Только в двух из этих исследований также сообщалось о положительном эффекте аспирина у пациентов с фибрилляцией предсердий. 65 В исследовании «Профилактика инсульта при фибрилляции предсердий» сообщается, что количество первичных событий или смертей снижалось на 58% (P = 0,01) варфарином и на 42% — аспирином (P = 0,02). Риск значительного кровотечения составлял 1,5%, 1,4% и 1,6% в год у пациентов, получавших варфарин, аспирин и плацебо соответственно.В этом исследовании применялись очень строгие критерии исключения для включения в группу варфарина, а мониторинг был чрезвычайно тщательным, что отражалось в низком риске кровотечения. В условиях отсутствия испытаний, когда критерии не всегда соблюдаются, преимущества будут уменьшены.
В исследовании SPAF II66, хотя варфарин казался более эффективным, чем аспирин, для предотвращения ишемических инсультов, этот агент был связан с большим количеством геморрагических инсультов, которые, как правило, в большей степени приводят к инвалидности, чем ишемические инсульты.У пациентов младше 75 лет, у которых в анамнезе не было артериальной гипертензии, сердечной недостаточности или предшествующей тромбоэмболии, абсолютный риск ишемического инсульта или эмболии аспирином составлял 0,5% в год. Ранее рекомендовали варфарин, если не было противопоказаний для антикоагуляции. В свете результатов SPAF II лечение аспирином теперь можно рекомендовать многим пациентам из-за его эффективности, безопасности, стоимости и простоты применения. Варфарин показан пациентам с высоким риском, таким как пациенты, перенесшие преходящую ишемическую атаку или инсульт.
Хотя у лиц старше 75 лет самый высокий риск инсульта из-за фибрилляции предсердий, они подвергаются более высокому риску кровотечения при приеме антикоагулянтов. Эти испытания необходимо повторить в учреждениях первичной медико-санитарной помощи, если предполагается, что лечение будет основано на первичной помощи. Также необходимы дальнейшие исследования преимуществ и рисков различных комбинаций доз аспирина и варфарина.
Европейская группа исследования фибрилляции предсердий67 пришла к выводу, что антикоагулянты эффективны в снижении риска повторных сосудистых событий у пациентов с неревматической фибрилляцией предсердий с недавней транзиторной ишемической атакой или легким ишемическим инсультом.В абсолютном выражении можно предотвратить 90 сосудистых событий (в основном, инсульт), если 1000 пациентов будут лечиться антикоагулянтами в течение одного года. Аспирин предотвращает 40 таких смертей. Эта же группа проанализировала оптимальную интенсивность антикоагуляции.68
Копенгагенское исследование фибрилляции предсердий с аспирином69 показало 12% -ное снижение серьезных сосудистых событий (инсульт, инфаркт миокарда или сосудистая смерть), а обзор 25 исследований Antiplatelet Trialists Collaborations 70 сообщил о 25% -ном снижении всех типов сосудистой смерти при терапии аспирином.Большинство испытаний, проведенных на сегодняшний день, оценивающих пользу аспирина у этих пациентов, дали результаты, которые не достигли значимости, и для определения пользы требуются более крупные испытания. Исследование SPAF II продолжило рандомизацию пациентов между аспирином и варфарином, и результаты будут способствовать этому обсуждению66.
Регистры инсульта по месту жительства оценивают распространенность сердечной эмболии у пациентов, впервые перенесших инсульт, между 8% -20% и ишемической болезнью сердца между 13% -38%.71 Таким образом, возможности вторичной профилактики значительны.
Предыдущая транзиторная ишемическая атака — Whisnant40 оценил связанный с ней риск предыдущей транзиторной ишемической атаки в 10%, а оценки 13-кратного увеличения риска инсульта для субъектов, перенесших транзиторную ишемическую атаку, были обнаружены в первый год после инсульт и семикратный риск в последующие семь лет. Восемьдесят процентов транзиторных ишемических атак происходят из-за эмболов в области сонной артерии.Риск фибрилляции предсердий увеличивается с возрастом, но его общая распространенность составляет 0,5–2%. Около 20% пациентов с первым инсультом перенесли одну или несколько преходящих ишемических атак. Риск инсульта после транзиторной ишемической атаки оценивается в 5% в год, что соответствует 13-кратному увеличению риска инсульта72.
Стеноз сонной артерии — В нескольких крупных испытаниях сообщалось о промежуточных или окончательных результатах оценки преимуществ каротидной эндартерэктомии и определения уровня периоперационного риска, при котором процедура становится приемлемой: Европейское исследование хирургии сонной артерии (ECST), 73 Североамериканское исследование симптоматики Исследование каротидной эндартерэктомии (NASCET) 74 и Программа совместных исследований ветеранов 75, опубликованные в 1991 г., с аналогичными результатами.В первые два испытания были включены пациенты с незначительным ишемическим событием в анамнезе и тяжелым стенозом по происхождению ипсилатеральной внутренней сонной артерии (70–99%). Риск инсульта или смерти в течение 30 дней после операции составил 7,5% по шкале ECST и 5,8% по шкале NASCET. Если пациент выживает без инсульта, риск инсульта на ипсилатеральной территории в течение следующих нескольких лет, оцененный анализом таблицы смертности, составил 2,8% для хирургических пациентов и 16,8% для контрольной группы через три года в исследовании ECST и 9% для хирургических. пациентов и 20% для контрольной группы в исследовании NASCET.Благоприятный эффект операции таков, что от пяти до десяти пациентов необходимо прооперировать, чтобы предотвратить инсульт. Только половина предотвращенных инсультов приводит к отключению. Если бы все пациенты с тяжелым стенозом сонной артерии, у которых была транзиторная ишемическая атака, перенесли бы операцию, общее снижение общей частоты инсульта составило бы от 0,2% до 0,5%, но это привело бы к пятикратному увеличению количества операций на сонной артерии76. 77 Эндартерэктомия не показана большинству, а возможно, и всем пациентам с умеренным симптоматическим стенозом сонной артерии.78
Отсутствие убедительных доказательств в пользу эндартерэктомии при бессимптомном заболевании привело к созданию двух многоцентровых исследований: бессимптомного исследования артеросклеротических артерий сонных артерий (ACAS) и его европейского аналога — исследования бессимптомной хирургии сонных артерий (ACST). Недавние данные ACAS показывают, что если стеноз больше 60%, эндартерэктомия приносит пользу со снижением относительного риска на 55%, но со снижением абсолютного риска на 5,9% .79 Проблема исследования заключается в том, что 9% исследуемой группы не получали назначенные лечение и среднее время наблюдения были короткими.80
Вторичная профилактика после транзиторной ишемической атаки или ишемического инсульта — Сотрудничество Antiplatelet Trialists Collaboration провело метаанализ испытаний, оценивающих пользу терапии аспирином во вторичной профилактике инсульта, и оценило, что длительная антикоагулянтная терапия снижает риск инсульта и других крупных сосудов. событий примерно на 25% .81 Это преимущество было четко продемонстрировано для доз аспирина от 300 мг до 1300 мг / день, но остается неясным, будут ли низкие дозы аспирина эффективны для определенных групп высокого риска.
Было проведено два недавних клинических испытания, подтверждающих пользу более низких доз аспирина. В голландском исследовании транзиторной ишемической атаки82 сравнивали 30 мг аспирина с 300 мг, и в первой группе инсульт миокарда или смерть от сосудов произошли в 14,7% по сравнению с 15,2% в группе 300 мг. Однако событий было слишком мало, чтобы разница достигла значения. Второе исследование, Шведское исследование низких доз аспирина, 83 было первым, продемонстрировавшим явный положительный эффект аспирина в суточных дозах менее 300 мг.Эта группа сравнила эффект 75 мг аспирина с плацебо и показала значительное снижение риска на 17% в группе лечения по сравнению с плацебо.
Результаты Исследовательской группы по транзиторной ишемической атаке в Соединенном Королевстве84 также предполагают аналогичный положительный эффект для различных доз аспирина выше 300 мг, но побочные эффекты, вероятно, зависят от дозы. Благоприятный эффект аспирина, по-видимому, одинаков у людей с гипертонией и диабетом, во всех возрастных группах и у обоих полов.
Тиклопидин — еще одно антитромбоцитарное средство, которое, как было показано, эффективно в качестве вторичного профилактического средства с аналогичной, если не большей эффективностью, чем аспирин, но которое связано со значительно большим количеством побочных эффектов. Рекомендуется тем, кто плохо переносит аспирин. Интересно, что его механизм действия отличается от аспирина, и есть некоторые свидетельства того, что комбинированная терапия аспирином и тиклопидином дает больший положительный эффект, чем каждый в отдельности, но со значительно увеличенным кровотечением.85-87 Было показано, что родственный тиенопиридиновый препарат (клопидогрель) имеет меньше побочных эффектов, но дает мало преимуществ по сравнению с аспирином.
Что касается вторичной профилактики после транзиторной ишемической атаки, важно отметить, что только 12% инсультов предшествуют транзиторной ишемической атаке, и только о половине из них сообщается врачу. Таким образом, если на этом этапе вводится вторичная профилактика, это может предотвратить не более 5% всех инсультов.
Сотрудничество антиагрегантов70 провело обзор рандомизированных испытаний длительной антиагрегантной терапии.Они пришли к выводу, что пока нет четких доказательств баланса рисков и преимуществ антитромбоцитарной терапии в первичной профилактике среди субъектов с низким риском. Среди гораздо более широкого круга пациентов с высоким риском окклюзионного сосудистого заболевания, чем обычно лечится в настоящее время (например, нестабильная стенокардия, подозрение на острый инфаркт миокарда, инсульт, транзиторная ишемическая атака и другие группы высокого риска, такие как пациенты со стабильной стенокардией или периферической стенокардией). сосудистые заболевания), несколько лет антиагрегантной терапии с 75-325 мг аспирина в день или некоторыми другими антиагрегантными режимами обеспечивает эффективную защиту от инфаркта миокарда, инсульта и смерти.
Та же группа89 указывает, что антитромбоцитарная терапия, в основном аспирин в отдельности или аспирин плюс дипиридамол, значительно снижает риск окклюзии сосудов у широкого круга пациентов с высоким риском этого осложнения, но когда лечение следует начинать и как долго требуются дальнейшие исследования .
Соблюдение режима приема варфарина и его безопасность менее вероятны при назначении варфарина в общей практике, чем в выбранных исследованиях.90
Ожирение — Во многих исследованиях не удается отличить ожирение от связанного с ним признака — гипертонии, и неясно, насколько независимое влияние ожирение оказывает на частоту инсульта.Фрамингемское исследование предоставляет некоторые доказательства того, что ожирение является независимым фактором5; однако большая часть эффекта ожирения опосредована другими факторами риска. Ларссон и др. 91 ясно показал, что увеличение соотношения талии и бедер было связано с инсультом, но не было независимым долгосрочным предиктором.
Сахарный диабет — Фрамингемское исследование задокументировало относительный риск инсульта 2,6 у мужчин и 3,1 у женщин с диабетом и относительный риск около 10%.55 Совсем недавно Стегмайр и Асплунд 92, используя данные реестра населения, оценили риск в 4,1 для мужчин и 5,8 для женщин. По их оценкам, относимый риск составлял 18% у мужчин и 22% у женщин, а 50 инсультов ежегодно напрямую связаны с диабетом на 100000 населения, хотя неясно, как авторы контролировали другие факторы, такие как гипертония и фибрилляция предсердий.
Пациенты с диабетом с протеинурией или ишемическими отклонениями ЭКГ подвергаются наибольшему риску нового цереброваскулярного заболевания.93
Инфекция —Syrjanen et al. , 94 в исследовании «случай-контроль» подсчитали, что фебрильная инфекция в предыдущем месяце после инсульта у пациентов в возрасте до 50 лет предполагала относительный риск, равный девяти (95% доверительный интервал 2,2–80). Наиболее частая инфекция была респираторной (80%), с относительным риском 14,5 (95% ДИ 1,9–112,3), оцененным в анализе условной логистической регрессии для согласованных пар. В исследовании использовались средства контроля со стороны сообщества. Исследование случай-контроль, опять же с использованием контроля со стороны сообщества, Грау и др. 95 показали, что недавняя инфекция, в основном бактериального происхождения, может быть фактором риска цереброваскулярной ишемии как у пожилых, так и у молодых пациентов.
Гомоцистеинурия — Преждевременный артериосклероз и тромбоэмболические явления являются хорошо известными осложнениями гомозиготной гомоцистеинурии из-за дефицита синтеза цистатионина, который повреждает эндотелиальные клетки сосудов. Гетерозиготность по гомоцистеинурии может быть оценена после введения метионина, и считается, что она встречается у каждого 70 населения и предрасполагает к преждевременному окклюзионному заболеванию артерий, включая церебральное заболевание96 Perry at al 97 сообщили в Британском региональном исследовании сердца, что общие концентрации гомоцистеина являются сильным независимым фактором риска инсульта с постепенным увеличением относительного риска инсульта во втором, третьем и четвертом квартилях общего распределения гомоцистеина (отношение шансов 1.3-2.8). Найгард и др. 98 в обзоре сообщества в западной Норвегии показали, что повышенные концентрации общего гомоцистеина в плазме были связаны с основными компонентами профиля риска сердечно-сосудистых заболеваний, а именно с мужским полом, старостью, курением, высоким кровяным давлением, повышенным уровнем холестерина и отсутствием упражнение.
Серповидно-клеточная анемия — Сержант99 подсчитал, что у пациента с серповидно-клеточной анемией риск инсульта повышается примерно в 10 раз по сравнению с населением в целом.Риск наиболее высок для субъектов с гомозиготным HbSS, но гетерозиготные носители также несут значительный риск.
Фибриноген — Повышение концентрации фибриногена увеличивает вязкость крови, что может еще больше повысить риск образования тромба. Вильгельмсен и др. 100 продемонстрировали значительную связь с инсультом, когда систолическое артериальное давление было выше 200 мм рт. -3 г / л.Фрамингемское исследование5 показало значимую независимую связь фибриногена с инсультом, но коэффициент не был значимым. Велин и др. 14 оценили, что относимый риск популяции составляет 0,18, а в многофакторном анализе было показано, что фибриноген является независимым фактором риска. Qizilbash и др. , 101 в исследовании случай-контроль с поправкой на другие переменные оценили отношение шансов для ишемического инсульта как 1,78 (0,91–3,18) для фибриногена более 3,6 г / л.
Факторы свертывания — В исследовании Northwick Park Study Мид и его коллеги102 показали, что повышенная концентрация фактора VII является самым сильным предиктором инфаркта миокарда за пять лет до события, за которым следуют повышенные концентрации холестерина и фибриногена.
В последнее время также обратили внимание на антифосфолипидные антитела, такие как антикардиолипиновые антитела и волчаночный антикоагулянт. Они могут быть связаны с различными системными заболеваниями, но чаще всего с системной красной волчанкой. В некоторых случаях они являются изолированной находкой без каких-либо клинических корреляторов заболевания соединительной ткани.103 Эти антитела связаны с повышенным риском тромботических эпизодов как в венозных, так и в артериальных сосудах. В исследовании 100 последовательных пациентов, поступивших в больницу с первым в истории инсультом, Chakravarty et al 104 обнаружили, что 21% были положительными на антикардиолипиновые антитела по сравнению с никем из 100 контрольных групп соответствующего возраста.В группе пациентов с положительным результатом на антикардиолипиновые антитела летальность за три месяца составила 62% по сравнению с 21,5% в группе отрицательных антител. Кроме того, доля пациентов со значительной инвалидностью после инсульта (оценка по шкале Бартеля 0–9) была значительно выше в группе антител. Авторы предположили, что наличие антикардиолипиновых антител может представлять собой независимый прогностический маркер смертности и исхода от инсульта. Исследования проводились в отдельных группах, и требуется долгосрочное наблюдение за пациентами, чтобы оценить, является ли присутствие антител эпифеноменом.
Объем упакованных клеток — Повышенное кровяное давление и курение сигарет увеличивают объем упакованных клеток и составляют большую часть ассоциации с инсультом. Фрамингемское исследование показало значимую независимую корреляцию только у лиц моложе 65 лет.5 Велин и др. 14 не смогли показать ассоциацию. Иёхара и др. 105 обнаружили, что низкий объем упакованных клеток важен как независимый фактор риска у женщин.
Поведенческие характеристики
Физическая активность — В Британском региональном исследовании сердца физическая активность была обратно пропорциональна риску инсульта, независимо от факторов риска коронарных заболеваний, чрезмерного употребления алкоголя и ранее существовавшей ишемической болезни сердца или инсульта (относительный риск 1 для бездействия, 0.6 умеренная активность и 0,3 высокой активности) .106 Также в исследовании Framingham через 14 лет наблюдения наблюдалась аналогичная обратная связь между физической активностью и риском инсульта107, но это не было значимым после поправки на возраст и классификации физического состояния. активность была широкой.
Липиды — История гораздо менее ясна и последовательна, чем для ишемической болезни сердца. В исследовании Framingham была выявлена отрицательная связь холестерина липопротеинов низкой плотности с инсультом, особенно у женщин старше 65 лет после учета других факторов риска.Не было четкой связи с триглицеридами и защитного действия липопротеинов высокой плотности. 55
В метаанализе 10 проспективных исследований Qizilbash et al 101 108 описал объединенный относительный риск 2,9 (95% ДИ 1,43–5,87) для людей с общим холестерином более 5,7 ммоль / л по сравнению с более низкими концентрациями. У белых людей риск инсульта из-за концентрации холестерина выше 5,7 ммоль / л был рассчитан как 0.2. В исследовании случай-контроль та же группа обнаружила, что фибриноген и липиды являются важными факторами риска ишемического инсульта. С поправкой на другие переменные отношения шансов ишемических событий составили 1,73 (0,9–3,29) для общего холестерина> 6 ммоль / л; 1,34 (0,69–2,61) для липопротеидов низкой плотности> 3,5 ммоль / л; и 0,32 (0,15-0,69) для липопротеидов высокой плотности> 1,2 ммоль / л.109
На шестилетнем этапе наблюдения в исследовании группы исследований по вмешательству множественных факторов риска (MRFIT), Iso et al 110 показали, что снижение концентрации холестерина в сыворотке крови было связано с трехкратным увеличением риска внутричерепного кровотечения после учета возраста, курения, диастолического артериального давления и расы или этнической группы.Группа подчеркнула, что повышенный риск был почти исключительно среди мужчин с высоким кровяным давлением и что на внутричерепные кровотечения приходилось только 2,9% всех смертей от цереброваскулярных заболеваний. Между высоким уровнем холестерина в сыворотке крови и смертью от негеморрагического инсульта и общего сердечно-сосудистого заболевания была выявлена подавляющая положительная связь.
Курение сигарет — Мета-анализ связи между курением сигарет и инсультом предоставил убедительные доказательства повышенного риска инсульта среди курильщиков с общим относительным риском, равным 1.5 (1,4-1,6) .111
Алкоголь —Алкоголь является фактором риска гипертонии, и исследования показали положительную связь между употреблением алкоголя и инсультом, которая может быть связана с гипертонией, а не с алкоголем, действующим независимо.112 Есть данные, что хроническое и чрезмерное употребление алкоголя (> 60 г этанола / день) связаны с повышенным риском всех типов инсульта.112 Роль низкого или умеренного потребления алкоголя остается неясной.
Данные более чем 19 проспективных когортных исследований не предоставляют убедительных доказательств какой-либо связи с общим инсультом, за исключением явно умеренного увеличения риска при самом высоком уровне потребления алкоголя.Когда данные разделены по двум широким категориям типа инсульта, выявляются согласованные закономерности. Для геморрагического инсульта есть убедительные доказательства связи, обычно с трехкратным повышением риска для более высоких уровней потребления алкоголя. Напротив, существуют столь же убедительные доказательства против любой существенной неблагоприятной связи с ишемическим инсультом.
Калий — В исследовании INTERSALT117 калий отрицательно влияет на кровяное давление.Khaw и др. 118 оценил относительный риск инсульта в калифорнийской когорте среди тех, чье потребление калия было на самом низком уровне по сравнению с двумя верхними тертилями, как 2,6 у мужчин и 4,6 у женщин. В многофакторном анализе увеличение суточного потребления калия на 10 ммоль (одна порция свежих фруктов и овощей) было связано с 40% снижением риска связанной с инсультом смертности, которое не зависело от других диетических переменных и известных факторов риска сердечно-сосудистых заболеваний.
Оральный контрацептив — Имеются сообщения о четырехкратном увеличении случаев геморрагического инсульта у людей старше 35 лет, особенно у курящих.119 Относительный риск ишемического инсульта составляет около трех, а абсолютный риск оценивается в 1 на 10 000 пользователей оральных контрацептивов. По оценкам Совместного исследования ВОЗ по сердечно-сосудистым заболеваниям и контрацепции стероидными гормонами, от 13% до 18% всех инсультов у женщин в возрасте 20-44 лет в Европе и развивающихся странах, соответственно, связаны с использованием оральных контрацептивов120.
Заместительная гормональная терапия —Meade и Berra121 рассмотрели 10 когортных исследований и оценили снижение смертности от инсульта у женщин, получающих заместительную гормональную терапию, на 15%, хотя между центрами наблюдались значительные различия.Исследование медсестры в США не показало значительного снижения ишемического инсульта при комбинированной заместительной терапии эстрогеном и прогестероном, с предположением о повышенном риске инсульта в подгруппе женщин, принимающих самые высокие дозы неконъюгированного эстрогена122.
Витамин C —Acheson и Williams123 в экологическом анализе показали возможное защитное действие витамина C в свежих фруктах и овощах при возникновении инсульта.
ИССЛЕДОВАНИЯ ПО ВМЕШАТЕЛЬСТВУ НА ОСНОВЕ НАСЕЛЕНИЯ
Стратегии снижения заболеваемости или рецидивов инсульта могут быть направлены либо на людей с высоким риском для определенного фактора, либо на всю популяцию.Для снижения частоты инсульта, вероятно, потребуются оба подхода, но оптимальная стратегия еще предстоит определить. Подход, связанный с высоким риском, может включать скрининг населения или выявление оппортунистических случаев, а также выборочное лечение небольшого числа людей, подвергающихся наибольшему риску. Альтернативная, дополнительная стратегия состоит в том, чтобы попытаться добиться небольшого снижения фактора риска у каждого человека в сообществе с помощью той или иной формы вмешательства.
Было проведено несколько крупных интервенционных испытаний на уровне сообществ, в основном предназначенных для определения эффективности стратегий снижения факторов риска при сердечно-сосудистых заболеваниях, но также регистрировалась цереброваскулярная смертность.Большинство доказательств также основано на мужчинах124-126.
Целевая группа по артериосклерозу, созванная Национальным институтом сердца и легких в США, рекомендовала провести исследование множественных факторов риска (MRFIT) у лиц с высоким риском сердечно-сосудистых заболеваний (в возрасте 35-57 лет) из-за комбинаций повышенных липидов. гипертония и курение сигарет должны быть установлены, чтобы определить, приведет ли программа вмешательства к значительному снижению смертности от сердечно-сосудистых заболеваний, нефатального инфаркта миокарда и цереброваскулярной смертности.
После 10,5 лет наблюдения было обнаружено 13% снижение частоты смертельного инсульта в специальной группе вмешательства.4 При рассмотрении вопроса о последующем наблюдении темнокожие субъекты и курильщики с большей вероятностью прекращали свое участие между скрининговыми обследованиями.126 Эффект, который оказал более здоровый образ жизни на контрольных субъектах неизвестно, но возможный эффект мог быть обнаружен в группе вмешательства.
В Северной Карелии, Финляндия127, весь пакет сообщества, представляющий «реальную жизнь», проверял выполнимость таких программ вмешательства с использованием существующих услуг и других ресурсов.В течение 10 лет 1972-82 гг. У субъектов в возрасте 30-59 лет наблюдалось небольшое, но значительное снижение курения, холестерина и артериального давления. Несмотря на несколько исследований, показывающих значительное снижение факторов риска, это не привело к аналогичному снижению показателей смертности от ишемической болезни сердца или инсульта в исследуемых популяциях. Исследование факторов риска Всемирной организации здравоохранения по множественному вмешательству для факторов риска ишемической болезни сердца не показало значительного снижения смертности от ишемической болезни сердца после шести лет вмешательства.128
Однако Вартиайнен и др. 129 оценили степень, в которой изменения основных факторов риска сердечно-сосудистых заболеваний (артериальное давление, курение и концентрация холестерина в сыворотке) могут объяснить изменения в смертности от инсульта в Северной Карелии за последние 20 лет. Они обнаружили, что две трети снижения смертности от инсульта у мужчин и половину у женщин можно объяснить изменениями трех вышеуказанных факторов риска сердечно-сосудистых заболеваний.
Общие медицинские осмотры, проводимые медсестрами в Англии, не помогают курильщикам бросить курить, но они помогают пациентам изменить свой рацион и изменить концентрацию общего холестерина.130 Значение этого изменения в питании для общественного здравоохранения зависит от того, сохранится ли оно. Британское семейное исследование сердца представляет собой рандомизированное контролируемое исследование в общей практике в 13 городах Великобритании для измерения воздействия программы скрининга сердечно-сосудистых заболеваний и изменения образа жизни, проводимой медсестрами.131 Поскольку в большинстве общих практик в Соединенном Королевстве не используются такие интенсивные методы лечения. В программе изменения факторов риска коронарных заболеваний, достигаемые с помощью пакета добровольных мер по укреплению здоровья для первичной медико-санитарной помощи, вероятно, будут еще меньше.Похоже, что такой подход к населению только через первичную медико-санитарную помощь не приведет к значительному снижению риска сердечно-сосудистых заболеваний. Вместо этого необходимо более эффективное законодательство для контроля за употреблением табака и поощрения потребления здоровой пищи.
ОТНОСИТСЯ К ФАКТОРАМ РИСКА ИНСУЛЬТА ИЛИ УБЕЖДЕНИЯ
Знание роли и распространенности факторов риска инсульта необходимо до того, как можно будет предпринять какие-либо попытки по их снижению.Однако при разработке стратегии укрепления здоровья для первичной и вторичной профилактики инсульта важно учитывать собственное отношение пациентов; их восприятие личного риска, воспринимаемого как управляемый или наследственный и, следовательно, фиксированный, и их желание изменить свое контролируемое рискованное поведение.
В этой области было сделано очень мало работы в отношении инсульта, восприятия пациентами или социального имиджа, а также значения инсульта в обществе. Исследование восприятия пациентами риска сердечно-сосудистых заболеваний, проведенное Silagy et al 132 показали, что риски для здоровья, связанные с курением и отсутствием физических упражнений, признаются большинством людей.Отношение к диете было более сложным. Избыточный вес был основным показателем восприятия человеком диеты как вредной для здоровья. Интересно отметить, что в отношении роли диеты имелись серьезные гендерные различия: женщины с ожирением с большей вероятностью думали, что их диета вредна, чем мужчины с ожирением. Мотивация к снижению факторов риска была высокой для курения, но низкой для физических упражнений.
Существуют также важные различия во взглядах и образе жизни между этническими группами, отражающие культурные различия в образе жизни и социальных привычках.Курение сигарет и потребление алкоголя, по-видимому, выше у белых, чем у чернокожих мужчин133, и могут составлять неотъемлемую часть социальной активности этого населения, которое, следовательно, может сопротивляться изменениям. На контроль веса влияет отношение к размеру тела, которого придерживаются различные культурные группы, и практические проблемы, связанные с изменениями в рационе питания в семейном контексте. Различия в отношении к западной медицинской практике между различными культурными группами являются важным фактором, определяющим соблюдение рекомендованных лекарственных препаратов для первичной и вторичной профилактики инсульта.
При разработке программы профилактики инсульта необходимо учитывать не только потенциальную пользу от снижения инсульта путем изменения индивидуальных факторов риска, но и собственное восприятие пациентами личных и социальных издержек изменения поведения.
Несосудистые события
ТРАВМА ГОЛОВКИ
В 1990 году в Соединенном Королевстве было зарегистрировано не менее 4191 случая смерти от перелома черепа и внутричерепных травм.137 Tiret et al 138, по оценке во Франции, было три пика приема — в возрасте до 5 лет, в возрасте от 25 до 24 лет и старше 75 лет.Шестьдесят процентов были связаны с дорожно-транспортными происшествиями и 30% — с падениями.
Политика национальных и местных органов власти играет важную роль в предотвращении несчастных случаев, так же как и укрепление здоровья. Медицинские услуги имеют отношение к сокращению летальности, а не к предотвращению самого заболевания, и не будут обсуждаться далее.
Предотвращение несчастных случаев среди молодых людей может стать возможным путем обучения их навыкам управления дорожным движением и дорожными системами. Важно научить маленьких детей переходить дорогу и дать уроки езды на велосипеде, а также улучшить их видимость.Томпсон и др. 139 обнаружили, что велосипедные защитные шлемы очень эффективны в предотвращении травм головы. Другие эффективные меры включают защитные шлемы для мотоциклистов и шляпы для верховой езды наездников.
Основным фактором, способствующим смерти мужчин в возрасте от 15 до 24 лет, является алкоголь.140 Кампании против алкоголя могут быть эффективными. Законодательство Оклахомы, увеличившее минимальный возраст для покупки алкоголя, сделав предел содержания алкоголя в крови во время вождения до 22 ммоль / л и способствовавшее отзыву лицензии у подозреваемых «пьяных водителей», могло быть ответственным за сокращение числа смертельных случаев и аварий со смертельным исходом. на треть.141 Другие законодательные акты, такие как более широкое использование ограничений скорости и обеспечение соблюдения правил использования передних и задних ремней безопасности, могут помочь снизить количество дорожно-транспортных происшествий или свести к минимуму травмы, которые в результате них.
Повреждения головы при падениях характерны для детей и пожилых людей. Их можно предотвратить, выявляя возможные риски несчастных случаев в доме и устраняя их.143 Особенно у пожилых людей острое заболевание может проявляться падениями. Ранняя диагностика и лечение в сочетании с квалифицированной оценкой жизненной среды для устранения опасностей могут предотвратить дальнейшие падения.
Рабочая группа по профилактике Королевского колледжа врачей144 указала, что превентивные меры лучше всего осуществлять с помощью правоохранительных органов, обучения и использования шлемов и ремней безопасности. Что касается алкоголя, они предложили повысить цены, сократить рекламу, на бутылках должно быть четко указано содержание алкоголя в напитке, следует продвигать слабоалкогольные напитки и должна быть улучшена координация между всеми вовлеченными агентствами.
Медицинский факультет общественного здравоохранения установил целевые показатели по сокращению числа несчастных случаев с 1790 в 1979 году до 55 в 2000 году.145 Стратегия «Здоровье нации» поставила цели в отношении несчастных случаев, чтобы снизить уровень смертности среди детей в возрасте до 15 лет не менее чем на 33%, среди детей в возрасте от 15 до 24 лет на 25% и среди детей старше 65 лет как минимум на 33% к 2005 году146.
ДЕМЕНТИАС
Недавняя конференция в Эдинбурге Проблема деменции показала, что исследования в области генетики и визуализации начинают проливать свет на факторы восприимчивости и открывать возможности для более ранней диагностики147.
С возвращением на повестку дня сосудистых причин деменции и с идеями о роли окислительного стресса мы, возможно, видим первые намеки на профилактические возможности, которые идут параллельно с улучшением физического и психического здоровья.147 Йошитаке и др. 148 провели одно из крупнейших популяционных исследований заболеваемости деменцией и факторами риска в Японии с очень высокими посмертными показателями. У мужчин самой частой причиной деменции является сосудистое происхождение, вопреки сообщениям из других частей мира о том, что болезнь Альцгеймера является преобладающей патологией.
БОЛЕЗНЬ АЛЬЦГЕЙМЕРА
Единственными факторами риска болезни Альцгеймера, о которых однозначно сообщалось, являются возраст, пол и семейный анамнез, ни на один из которых пока что нельзя повлиять.Есть некоторые доказательства, подтверждающие отрицательную связь между курением и болезнью Альцгеймера149.
Роттердамское исследование150 показало обратную зависимость доза-реакция между образованием и деменцией, в частности болезнью Альцгеймера. Это не было связано с сердечно-сосудистыми заболеваниями. Это подтверждает канадское исследование здоровья и старения, 151 которое показало, что люди с низким уровнем образования подвержены более высокому риску болезни Альцгеймера с отношением шансов 4 (95% ДИ 2,49–6,43). Таким образом, возможность первичной профилактики может быть направлена на повышение уровня образования населения.
ДЕМЕНЦИЯ ВТОРИЧНАЯ ПРИ ДРУГИХ МЕДИЦИНСКИХ УСЛОВИЯХ
Связь деменции с гипотиреозом, дефицитом витамина B12, сифилисом и ВИЧ хорошо известна. Первичная профилактика этих состояний может снизить бремя деменции в обществе. Пренатальный скрининг на синдром Дауна и хорею Хантингтона дает возможность предотвратить некоторые случаи деменции.
СОСУДИСТАЯ ДЕМЕНЦИЯ
По сравнению с инсультом было проведено относительно мало исследований потенциальных факторов риска мультиинфарктной деменции.Кажется вероятным, что между этими двумя условиями существует тесная связь. В настоящее время проводятся два исследования факторов риска, но с использованием разных методологий и факторов риска можно назвать только предположительными.152 153
Возраст, пол, раса, образование и риск инфаркта головного мозга были названы факторами риска. Ladurner и др. 154 обнаружили, что гипертония в анамнезе чаще встречается среди пациентов с когнитивными нарушениями и инфарктом головного мозга, чем среди пациентов с инфарктом головного мозга без когнитивных нарушений.Горелик и др. 155 показали, что недавнее курение сигарет и инфаркт миокарда в анамнезе, оба из которых связаны с атеросклерозом, независимо друг от друга позволяют прогнозировать когнитивные нарушения, связанные с множественными церебральными инфарктами, среди преимущественно афроамериканских пациентов с множественным инфарктом. Кроме того, более высокий уровень систолического артериального давления оказывал защитный эффект. Татемичи и др. 156 с другой стороны, обнаружили, что гипертония и сердечно-сосудистые заболевания не были связаны с деменцией после инсульта, а сахарный диабет был.Необходимо уточнить роль других факторов риска.
головная боль или мигрень
В Вашингтоне (США) 157 8% мужчин и 14% женщин пропускали всю или часть работы или учебы из-за головной боли в течение четырех недель перед собеседованием.
По оценкам, ежегодная стоимость потери производительности из-за мигрени в Соединенных Штатах составляет 1,2-17,2 миллиарда долларов. Прямые затраты на медицинские услуги оцениваются в более одного миллиона посещений врача в год.158
Имеющиеся данные подтверждают мнение о том, что семейные и генетические факторы играют роль в определении риска мигрени. Точный генетический образец передачи еще предстоит определить159.
При имеющихся знаниях первичная профилактика не играет никакой роли.
МНОЖЕСТВЕННЫЙ СКЛЕРОЗ
В таблице 6 указаны предполагаемые факторы риска рассеянного склероза, большинство из которых не поддаются первичной профилактике. Доказательства того, что травма является основной причиной заболевания, не являются убедительными.161
Таблица 6Факторы риска рассеянного склероза
Было рассмотрено несколько вирусных инфекций, включая корь и HTLVI, как причины рассеянного склероза. Однако данные о том, что вакцинация против кори защищает от рассеянного склероза, противоречивы162. 163
Никакая первичная профилактическая стратегия невозможна при нынешнем понимании этиологии рассеянного склероза.
БОЛЕЗНЬ ПАРКИНСОНА
Было проведено всего около 10 исследований заболеваемости, и есть проблемы с определением случая.Распространенность паркинсонизма значительно увеличивается с возрастом (14,9% в возрасте 65-74 лет, 29,5% в возрасте 75-84 лет и 52,4% в возрасте 85+ лет) 164.
Мартин и Осмонд, 165 лет, в исследовании случай-контроль не обнаружили никаких доказательств связи между массой тела при рождении и массой тела в возрасте 1 года или домашними условиями для последующего риска. Пациенты с большей вероятностью, чем контрольные, никогда не курили (отношение шансов 2 (95% ДИ 1,1–3,6)). В течение 26 лет наблюдения за более чем 8000 мужчин, участвовавших в кардиологической программе Гонолулу, у курящих мужчин был сниженный риск развития болезни (относительный риск 0.39) .166
Было высказано предположение167, что жизнь в сельской местности, потребление воды из колодцев и воздействие пестицидов увеличивают риск. Проживание в сельской местности также может объяснить связь с двумя вездесущими сельскими патогенами — коронавирусом и Nocardia asteroides , которые, как недавно было предложено, вызывают болезнь Паркинсона168. 169
ЗАБОЛЕВАНИЕ ДВИГАТЕЛЬНЫХ НЕЙРОНОВ
Различные исследования случай-контроль выявили различные факторы риска, включая предшествующие механические, клинические и электрические травмы, воздействие тяжелых металлов и профессии, связанные с кожей, физический труд и физическую активность.170 В одном исследовании случай-контроль не было обнаружено связанного предшествующего заболевания.171
Исследование случай-контроль в Шотландии172 выявило связь между избыточной частотой переломов, физическим трудом и профессиональным воздействием свинца и растворителей, а также повышенным риском заболевания двигательных нейронов. Не было обнаружено связи с социально-экономическими лишениями в детстве, детскими инфекциями или социальным классом. Если эти данные подтвердятся, есть потенциал для первичной профилактики в этой области.
ЭПИЛЕПСИЯ
В эпидемиологических исследованиях использовались самые разные определения эпилепсии.В общих популяционных исследованиях в развитых странах, определяющих эпилепсию как повторяющиеся неспровоцированные приступы, показатели заболеваемости с поправкой на возраст варьируются от 24 до 53 на 100 000 человеко-лет. Исследования первых незарегистрированных изъятий дают несколько более высокие оценки — от 26 до 70 на 100 000 человеко-лет173.
В исследовании, проведенном в Рочестере, США, эпилепсия, связанная с цереброваскулярным заболеванием (8% случаев), была наиболее важной единичной выявленной этиологией среди широко распространенных случаев. Затем последовала эпилепсия, связанная с неврологическим дефицитом с рождения (5%).Травмы головы и инфекция ЦНС являются наиболее важными группами в среднем зрелом возрасте, а также цереброваскулярные заболевания и дегенеративные заболевания головного мозга у пожилых людей.173 Они также связаны с опухолями головного мозга, алкоголем и рассеянным склерозом (см. Другие разделы).
Раннее воздействие пищевых контаминантов изменяет созревание оси человеческого мозга-кишечника-микробиоты
ОСНОВАНИЕ И СОЗРЕВАНИЕ ОСИ МОЗГ-ЖИВОТ-МИКРОБИОТА
Урегулирование кишечного гомеостаза определяется как установление равновесия между микробиотой кишечника, иммунной системой и целостность эпителиальных клеток [3].Это событие происходит в течение 1000 первых дней жизни, от стадии плода до 2 лет [4]. Поскольку микробиота кишечника играет центральную роль в созревании слизистой оболочки кишечника и иммунной системы, многие исследования сосредоточили свой интерес на том, где и когда начнется передача материнской микробиоты. В начале 20 — годов была принята идея о том, что приобретение кишечной микробиоты у младенца начинается с момента родов после заражения вагинальными и фекальными бактериями материнской флоры. Однако новые методы, основанные на молекулярной технике независимого культивирования, предполагают, что перенос бактерий от матери к ребенку начинается с в утробе матери с [5], хотя точные механизмы этого события до конца не изучены.В 2005 и 2008 годах Хименес и др. [6,7] с помощью полимеразной цепной реакции обнаружили присутствие бактерий в околоплодных водах, пуповине и меконии или первом стуле новорожденных. Совсем недавно были проведены исследования по идентификации микробиома плаценты человека после условных и преждевременных беременностей. Firmicutes, Tenericutes, Proteobacteria, Bacteroidetes, Fusobacteria phyla и непатогенная комменсальная микробиота были обнаружены и были связаны с антенатальной инфекцией [8,9].Тем не менее, даже если структура бактерий могла передаваться от матери к ребенку в утробе матери, наблюдались различия в составе микробиоты новорожденного в зависимости от способа родов, преждевременных родов и использования антибиотиков [10-13]. В настоящее время признано, что Staphylococci, Streptococci, Enterococci, Lactobacillus и Bifidobacteria являются первыми бактериями, которые колонизируют наш желудочно-кишечный тракт из вагинальной и фекальной флоры, а некоторые из них происходят в период лактации [14].До третьего года индивидуальная изменчивость выше, чем у взрослых, но диверсификация рациона питания приводит к эволюции микробиоты кишечника за счет увеличения их богатства / разнообразия и будет иметь тенденцию к достижению стабильности во времени, также называемой состоянием эубиоза [15]. Микробиота кишечника способствует улучшению здоровья, например метаболизму пищи, конкуренции с патогенами, росту кишечных клеток и стимуляции иммунной системы [16,17]. В течение этого периода микробиота ребенка остается очень чувствительной к различным воздействиям, которые могут повлиять на ее состав и оказать долгосрочное воздействие на нашу физиологию [18].Тем не менее, изменение микробиоты кишечника, вызывающее дисбактериоз, обычно связано со многими заболеваниями, такими как воспалительные заболевания кишечника, аутизм, диабет 2 типа, ожирение, аутоиммунные заболевания и аллергия [19,20]. Например, клинические испытания предоставляют доказательства взаимосвязи между развитием аллергии и дисбактериозом микробиоты [21] в зависимости от способа родоразрешения у младенцев [22]. Поведение детей в первые 3 года жизни явно способствует значительному воздействию микробов: кормление грудью и прямой контакт с кожей матери, постоянное попадание предметов, рук или ног в рот, особенно во время ползания и на ранних этапах ходьбы, когда руки соприкасаются с напольные поверхности.Однако фактическая чрезмерная гигиена нашей окружающей среды снижает воздействие микробов в молодом возрасте, влияющих на созревание иммунной системы. Это упоминалось в росте иммунных расстройств (аутоиммунные заболевания, аллергии, и т.д. ) на протяжении более 40 лет как «гипотеза гигиены» [23,24].
Напротив, поскольку кишечная микробиота недоношенных новорожденных при рождении менее разнообразна, чем у доношенных, она представляет собой незрелый кишечник с недоразвитой перистальтикой, барьерной функцией и иммунитетом.Таким образом, кишечник является потенциальным источником воспаления и инфекций, вызываемых патогенными бактериями, такими как Enterobacter , Enterococcus и Staphylococcus [25, 26].
Гомеостаз слизистой оболочки также является результатом перинатального установления индуцированной слизистой оболочкой иммунной толерантности, феномена, регулируемого набором сигналов, обеспечиваемых клетками врожденного иммунитета, которые формируют адаптивные иммунные ответы. Благодаря своей барьерной функции, опосредованным контактом клеткам сигналам и продукции цитокинов, эта регуляторная иммунная сеть контролируется эпителием слизистой оболочки.Перинатальное созревание иммунной системы слизистой оболочки также может регулироваться прямым или непрямым воздействием факторов окружающей среды, независимо от того, какой путь воздействия рассматривается. Во время беременности иммунная система матери и плода взаимодействует двунаправленным образом. Развитие и созревание иммунной системы начинается в раннем возрасте плода и продолжается в младенчестве и раннем детстве. Это окна высокой восприимчивости и уязвимости к экологическим оскорблениям, таким как недоедание, стресс или загрязнение окружающей среды.Развитие иммунной системы плода характеризуется наличием нескольких отличительных особенностей, которые способствуют активной толерантности плода как к материнским антигенам in utero , так и к экзогенным антигенам, таким как инфекционные агенты. Хорошо известно, что эндогенные и экзогенные факторы определяют развитие и созревание иммунной системы кишечника. У грызунов и человека образование вторичных лимфоидных структур, таких как бляшки Пейера или брыжеечные лимфатические узлы (MLN), происходит еще до рождения.Однако их размер и развитие зародышевых центров зависят от послеродовой микробной колонизации [27,28]. Миелоидные клетки, такие как макрофаги, дендритные клетки (ДК) или микроглиальные клетки головного мозга, начинают заселять ткани уже на 4-7 неделе беременности и продолжают увеличиваться в количестве на протяжении второго триместра [29]. Затем с 8 по 18 неделю тимус будет заселен лимфоидными клетками в процессе созревания лимфоцитов и положительного и отрицательного отбора (центральная толерантность).Популяция в тимусе будет продолжать увеличиваться по мере протекания беременности [29]. Первые зрелые Т-клетки обнаруживаются в периферических тканях между 10 и 12 неделями беременности, и значительное их количество циркулирует в конце второго триместра [30]. У мышей врожденные клетки, такие как индуцированные лимфоидной тканью клетки, естественные клетки-киллеры или Т-хелперы 2, мигрируют из печени плода в слизистую оболочку кишечника благодаря эндогенным сигналам в течение первых 4 недель после рождения [31]. Бактериальная колонизация будет стимулировать рекрутирование вилочкового белка 3 (FOXP3) и регуляторных Т-клеток (Treg) на слизистую оболочку кишечника и секрецию противовоспалительного цитокина интерлейкина-10 (IL-10) [31,32].С помощью этого механизма Treg-клетки держат провоспалительные Th-клетки под контролем, чтобы сохранить эпителиальный барьер. Было обнаружено большое количество Treg-клеток в человеческих плодных MLNs [33], и их возвращение, по-видимому, особенно важно в младенчестве [34]. Bifidobacterium infantis , в основном присутствующие в просвете кишечника, сильно индуцирует FOXP3 + Treg-клетки у детей грудного возраста [35]. И наоборот, с развитием врожденного иммунного ответа большинство микроорганизмов, колонизирующих тонкую и толстую кишку, будут анаэробами.Слизистая оболочка тонкой кишки новорожденных людей имеет архитектуру зрелого крипта-ворсинка с непрерывной пролиферацией стволовых клеток, а также миграцией и дифференцировкой эпителиальных клеток. Изменения в составе антимикробных пептидов и антибактериальной активности в ранний постнатальный период также наблюдались в просвете кишечника новорожденных человека [36]. Кишечный иммунный гомеостаз также устанавливается благодаря тесному сотрудничеству между эпителиальными клетками кишечника (IEC) и компонентами лимфоидной ткани, связанной с субэпителиальной кишкой.В слизистой оболочке кишечника молекулы, часто связанные с патогенами, могут распознаваться поверхностными рецепторами распознавания образов и, среди них, Toll-подобными рецепторами (TLR). После активации передачи сигналов TLR, IECs будут высвобождать трансформирующий фактор роста бета (TGFβ), с помощью которого вызываются толерогенные фенотипы DCs [37]. Затем DC секретируют IL-10 в начале иммунного ответа, в котором доминируют Treg-клетки. Параллельно может индуцироваться Th2-иммунитет и секретироваться противовоспалительные цитокины [38,39].Генерация FOXP3 + Treg и сопутствующее установление иммунной толерантности являются результатом двух этапов. С одной стороны, пул-основатель FOXP3 + Tregs продуцируется активацией антиген-специфичных наивных Т-клеток в кишечных дренирующих MLN. С другой стороны, перемещение активированного Treg в собственную пластинку кишечника благодаря макрофагам кишечника. Самонаводящиеся клетки FOXP3 + Treg размножаются, и необратимо устанавливается иммунная толерантность. Чтобы обеспечить пассивный иммунитет плоду или новорожденному, материнские антитела передаются плоду через плаценту (начиная с 6 недель беременности с быстрым увеличением к 24-26 неделям), а затем младенцу через грудное вскармливание.Периферическая активация иммунной системы с выработкой цитокинов может влиять на центральную нервную систему (ЦНС) напрямую через цитокинов, пересекающих гематоэнцефалический барьер, или косвенно, активируя гипоталамо-гипофизарную ось надпочечников [40].
Наконец, гомеостаз эндокринной среды матери и плода также важен для нормального развития и предотвращения побочных эффектов, возникающих в результате ранних инсультов. Изменение гестационной эндокринной среды из-за болезней затем нарушит гормональный гомеостаз и может быть причиной программирования измененного эндокринного профиля у плода.Важный путь, по которому кишечные микробы и их метаболиты связываются с ЦНС, включает клетки, составляющие эндокринную систему кишечника [41]. Энтероэндокринные клетки расположены между эпителиальными клетками кишечника по всей длине кишечника и способны выделять различные типы молекул, которые могут проникать в системный кровоток и влиять на регуляторные пути ЦНС. 5-гидрокситриптамин или серотонин (5-HT), главный гормон, регулирующий моторику и секрецию желудочно-кишечного тракта, вырабатывается энтерохромаффинными клетками и накапливается в этих клетках и кишечных нейронах, а также в ЦНС.Большинство молекул, участвующих в нейроэндокринной передаче сигналов по оси мозг-кишечник-микробиота, происходят из метаболизма пищевой микробиоты кишечника, как и в случае триптофана, предшественника нейромедиатора 5-HT [42].
В то время как нейронные связи начинают устанавливаться во время эмбрионального периода, их полное развитие не достигается до достижения взрослого возраста. Это причина того, почему нейрональная организация мозга / периферии остается очень чувствительной в течение длительного периода жизни. Благодаря эпидемиологическим наблюдениям, развивающийся мозг, как известно, очень чувствителен как к внешним, так и к внутренним сигналам окружающей среды еще в пренатальном периоде.В этот период метаболическая и иммунная деятельность матери будет зависеть от внешней среды. Их реактивность будет мешать развитию нервной системы в утробе матери, и любое изменение материнской эндокринной или иммунной реакции может иметь серьезные необратимые последствия для развития плода из-за эпигенетических модификаций. Эти исследования связывают нарушения развития нервной системы, такие как шизофрения или аутизм, с микробной инфекцией во время жизни плода [43–45]. Хотя считается, что плацента защищает плод, помещая его в почти стерильную среду, материнская среда во многих отношениях влияет на развитие мозга плода.Действительно, метаболиты материнской микробиоты могут преодолевать плацентарный барьер и влиять на программирование построения мозга плода, которое зависит от плацентарных нейрогормонов, таких как серотонин, необходимый для развития переднего мозга [46], или метаболических гормонов, таких как адипокины или стероиды, контролирующих пролиферацию, дифференциацию, развитие нервной системы. [47,48]. Более того, микробиом кишечника матери регулирует выработку серотонина энтерохромафинными клетками [49]. Его истощение приводит к резкому развитию мозга [50].Дисбиоз матери также влияет на формирование гематоэнцефалического барьера [51].
Развитие мозга будет продолжаться постнатально, и метаболическое программирование расхода энергии будет зависеть от правильного урегулирования связей между ядром дуги и гипоталамуса, которые имеют место в первые пару дней после рождения. Синаптогенез начинается вскоре после рождения и достигает максимального уровня примерно к 2 годам до того, как синаптическое очищение происходит в середине подросткового возраста для получения взрослой нейрональной организации [52].
Этот период развития нейронов параллелен заселению и созреванию кишечной микробиоты — процессу, который, как известно, необходим для установления надлежащей иммунной функции [56-60], нейроэндокринной системы [61] и регуляции метаболизма [15,62]. Кишечные микробы связываются с ЦНС по крайней мере через три параллельных и взаимодействующих канала, включая эндокринные, нервные и иммунные сигнальные механизмы. Таким образом, урегулирование этой двунаправленной коммуникации очень важно для сохранения здоровья, согласно большому количеству литературы.Изменение этого перекрестного разговора будет лежать в основе патогенеза и патофизиологии классических расстройств мозга / кишечника, таких как желудочно-кишечные расстройства, такие как синдром раздраженного кишечника или воспалительные заболевания кишечника, но также и растущий список психических расстройств или аутизма [63,64], Болезнь Паркинсона [65] или хроническая боль [66]. Можно подозревать, что неправильное урегулирование этой двунаправленной коммуникации лежит в основе всех этих хронических неинфекционных заболеваний, а именно изменения заселения кишечной микробиоты в раннем возрасте.
Во время чувствительного окна развития химическое воздействие окружающей среды на химические вещества, особенно через пищевые пути, будет действовать при формировании тканей, влияя на фенотип, сдвигая взаимодействия иммунного метаболизма и микробиоты с последствиями для нейро-иммуно-эндокринной системы. на уровне кишечника и мозга, и это затем повлияет на функции органов и предрасположенность к развитию хронических заболеваний в более позднем возрасте [67]. Кроме того, многие механизмы, представленные здесь, находятся под генетическим контролем, включая специфическую экспрессию генов на каждой стадии развития и созревания.Эти механизмы допускаются конкретными факторами транскрипции, которые взаимодействуют с ДНК для активации или ингибирования экспрессии генов. Эпигенетические механизмы связаны с доступностью ДНК для этих факторов транскрипции и не передаются классической менделевской генетикой. Эпигенетика связана с биохимическими модификациями, которые влияют на уплотнение и доступность ДНК, в основном с метилированием ДНК по специфическому азотному основанию цитозина и посттрансляционным модификациям гистоновых белков.miRNA — еще один эпигенетический механизм, при котором эти малые РНК от 21 до 24 нуклеотидов способны гибридизоваться в основном в области 3’-UTR мРНК и блокировать трансляцию или вызывать деградацию мРНК [68]. Изменения эпигенетических модификаций представлены как ключевые события программирования развития [69,70]. Эпигенетические механизмы по своей природе пластичны и могут накапливаться с течением времени, что во время программирования развития индивидуумов окажет глубокое влияние на экспрессию генов и предрасполагает к развитию профиля фенотипов заболевания [71].
«Триалог» среди бактериальных обитателей, кишечные барьеры и нейро-иммуноэндокринные компоненты, ассоциированные с кишечником, обычно должны обеспечивать возникновение и сохранение стабильных симбиотических отношений, укрепляющих гомеостаз кишечника. Однако, если организм вступает в контакт с токсичными веществами окружающей среды, которые переносятся в просвет через прием пищи, эти состояния могут быть причиной нестабильности, увеличивая риск развития хронических заболеваний, особенно в критических периодах жизни, таких как перинатальная стадия.
РОЛЬ ПИЩЕВЫХ КОНТАМИНАНТОВ В ПРОГРАММИРОВАНИИ ПЛОДА
На основании всей этой литературной информации можно задаться вопросом, может ли пренатальное или послеродовое воздействие пищевых загрязнителей каким-либо образом перепрограммировать общее развитие плода или младенца и, в частности, может помешать заселению микробиоты, восстанавливая созревание слизистой оболочки кишечника не только на иммунном, но и на нейроэндокринном уровне. Можно предположить, что все эти модификации естественным образом повлияют на развивающуюся связь между кишечником и мозгом.Это ясно доказано, что воздействие определенных химических веществ в послеродовой период и в раннем детстве предрасполагает к значительным и необратимым повреждениям развивающегося мозга. Беременность — это особенное состояние высокой уязвимости для женщины и плода из-за необходимой адаптации к предположительно вредной среде, которая может изменить симбиотическое взаимодействие между матерью и ребенком и может, в свою очередь, повлиять на процесс программирования плода. Это может привести к некоторым постоянным, часто незначительным изменениям в различных органах, среди которых кишечник или мозг.Многие факторы во время и после беременности могут повлиять на состояние здоровья человека в будущем: генетический статус, воспаление во время беременности, а также диета. Действительно, пища не только обеспечивает питательные вещества, но также является наиболее важным источником загрязнителей окружающей среды различного происхождения, включая тяжелые металлы [72] или стойкие органические загрязнители (СОЗ), поступающие в результате промышленной деятельности и содержащиеся в продуктах питания, особенно в жирной рыбе [73]. Некоторые другие, такие как пестициды, могут быть обнаружены в собранных культурах [74] или могут появиться во время хранения, как в случае с грибковыми или бактериальными токсинами [75], или во время обработки пищевых продуктов и, в частности, нагревания с появлением продуктов реакции Майяра (MRP). [76].Плод подвергается воздействию загрязняющих веществ из рациона матери. Поскольку все эти вещества непреднамеренно обнаруживаются в пищевых продуктах, при определенных обстоятельствах они могут нанести вред здоровью. Человеческий плод — это не маленький взрослый человек. Эта уязвимая субпопуляция, вероятно, подвергается большему риску заражения пищей из-за более высокой скорости абсорбции, плохой способности к детоксикации, быстрой пролиферации клеток и незрелых механизмов восстановления. В этой части мы рассмотрим последствия воздействия пищевых загрязнителей на мать во время беременности на созревание плода, а затем на младенца и ребенка.
Воздействие антибиотиков на плод
Как упоминалось выше, материнская микробиота передается плоду во время беременности через механизмы, частично понятные, и во время естественных родов микрофлора попадает в кишечник новорожденного, определяя первоначальную микробную популяцию при условии, что роды осуществлялись без кесарево сечение, без дезинфицирующих средств, антибиотиков во время родов, антисептических кремов, и т. д. [77]. Таким образом, нарушения микрофлоры влагалища матери (диета, состав тела, инфекции, лечение антибиотиками, стресс, пробиотики, и т. Д. .) определенно будет иметь серьезные последствия для потомства. Во время беременности вагинальные бактериальные инфекции могут повлиять на раннее развитие нервной системы и, по-видимому, связаны с расстройством аутистического спектра [78], а после родов могут изменить микробный состав кишечника новорожденного. Это уменьшит разнообразие и стабильность популяций микробиоты, которые будут влиять на ферментацию, переваривание и абсорбцию метаболитов, увеличение числа устойчивых к антибиотикам бактерий, таких как Klebsiella , Citrobacter , Enterobacter , E.coli [79-81]. В совокупности эти эффекты изменят развитие новорожденного и младенца и увеличат риск его заболевания.
У грызунов воздействие антибиотиков на самок во время лактации приводит к истощению численности Lactobacilli , увеличению жировой массы и изменению секреции метаболических гормонов у их потомства [82,83].
Воздействие пестицидов на плод
Воздействие пестицидов на потомство после воздействия на мать широко задокументировано в литературе.Тем не менее, несколько исследований подтвердили их вредное воздействие при многократном воздействии на низком уровне. Многие из них уже давно описаны как эндокринные разрушители. Органофосфаты и карбаматы, такие как хлорпирифос (CPF), малатион, диазинон, карбарил, обладают общей ингибирующей активностью в отношении холинэстеразы. Более того, эти органофосфаты сильно влияют на баланс микробиоты [84]. Было доказано, что воздействие диазинона на самок приводит к поведенческим нарушениям у их потомства [85]. Признано, что химические вещества окружающей среды, особенно факторы, нарушающие работу эндокринной системы, будут вызывать функциональную модификацию экспрессии генов, и когда эти изменения происходят в течение жизни плода, это может быть причиной повышенного риска дисфункции и заболевания в более позднем возрасте, хотя любое изменение фенотипа будет наблюдаться. при рождении [67].
Карбофуран, карбамат антихолинэстеразы, широко используется в качестве инсектицида, нематицида и акарицида в сельскохозяйственной практике во всем мире. Карбофуран и / или его основные метаболиты могут проникать через плацентарный барьер и оказывать серьезное воздействие на материнско-плацентарно-плодный блок. Токсичность карбофурана может быть усилена одновременным воздействием с другими ингибиторами холинэстеразы [86].
Хлорпирифос [O, O-диэтил-O- (3,5,6-трихлор-2-пиридинил) фосфоротиоат: CPF] — это фосфорорганический инсектицид, используемый во всем мире для обработки фруктов и овощей.Наиболее широко изученный органофосфат, CPF, был четко идентифицирован как убийца нейронов, нарушитель миграции нервных клеток и соединений мозга, а его остатки часто обнаруживаются в пище и питьевой воде [87]. Хотя пищеварительный тракт является первым органом, вступающим в контакт с загрязнителями пищи, мало что известно о влиянии CPF на эпителиальный барьер. Исследования Joly Condette и др. [88] показали, что хроническое воздействие CPF во время критических пре- и постнатальных периодов развития и созревания органов изменяет функцию эпителиального барьера, что, в свою очередь, связано с повышенной проницаемостью и бактериальной транслокацией.Кроме того, дисфункция барьера связана с изменениями экспрессии белков плотных контактов. В той же модели крыс, подвергшихся воздействию CPF в дозе 5 мг / кг / сут, были значительно меньше (длина тела) и легче, чем в контрольной группе. Воздействие CPF было связано с изменениями гистологических структур (более короткие и тонкие кишечные ворсинки), микробным дисбиозом кишечника и повышенной бактериальной транслокацией в селезенке и печени. Эти значительные микробные изменения в кишечнике были связаны с нарушенной защитой эпителия (муцин-2) и экспрессией генов рецепторов распознавания микробных паттернов (TLR 2 и 4) [84].Таким образом, остатки пестицидов в пище могут влиять на функцию пищеварительного тракта и его способность адаптироваться к изменениям окружающей среды. У крыс этот эффект проявляется еще сильнее во время отлучения от груди (, т.е. , когда происходит диверсификация корма). Все больше данных указывает на то, что CPF участвует в метаболических нарушениях. Данные Reygner и др. [89] указывают на то, что воздействие CPF на развитие нарушает метаболизм с дозозависимыми эффектами, очевидными во взрослом возрасте. Сегодня, когда интеллектуальные нарушения развития затрагивают каждого шестого ребенка в промышленно развитых странах, растет интерес к выявлению воздействия пестицидов на развитие и созревание мозга, а также к этиологии интеллектуальных нарушений.Исследование, проведенное той же французской группой, оценило, может ли употребление матерью низкой дозы CPF у крыс нарушить церебральную функцию их потомства [90]. Согласно результатам, потомство самок, обработанных CPF, показало более медленный отрицательный геотаксис у новорожденных, меньшую новизну исследования у молодых и более быстрый рефлекс испуга у подростков и взрослых. Эти данные свидетельствуют о том, что развитие CPF, связанное с воздействием на человека, может нарушать активность, связанную с новизной, и сенсомоторные функции, тем самым адаптируясь к окружающей среде.Эти данные подтверждают гипотезу о том, что CPF может способствовать поведенческим расстройствам, включая задержку приобретения и последствия во взрослом возрасте. Еще одно фосфорорганическое соединение было изучено на предмет его потенциального воздействия на здоровье. Ацефат — это пестицид, поражающий насекомых, который также относится к хлорорганическим соединениям. Что касается CPF, модель крыс Wistar на животных предполагает, что воздействие ацефата во время беременности и кормления грудью вызывает изменения в метаболизме глюкозы у матери и программирует у потомства предрасположенность к диабету 2 типа во взрослом возрасте [91].
Хлорорганические пестициды (ХХП) — это загрязнители окружающей среды, которые сохраняются в окружающей среде и биоаккумулируются через пищевую цепочку у людей и животных. Результаты Yamazaki и др. [92] подтверждают гипотезу о том, что пренатальное воздействие ХОП, особенно эпоксида цис-гептахлора, может оказывать неблагоприятное воздействие на развитие нервной системы младенцев в определенном возрасте, даже при низких уровнях. Хлордекон — это стойкий ХОП, который использовался во Французской Вест-Индии до начала 1990-х годов для борьбы с банановым долгоносиком.В проспективном продольном исследовании, проведенном в Гваделупе (когортное исследование матери и ребенка Тимун), воздействие хлордекона измерялось при рождении по образцу пуповинной крови и по образцу грудного молока, взятому через 3 месяца после родов. Полученные результаты свидетельствуют о том, что пренатальное воздействие хлордекона связано со специфическими нарушениями мелкой моторики у мальчиков, и дополняют растущее число свидетельств того, что воздействие ХОП в раннем возрасте ухудшает развитие ребенка [93]. Аналогичным образом, в той же когорте Тимун в Гваделупе другой целью исследования было изучить связь между пренатальным воздействием хлордекона и ростом плода, измеренным по массе при рождении [94].Они обнаружили значительную U-образную связь между массой тела при рождении и воздействием хлордекона в пределах верхних квартилей увеличения веса во время беременности или чрезмерного увеличения веса во время беременности. Воздействие хлордекона может повлиять на рост плода, особенно при чрезмерном увеличении веса во время беременности. Другие пестициды, такие как дихлордифенилтрихлорэтан и его высокотоксичный метаболит дихлордифенилдихлорэтилен (ДДЭ), являются стойкими ХОП, которые представляют собой липофильные загрязнители окружающей среды, накапливающиеся в пищевой цепи.Эти химические вещества недавно были изучены на предмет их возможных опасностей для здоровья, таких как рак и репродуктивные последствия, включая низкий вес при рождении. В исследовании Ханджани и др. [95] изучали, отличаются ли матери с более высоким уровнем загрязнения пестицидами от матерей с низким уровнем заражения в отношении исходов рождения их потомства, таких как масса тела при рождении, малая для возраста гестации, недоношенность, окружность головы, пол. соотношение, и предыдущий выкидыш или мертворождение.В этом случае не было обнаружено, что эти пестициды связаны с неблагоприятными исходами родов у зараженных матерей в диапазоне контаминации нашего населения (<7,5 мг / кг липидов в материнском молоке), хотя есть слабые доказательства того, что соотношение полов может быть затронуто. . Воздействие на мать рыбы, пойманной в ходе спортивных соревнований в районе Великих озер, загрязненной ДДЕ, предполагает, что воздействие ДДЕ на плод (на что указывает концентрация ДДЭ в сыворотке крови матери) может снизить вес при рождении [96].
Амитраз — активное средство защиты растений, обладающее противопаразитарным действием и принадлежащее к химическому семейству формамидинов.Kim и др. [97] исследовали потенциальные побочные эффекты амитраза на начало и поддержание беременности у крыс Sprague-Dawley, а также его влияние на развитие эмбриона и плода после воздействия на мать в течение всего периода беременности. Амитраз вводили беременным крысам через желудочный зонд с 1 по 19 дни беременности в дозах 0, 3, 10 и 30 мг / кг / день. Всем самкам было выполнено кесарево сечение на 20-й день беременности, а их плоды были обследованы на наличие каких-либо внешних, висцеральных и скелетных аномалий.При дозе 30 мг / кг материнская токсичность проявлялась в увеличении частоты патологических клинических признаков и снижении увеличения массы тела и потребления пищи. Токсичность, связанная с развитием, включала увеличение смертности плода, уменьшение размера помета и уменьшение массы тела плода. Кроме того, увеличилась частота внешних, висцеральных и скелетных аномалий плода. При дозе ниже 10 мг / кг наблюдаемая материнская токсичность включала снижение прибавки массы тела и уменьшение потребления пищи, связанное с минимальной токсичностью для развития.Это включает уменьшение массы тела плода, увеличение висцеральных и скелетных аберраций и задержку окостенения плода. Эти результаты показывают, что амитраз, вводимый крысам в течение всего периода беременности, является эмбриотоксичным и тератогенным в дозе токсичности для матери (, т.е. , 30 мг / кг / день) и минимально эмбриотоксичен при минимальной дозе для материнской токсичности (, т.е. . , 10 мг / кг / сут).
Диурон широко используется в качестве средства уничтожения сорняков для уничтожения нежелательных трав и других однолетних и вечнозеленых широколистных сорняков, особенно в виноградарстве.Он также используется в садах и для прополки обочин дорог или железных дорог. Перинатальное воздействие диурона [3- (3,4-дихлорфенил) -1-1-диметилмочевины] может оказывать неблагоприятное воздействие на лимфоидные органы крысы. Беременным крысам Sprague-Dawley вводили диурон в дозе 500, 750 или 1250 частей на миллион в рационе с 12 по 21 день гестации (GD 12-21) и во время лактации. Проточный цитометрический анализ выявил значительное снижение B-лимфоцитов (CD45RA +) у самцов, но на Т-лимфоциты (CD4 +, CD8 + и CD4 + / CD8 +) заметного воздействия не наблюдалось.Таким образом, данные свидетельствуют о том, что вызванная диуроном материнская токсичность у самок, подвергшихся воздействию высоких доз и перинатальное воздействие этого гербицида, вызвала токсичность для селезенки, о чем свидетельствует снижение количества B-лимфоцитов у самцов детенышей Sprague-Dawley [98].
Воздействие микотоксинов на плод
В связи с растущими доказательствами пагубного воздействия пестицидов на здоровье все больше и больше людей предпочитают употреблять органические продукты. Однако в западных странах растет внимание к растущему потреблению органических продуктов, поскольку фактические условия хранения урожая не могут предотвратить заражение пищевых продуктов микотоксинами, особенно когда это касается органических продуктов.Это беспокойство еще более важно, когда речь идет о беременных женщинах и младенцах. Действительно, зерновые, предназначенные для потребления человеком, могут быть заражены видами грибов из родов Aspergillus , Penicillium, Fusarium и Alternaria , которые способны продуцировать очень вредные вторичные метаболиты: микотоксины [75]. Наиболее актуальными во всем мире микотоксинами в зерновых являются афлатоксины (AF), охратоксин A (OTA), патулин (PAT), фумонизины, зеараленон (ZE) и трихотецены, включая токсин Т-2 и дезоксиниваленол (DON) [99].На основании имеющихся в настоящее время данных можно предположить, что перенос микотоксинов через плаценту происходит уже на стадии гестации и накапливается в кровообращении плода к концу беременности. Микотоксины, образующиеся в результате потребления зараженной материнской пищей, могут проникать через плацентарный барьер, метаболизироваться развивающимся плодом и могут влиять на системы плода [100]. Эта уязвимая популяция, вероятно, подвергается большему риску от канцерогенных токсинов окружающей среды из-за высокой скорости абсорбции, плохой элиминационной способности, быстрой пролиферации клеток и незрелых механизмов восстановления [101].О влиянии микотоксинов на плод во время вынашивания проводится мало исследований. Более того, профессиональное воздействие микотоксинов в зерне вызывает роды на ранних сроках беременности [102].
Только одно исследование на людях изучали воздействие ФП и недоношенность, но исследования на животных показывают, что воздействие ФП может повышать риск недоношенности и потери беременности. Плод может пострадать от материнской AF через прямую токсичность, а также косвенную токсичность, через материнское системное воспаление, нарушение роста плаценты или повышение уровня плацентарных цитокинов.Цитотоксические и системные эффекты ФП могут быть причиной материнской анемии, задержки внутриутробного развития, потери плода и преждевременных родов [103]. Исследование, проведенное на конкретной человеческой популяции, представленной на экспозицию AF, показало нарушение эпигенетических модификаций. В частности, была показана корреляция между частотой ФП у матери во время беременности и характером дифференциального метилирования многих генов у ребенка после рождения, чувствительных к влиянию на экспрессию генов [104]. Действительно, другое исследование клеток пищеварительного тракта показало, что воздействие другого микотоксина фумонизина B1 изменяет метилирование промоторов генов, показывая, что этот механизм, возможно, является общим [105].
Wangikar et al [106], 2014 изучали действие микотоксинов ОТА и афлатоксина B1 (AFB 1 ), вводимых перорально беременным крысам, и выявили, что эти микотоксины вызывают висцеральные аномалии, такие как гастрошизис. Arora и др. [107] сообщили, что однократная доза AFB 1 , введенная беременной самке через желудочный зонд, вызвала выпячивание кишечника после воздействия на 8-й день беременности. Лечение беременных крыс AFB 1 привело к образование доброкачественных и злокачественных опухолей в печени, желудке, кишечнике, эндокринных органах, а также в центральной и периферической нервной системе у потомства.
ZE, эстрогенный микотоксин, продуцируемый Fusarium graminearum или F. roseum , является одним из наиболее распространенных контаминантов зерновых культур во всем мире. Целью исследования Коллинза и его коллег было определение эффектов ZE на внутриутробное развитие крыс [108]. Беременным самкам крыс Charles River Sprague-Dawley один раз в день вводили через желудочный зонд ZE (в кукурузном масле) в дозах 0, 1, 2, 4 или 8 мг / кг массы тела на GD 6-19. Таким образом, ZE был токсичен для матери и плода, но не был тератогенным.
Воздействие других органических загрязнителей на плод
Бисфенол А (BPA) — это органическое соединение из семейства ароматических, которое используется в основном в производстве пластмасс и смол. Он отмечен как фактор риска развития пищевой аллергии и пищевой непереносимости — двух нежелательных пищевых реакций, которые учащаются во всем мире. Menard и др. [109] оценили последствия перинатального воздействия низких доз BPA на иммуноспецифический ответ на пищевой антиген овальбумин (OVA) в зрелом возрасте.Когда OVA-толеризованным крысам, обработанным BPA, вводили перорально OVA, возникало воспаление толстой кишки с инфильтрацией нейтрофилов, увеличением IFNγ и снижением TGFβ. Они показывают, что перинатальное воздействие BPA изменило пероральную толерантность и иммунизацию к диетическим антигенам (OVA). Таким образом, наивная иммунная система новорожденного уязвима для низких доз BPA, которые вызывают пищевую непереносимость в более позднем возрасте.
СОЗ — это химические вещества с высокой устойчивостью, включая диоксины, фураны, полихлорированные бифенилы и ХОП, созданные в результате промышленной деятельности и обнаруженные в продуктах питания, причем самые высокие уровни содержания обнаруживаются в жирной рыбе [73].Было обнаружено, что эти высокотоксичные соединения проникают через плаценту и выделяются с грудным молоком [110]. Воздействие СОЗ в раннем возрасте связано с иммунотоксичностью развития и нарушениями развития нервной системы, такими как расстройства аутистического спектра (см. Обзор Dietert et al [111]). Это сложное взаимодействие между иммунной системой, загрязнителями окружающей среды и развитием нервной системы было описано разными авторами, хотя точные механизмы до сих пор не совсем понятны [112].Однако пренатальное и послеродовое воздействие СОЗ явно связано с повреждением головного мозга [113,114]. Наконец, было описано, что большинство СОЗ и особенно тетрахлордибензофлуран, полихлорированные бисфенилы (ПХБ) или хлороталонил, еще один ХОП, сильно влияют на состав кишечной микробиоты [115], но это еще предстоит подтвердить у младенцев.
Воздействие тяжелых металлов на плод
Для населения в целом наиболее распространенными источниками воздействия токсичных тяжелых металлов являются воздух и потребление пищи.Из-за своей высокой уязвимости беременные женщины и младенцы даже больше, чем кто-либо другие, обеспокоены их воздействием, поскольку их воздействие на здоровье является серьезным. Ртуть — это глобальные загрязнители окружающей среды, возникающие в результате естественных процессов и антропогенной деятельности. В большинстве случаев воздействие ртути на человека происходит через потребление рыбы, моллюсков и морских млекопитающих, загрязненных метилртутью (MeHg) [116]. Нейротоксическая опасность, которую представляет MeHg для людей, и уникальная восприимчивость развивающегося мозга были хорошо задокументированы после массовых отравлений, произошедших в Японии и Ираке.Почему у плода проявляются разные нейропатологические эффекты и более высокая чувствительность к MeHg по сравнению со взрослым, пока неизвестно. В зависимости от степени воздействия in utero , MeHg может приводить к эффектам, варьирующимся от гибели плода до незначительной задержки нервного развития. На основании эпидемиологических исследований, проведенных в группах населения с умеренным хроническим воздействием MeHg, на сегодняшний день не было достигнуто окончательного консенсуса относительно уровня безопасности воздействия на мать во время беременности [117]. В исследовании, проведенном Cambier и др. [118], беременные крысы были заражены экологически значимыми дозами 36 и 76 нг MeHg / г корма с использованием рационов, содержащих рыбу, содержащую ртуть.Новорожденные были защищены от воздействия ртути через плацентарный барьер, поскольку у новорожденных от матерей, получавших рацион, содержащий 76 нг MeHg / г пищи, концентрации Hg в мозге, почках, печени и скелетных мышцах составляли 12, 3, 21 и 18% ткани их матери, соответственно. Эти результаты предполагают существование, по крайней мере, у крыс, порогового уровня с точки зрения воздействия MeHg, выше которого разрушается плацентарный барьер. В поведенческих тестах, проведенных в возрасте 5 и 6 недель, крысы, подвергшиеся воздействию MeHg, показали значительный дефицит моторной координации в тесте с вращающимся стержнем и неспособность к обучению в тесте реакции пассивного избегания по сравнению с контрольной группой.Гистопатологически наблюдалась очаговая дисплазия мозжечка, включая гетеротопическое расположение клеток Пуркинье и гранулярных клеток. Эти нарушения могут быть вызваны действием большого количества MeHg, накопленного в головном мозге в период беременности [119]. Как показано выше, плоды особенно чувствительны к воздействию MeHg, и неблагоприятное воздействие на развитие ребенка было связано с уровнями воздействия, которое приводит к небольшому количеству признаков материнского клинического заболевания или токсичности, если таковые имеются. Высокие уровни пренатального воздействия на людей приводят к нейроповеденческим эффектам, таким как церебральный паралич и тяжелая умственная отсталость.Пренатальное воздействие MeHg в сообществах с хроническим воздействием низкого уровня связано со снижением массы тела при рождении и ранней сенсомоторной дисфункцией, такой как задержка начала ходьбы [120]. Более того, воздействие ртути на поздних сроках беременности усилило токсическое воздействие свинца на развитие нервной системы младенцев в возрасте 6 месяцев [121]. Кадмий, как известно, оказывает разрушающее действие на эндокринную систему [122]. In utero Воздействие кадмия на человека было связано с модификацией метилирования генов, связанных с метаболизмом липидов и минерализацией костей [123].В результате можно запрограммировать повышенное системное накопление жира и повышенную частоту переломов костей и остеопороза в более позднем возрасте [123]. Мышьяк является сильнодействующим токсичным веществом и канцерогеном, и его можно найти в самых разнообразных продуктах питания, включая рыбу и рис [124]. Клиническое исследование, основанное на образцах пуповины, показало, что воздействие мышьяка на беременных женщин было ответственно за повышенную экспрессию 12 miRNA, которая, как предполагается, участвует в регуляции сигнальных путей, обычно нарушенных при раке и сахарном диабете [125].Таким образом, это исследование подтвердило нарушение регуляции экспрессии miRNA после воздействия мышьяка на плод и частично объясняет, как мышьяк способствует развитию этих заболеваний.
У детей с Фарерских островов, рожденных от практически здоровых матерей, которые ели загрязненную ртутью рыбу во время беременности, были обнаружены измененные нейронные связи, о чем свидетельствует снижение показателей внимательности и памяти [126]. Воздействие свинца может вызвать повышенный риск гипертонии при беременности и выкидыша [127]. Мышьяк, свинец и MeHg связаны с сокращением срока беременности или преждевременными родами [127–130].Французская группа сообщила о результатах перекрестных исследований женщин до беременности и во время третьего триместра. Они показали, что воздействие до беременности было выше, чем в последнем триместре. Причина такой разницы, по-видимому, частично связана с увеличением массы тела, которое не было компенсировано параллельным увеличением количества потребляемой пищи [131]. Это было одно из первых исследований во Франции, показывающее уровень воздействия пищевых загрязнителей на плод и влияние на него изменений в потреблении пищи во время беременности и сезонных колебаний.Потребление молочных продуктов связано с более низким уровнем ртути и свинца как у беременных женщин, так и у детей. Всасывание свинца усиливается при дефиците кальция, тогда как свинец может нарушать цикл гомеостаза и функции кальция [132, 133]. Это объясняет, почему высокое потребление молочных продуктов, то есть высокое потребление кальция, может снизить абсорбцию свинца и связанные с ним нейротоксические эффекты [134]. До сих пор очень мало исследований оценивали совместное воздействие тяжелых металлов. Насколько нам известно, только одна недавняя работа проиллюстрировала, что совместное воздействие свинца и ртути на поздних сроках беременности отрицательно сказывается на нервном развитии младенцев в возрасте 6 месяцев [121].
Воздействие MRP на плод
С тех пор, как первый человек обнаружил огонь, наши пищевые привычки сильно изменились, и эта эволюция заметно усилилась в прошлом веке параллельно с глубоким изменением нашего образа жизни, особенно в северных модернизированных странах. Приготовление пищи стало необходимым для улучшения пищевых и органолептических качеств, гигиены, а также для облегчения их хранения в течение более длительного периода. Однако этот процесс трансформации сопровождается появлением новых соединений, среди которых MRP.Этот широкий набор молекул появляется при варке и является результатом реакции между карбонильной группой восстановленного сахара и свободной аминогруппой аминокислоты, обычно ε-аминогруппой остатков лизина в белках. Эта первоначальная реакция затем приводит к образованию большого количества молекул в результате множественных перегруппировок в источнике вкуса приготовленной пищи, от коричневатого до жареного на гриле. Однако конечные продукты продвинутого гликирования (AGE), акриламид или гетероциклический амин, принадлежащие к гликотоксинам, также могут оказывать негативное влияние на здоровье [135].Пищевые AGE всасываются через тонкий кишечник, и их концентрация в крови увеличивается на протяжении всей жизни и еще больше усугубляет их эндогенное производство [136]. Воздействие высоких AGE во время внутриутробного развития может привести к воспалительным состояниям в молодом возрасте, таким как ювенильный диабет [137]. Как у грызунов, так и у человека потребление диеты, богатой AGE (AGE-RD), отрицательно влияет на гомеостаз глюкозы и способствует появлению свободных радикалов. Csongová и др. [138] недавно показали, что потребление AGE-RD во время беременности может ускорить созревание рефлексов у их потомства.Более того, они показали, что эта диета предрасполагает потомство мужского пола к увеличению веса и влияет на метаболизм глюкозы [138]. К сожалению, исследование не выявило какого-либо воздействия на пищеварительный тракт. В литературе описывается, что эндогенно продуцируемые AGE способны взаимодействовать с рецептором конечных продуктов гликозилирования (RAGE), что приводит к пролонгированной активации ядерного фактора каппа B (NF-κB), что приводит к транскрипции провоспалительных цитокинов, включая TNFα [139, 140 ]. Затем ограничение потребления AGE может быть полезным для снижения уровней некоторых воспалительных маркеров, помогая ограничить риски развития в более позднем возрасте прогрессирования хронических состояний [141].Два продольных исследования показали связь между воздействием акриламида на мать и более низкой массой тела при рождении или более высоким риском рождения маленького ребенка для гестационного возраста [142, 143]. Более того, увеличение воздействия акриламида с пищей на мать во время беременности было связано с более высокой распространенностью детей с избыточной массой тела / ожирением в возрасте 3, 5 и 8 лет [144]. Пищевой акриламид, который в основном содержится в жареном картофеле, хлопьях для завтрака, печенье и кофе, всасывается из желудочно-кишечного тракта и, попав в системный кровоток, быстро распространяется в тканях.Акриламид признан нейротоксикантом [145] и может оказывать токсическое воздействие на репродуктивную функцию и развитие [146]. Это очень важно, поскольку было подсчитано, что 10-50% акриламида проходит через плацентарный барьер [147,148]. Гормональный и эндокринный эффект акриламида неоднозначен. Акриламид изменяет активность щитовидной железы, поскольку сообщалось, что метаболиты акриламида в моче отрицательно связаны со свободным тироксином [149] у подростков и молодых людей. Функция щитовидной железы плода также может влиять на рост плода [150], поскольку гормоны щитовидной железы необходимы для оптимального роста.Это может частично объяснить задержку роста, наблюдаемую у детей, подвергшихся воздействию акриламида. Diamanti-Kandarakis и др. [151] показали, что воздействие пищевых AGE влияет на метаболические и гормональные функции самок крыс. Шестимесячное воздействие сильно нагретой пищи грызунов, богатой AGE, увеличило: (1) отложение AGE в тканях; (2) проявление ярости; и (3) повышенные уровни тестостерона в плазме в клетках теки яичников. Кроме того, у самок крыс линии Wistar, получавших диету с высоким возрастом, снизились уровни эстрадиола и прогестерона.Эти женские половые гормоны имеют решающее значение для имплантации оплодотворенной яйцеклетки в матку, поддержания беременности, а также для регулирования полового и менструального циклов [151]. Такие эффекты могут иметь серьезные последствия для развивающегося плода. Было доказано, что мононуклеарные клетки периферической крови крыс, изолированные от самок крыс, получавших высокий AGE, имели повышенную экспрессию RAGE, рецептора AGEs, связанного с провоспалительными путями [152, 153]. Это может быть механизм, также стимулируемый у развивающегося ребенка, который может способствовать изменению функции программирования.
РОЛЬ ПИЩЕВЫХ КОНТАМИНАНТОВ В РАЗВИТИИ И СОЗРЕВАНИИ МЛАДЕНЦЕВ И ДЕТЕЙ
Хотя воздействие загрязнителей пищевых продуктов на плод может сильно повлиять на программирование развития, нельзя исключать постнатальное воздействие этих загрязнителей как важный фактор, способствующий модификации этого программирования. Нарушение развивающегося кишечника и установления перекрестной связи между мозгом и кишечником загрязнителями пищи может привести к снижению способности устанавливать местные и системные нейро-иммуно-эндокринные функции и повышенному риску развития неврологических и воспалительных заболеваний.В самом деле, серьезные или новые эволюционные проблемы могут быть причиной нарушения нормальных сигнальных путей, объясняющих появление этих нарушений рано или поздно. Однако, несмотря на то, что все больше и больше исследований подтверждают последствия воздействия материнского вещества во время беременности на плод и младенца / ребенка, очень мало исследований было посвящено прямому воздействию загрязнителей пищевых продуктов на ребенка и младенца. Это будет важно для выяснения всех этих эффектов, поскольку младенцы и дети очень уязвимы для пищевых загрязнителей, поскольку они потребляют большое количество пищи по сравнению с их весом, имеют более высокий уровень метаболизма и, таким образом, подвергаются большему воздействию на них, чем взрослые .В этой части обзора мы суммируем фактическое состояние дел, учитывая, что большинство из приведенных ниже исследований часто связаны с заселением кишечной микробиоты, но очень мало работ, связанных с нейротоксичностью пищевых контаминантов для молодых особей.
Послеродовое воздействие антибиотиков
Фармацевтические препараты, среди которых слабительные [154] или антибиотики [155], имеют большое влияние на активность микробиоты кишечника и могут иметь последствия для урегулирования местных и системных взаимодействий с нейроиммуноэндокринными функциями .Нарушение общего эубиоза приводит к перемещению обилия резидентных кишечных бактерий и предрасполагает ребенка к определенным воспалительным или функциональным расстройствам [156]. Использование антибиотиков в послеродовой период может серьезно изменить эту хрупкую экосистему расселения, особенно если их вводить очень рано и в течение длительных периодов времени [157]. Этот нарушенный профиль, связанный со сниженной микробной активностью, будет условиями, способствующими расселению кишечных патогенов [158, 159]. Это было связано с долгосрочным нарушением функции кишечного барьера, но также будет влиять на рост доминирующих бактерий в кишечнике человека [157], подтверждая, что воздействие антибиотиков в автомобиле в раннем возрасте делает ребенка восприимчивым к многочисленным заболеваниям в более позднем возрасте [160, 161]. .Нарушение состава и биоразнообразия кишечной микробиоты часто связано с увеличением протеобактерий из-за более высокого содержания генов устойчивости к противомикробным препаратам в этом типе [162], а нарушение нашей микробиоты, вызванное антибиотиками, способствует развитию неинфекционных хронических заболеваний. заболевания (NCCD), характерные для дисбактериоза. Наконец, изменение состава микробиоты, наблюдаемое у младенцев, получавших антибиотики, было близко к таковому у детей от матерей, получавших антибиотики до родов, с последствиями в позднем возрасте [163].Более того, лечение антибиотиками имеет основные недостатки, заключающиеся в обогащении нашей микробиоты генами устойчивости к антибиотикам, которые впоследствии могут стать проблемой для гомеостаза кишечника. Кроме того, нарушение расселения микробиоты также нарушит ее общую метаболизирующую активность, что касается ее способности преобразовывать пищевые загрязнители и ограничивать их влияние на наше здоровье (для обзора см. Flandroy et al [164]). Вдобавок к этому, хотя использование антибиотиков может быть очень важным для предотвращения сепсиса у недоношенных или новорожденных детей, при некоторых обстоятельствах они могут влиять на рецепторы нейротрансмиттеров, особенно гамма-аминомасляной кислоты [165] или серотонина [ 166], ставя под угрозу правильные центральные и периферические нейронные цепи, которые могут при длительном воздействии, иметь серьезные последствия [167].Эти токсикологические эффекты были подтверждены для амоксициллина у молодых крыс [168].
Ротавирусная инфекция — очень частая причина гастроэнтерита, поскольку поражает в основном желудочно-кишечный тракт. У животных совпадают несколько наблюдений: ротавирусная инфекция снижает активность кишечных ферментов как у поросят [169], так и у молодых мышей [170,171]. У кроликов он нарушает гомеостаз эпителия, изменяя мембрану щеточной каймы кишечника [172]. Ротавирусная инфекция у детей приводит к появлению плоской слизистой оболочки с тотальной вирусной атрофией [173].Воздействие микроорганизмов окружающей среды на ранний период жизни сильно влияет на экспрессию белка на щеточной кайме, поскольку явно стимулирует экспрессию генов TLR5 и TLR9 у облученных новорожденных мышей [174]. Это несоответствие при рождении ослабит созревание кишечного эпителия.
Послеродовое воздействие пестицидов
Исследования на животных свидетельствуют о том, что воздействие токсического стресса в раннем возрасте может усиливать воздействие химических веществ окружающей среды на развитие нервной системы [175].Загрязнение воздуха и пестициды могут влиять на рост в младенчестве или детстве, и они связаны с проблемами развития нервной системы и поведением [176].
Фосфаторганические пестициды в высоких дозах нейротоксичны и нацелены на нервную систему детей и связаны с изменениями нейромедиаторов, включая серотонин, норадреналин, ацетилхолин и дофамин. Связь фосфорорганических пестицидов и IQ детей в возрасте 7 лет была отмечена Stein и др. [177] в 2016 году и уже отмечена [178] как более важная для детей, которые также испытывают определенные социальные невзгоды.Те, кто подвергается наибольшему воздействию токсичных веществ из окружающей среды, часто являются теми же группами и людьми, которые испытывают наибольшие невзгоды. Дети с высоким пренатальным воздействием органофосфатов, которые также испытывают специфический ранний и стойкий токсический стресс, могут подвергаться большему риску неблагоприятного когнитивного развития [177]. Однако постнатальных исследований органофосфатов немного, и большинство результатов противоречивы [179].
Продолжительное грудное вскармливание было связано с повышением содержания ОКП в образцах крови норвежских детей [180].В исследованиях, в которых измеряли уровни ДДЭ и ПХБ, хлорорганических соединений в постнатальном периоде, авторы сообщили о более медленном росте и более высоком риске нарушения эндокринной функции у детей [181].
Глифосат — один из агрохимикатов, который также заслуживает особого внимания у младенцев и детей. Фактически, этот гербицид ингибирует путь шикимата, который, если он присутствует в бактериях, контролирует синтез ароматических соединений, таких как тирозин или триптофан, ключевых посредников в системных эффектах нашей микробиоты (см. Первый раздел этого обзора) [164] .С настоящего момента, похоже, нет прямого влияния неонатального воздействия глифосата на созревание кишечной оси мозга. Однако, поскольку он может изменить развитие матки, нельзя исключить какое-либо изменение гомеостаза кишечника [182], поскольку было доказано, что этот гербицид может временно изменять состав комменсального бактериального сообщества [183].
Послеродовое воздействие микотоксинов
Младенцы и маленькие дети более уязвимы для микотоксинов, чем взрослые, из-за их постоянно растущей, плохо развитой нервной, иммунной, репродуктивной и пищеварительной систем, высокого уровня метаболизма и ограниченного питания.Воздействие микотоксинов в детстве, вызванное употреблением зараженных пищевых продуктов: грудного молока [184], детских смесей или продуктов для прикорма с зерновыми [185], овощами или орехами [186 187]. Это было связано с плохим ростом и развитием ребенка [188], повышенной восприимчивостью к инфекциям и многими другими эффектами, такими как иммунные нарушения [189], развитие опухолей [190], преждевременное половое созревание [191, 192], и т. Д. . Оценка рисков для здоровья, связанных с приемом микотоксинов в детстве, была проведена в последние годы.Присутствие микотоксинов в кишечной жидкости ребенка может вызвать более значительное повреждение кишечных энтероцитов из-за меньшего размера кишечного эпителия по сравнению со взрослым. Вспышка среди детского населения Кашмирской долины была связана с употреблением в пищу естественно загрязненного хлеба с различным количеством трихотеценовых микотоксинов [193]. Симптомы желудочно-кишечного заболевания проявились через 15 минут — 1 час после его употребления. ДОН, принадлежащий к этим трихотеценам, может влиять на жизнеспособность и пролиферацию иммунных клеток.Затем это событие подавляет биосинтез белков, которые предотвращают продукцию цитокинов [194]. Прямым следствием этого события является более высокая восприимчивость к инфекционным заболеваниям, что особенно важно для таких уязвимых людей, как младенцы и младенцы. Более того, было доказано, что ДОН влияет на барьерную функцию кишечника у кур [195].
AF может быть сгенерирован Aspergillus Fungi . Один из его метаболитов, 4-гидроксилированная форма, может секретироваться с молоком млекопитающих после приема микотоксина.Что касается ДОН, ФП снижают экспрессию противовоспалительных цитокинов ИЛ-4 вместе с повышением восприимчивости к инфекции [196].
Другая вспышка диареи у детей в Швеции была связана с употреблением фруктов, загрязненных PAT [197]. Действительно, сообщалось, что потребление микотоксинов в высоких концентрациях (ДОН и ПАТ) может вызвать у людей гастроэнтерит с рвотой [198,199]. PAT цитотоксичен для энтероцитов, увеличивает проницаемость кишечных монослоев Caco-2 и изменяет транспорт ионов в интактных слизистых оболочках кишечника [199,200]. Fusarium Микотоксины довольно часто встречаются в детском питании, особенно в развивающихся странах. Несмотря на усилия, предпринятые для корректировки процесса нагревания, чтобы ограничить или устранить микотоксины из кукурузы или сои, было доказано, что это преобразование пищи не только не устраняет микотоксины, но, скорее, способствует образованию многих других микотоксинов, что представляет более высокий риск для потребительские и особенно младенцы [184].
Другие группы метаболитов, продуцируемых некоторыми Aspergillus или Penicillium , являются охратоксинами.Однако, исходя из фактических знаний, кажется, что это не имеет никаких последствий ни для кишечника, ни для зерна, а, скорее, для почек.
Наконец, фумонизины, микотоксины Fusarium, также могут быть найдены в детском и детском питании. Из реальной литературы до сих пор не установлено, что они ответственны за заболевания кишечника, за исключением вспышки болезней пищевого происхождения в Индии [193]. Однако они, по-видимому, изменяют расселение нервной системы, поскольку связаны с дефектами нервной трубки на границе Мексики и Техаса [201].
Послеродовое воздействие других органических загрязнителей
Несколько СОЗ были связаны с увеличением проблем с поведением детей [202]. Некоторые исследования показали, что воздействие в течение 24 часов до 5 загрязнителей [TCDD, дельтаметрин, гексабромциклододекан, бензо (а) пирен и 2-амино-1-метил-6-фенилимидазо (4,5-b) пиридин] и одна смесь Загрязняющие вещества полициклические ароматические углеводороды (ПАУ) значительно изменили структуру летучих микробов, предполагая, что это могло повлиять на активность микробиоты [203].Более того, их прямое или косвенное влияние на нейронную коммуникацию и развитие сети уже описано [204]. Принимая во внимание всю эту информацию, нельзя исключить возможность неправильной формы поселения кишечной микробиоты у новорожденных и ощутимые последствия для его / ее здоровья. СОЗ, ПХБ, ПАУ были описаны как значительно изменяющие метаболизм микробиоты, лежащие в основе индукции провоспалительного статуса кишечника [203]. 2,3,7,8-тетрахлордибензофуран, СОЗ, может изменить соотношение фирмикутов / Bacteroidetes , которое лежит в основе изменения метаболизма желчных кислот [205].Более того, ПАУ могут трансформироваться микробиотой кишечника в эстрогенные метаболиты [206].
В том же направлении, фталаты, категория пищевых пластификаторов, могут сильно нарушить баланс оседания микробиоты кишечника и мозга. Действительно, воздействие диэтилгексилфталата во время лактации показало отрицательную связь с умственным индексом [202]. Концентрация метаболитов фталата была выше у детей 2-5 лет по сравнению с 11-летними. Послеродовое высокое воздействие эфира фталевой кислоты связано с симптомами синдрома дефицита внимания и гиперактивности у детей [207], и, в частности, диэтилфталат вызывает дисбактериоз кишечника и постоянную потерю веса.Один из предложенных механизмов заключался в активации рецептора арилуглеводородов [208]. Более того, воздействие фталатов на детей коррелировало со снижением метилирования ДНК конкретных генов, среди которых TNFα. Поскольку это деметилирование связано со снижением уровня белка TNFα, это может объяснить нарушение регуляции экспрессии генов, наблюдаемое во время воспалительной реакции, как это предполагалось при астме [209]. Эти недавние наблюдения говорят в пользу корреляции, которую можно указать между контаминантами и молекулярными путями, участвующими в данном заболевании.
Продолжительность грудного вскармливания была основным определяющим фактором и была в значительной степени связана с повышением концентрации СОЗ у детей. Продолжительное грудное вскармливание было связано с повышением концентрации СОЗ, таких как ПХБ, и полибромированных дифениловых эфиров в образцах крови детей [180]. В своем исследовании Caspersen и др. [180] выявили, что концентрации СОЗ у 3-летних детей были в 1,4 раза выше, чем в независимой выборке беременных женщин.
Послеродовое воздействие тяжелых металлов
Существует множество исследований, касающихся пренатального и послеродового воздействия тяжелых металлов, таких как кадмий, мышьяк, ртуть или свинец.Авторы сообщают о более медленном послеродовом росте у детей во всех случаях, но из-за ограниченного числа исследований по конкретным тяжелым металлам авторы не смогли сделать вывод о каких-либо других побочных эффектах [210]. Воздействие ртути, кадмия и свинца в молодом возрасте может нарушить развитие нервной системы, и такое воздействие может повлиять на когнитивное, двигательное и поведенческое развитие ребенка [72]. Основным путем воздействия кадмия является его вдыхание, а затем прием внутрь из-за употребления в пищу злаков и овощей, выращенных на загрязненных почвах.
Из 278 девятилетних детей у 21% общая концентрация ртути в ногтях ног (от 1,5 до 6 мкг / г) была выше, чем уровни, рекомендованные Агентством по охране окружающей среды США (1 мкг / г Hg) для оптимального здоровья детей, связанные с этим. с агрессивным поведением [211].
Снижение коэффициента интеллекта к 1,5 и 5 годам было связано с продолжительностью детства и количеством воздействия мышьяка [212]. Из других исследований на мышах было установлено, что микробиота кишечника может метаболизировать интродуцированные токсичные метаболиты мышьяка [213], которые будут мешать передаче сигналов нейронов, ограничивать массу мозга, а также количество нейронов и глии [214].
Послеродовое воздействие MRP
В зависимости от продолжительности и температуры нагрева пищи количество, тип и разнообразие MRP могут значительно измениться. До сих пор не было описано прямых доказательств того, что MRP влияют на развитие оси микробиоты кишечника головного мозга, хотя большая часть AGE всасывается через кишечник [215]. Однако есть несколько свидетельств, подтверждающих активацию иммунного ответа в присутствии MRP, вероятно, из-за фиксации этих молекул на RAGE и его последующей активации пути NF-κB, приводящего к высвобождению провоспалительных цитокинов, таких как как TNFα, IL-6 или IL-1β и увеличение окислительного стресса тканей наряду с воспалительной реакцией [216].Мы и другие в настоящее время работаем над установлением какой-либо корреляции с перинатальными событиями.
У младенцев, подвергшихся воздействию акриламида, наблюдается задержка роста [217]. Частично это объясняется пренатальным изменением программирования щитовидной железы плода. Однако следует исследовать другое окно восприимчивости, и в дополнение к пренатальной экспозиции следует учитывать послеродовую экспозицию ребенка. Другой возможный механизм, связывающий воздействие акриламида и рост, — это окислительный стресс и воспаление.Действительно, воздействие акриламида может привести к усилению окислительного стресса за счет увеличения экспрессии CYP2E1, что в дальнейшем приводит к усилению перинатального воспалительного статуса [218, 219]. Пищевые AGE ответственны за окислительный стресс и воспаление в некоторых тканях, вмешиваясь в жизненно важные гормоны, вызывая физиологические реакции. Воздействие высокого ВОЗРАСТА в детстве является причиной воспалительного статуса. Это предположение подтверждается взаимосвязью между высокими материнскими AGE и уровнями инсулина в плазме младенцев [137].Новорожденные и младенцы могут быть очень обеспокоены AGE, особенно те, которым необходимо получать гидролизованные смеси для младенцев, которые имеют очень высокие уровни N ε -карбоксиметиллизина (CML) по сравнению с грудным молоком из-за тепловой обработки молочных белков в наличие лактозы [220]. В недавнем обзоре мы предположили, что вновь образованные соединения из реакции Майяра и особенно ХМЛ могут быть новым фактором риска аллергии [221], поскольку они могут реагировать с рецепторами RAGE, экспрессируемыми на поверхности многочисленных иммунных клеток (мононуклеарных клеток, DC и T клетки), присутствующие в областях, где AGE часто более многочисленны [222-224].
Как лечится заболевание ЦНС при остром лимфобластном лейкозе (ОЛЛ) у детей?
Pui CH, Robison LL, Look AT. Острый лимфобластный лейкоз. Ланцет . 2008 22 марта. 371 (9617): 1030-43. [Медлайн].
Кампана Д. Роль мониторинга минимальной остаточной болезни при остром лимфобластном лейкозе у взрослых и детей. Гематол Онкол Клин Норт Ам . 2009 г., 23 (5): 1083-98, vii. [Медлайн]. [Полный текст].
Рибера Дж. М., Ориол А.Острый лимфобластный лейкоз у подростков и молодых людей. Гематол Онкол Клин Норт Ам . 2009 23 октября (5): 1033-42, vi. [Медлайн].
Марголин Дж. Ф., Стеубер С. П., Поплак Д. Г.. Острый лимфобластный лейкоз. Pizzo PA Poplack DG, ред. Принципы и практика детской онкологии . 15-е изд. 2006. 538-90.
Hunger SP, Lu X, Devidas M, Camitta BM, Gaynon PS, Winick NJ, et al. Повышение выживаемости детей и подростков с острым лимфобластным лейкозом в период с 1990 по 2005 год: отчет детской онкологической группы. Дж Клин Онкол . 2012 10 мая. 30 (14): 1663-9. [Медлайн]. [Полный текст].
Pui CH, Mullighan CG, Evans WE, Relling MV. Острый лимфобластный лейкоз у детей: куда мы идем и как мы туда доберемся ?. Кровь . 2012 августа 9. 120 (6): 1165-74. [Медлайн]. [Полный текст].
Ribeiro KB, Buffler PA, Metayer C. Социально-экономический статус и заболеваемость острым лимфолейкозом в детском возрасте в Сан-Паулу, Бразилия. Инт Дж. Рак .2008 15 октября. 123 (8): 1907-12. [Медлайн].
Питерс Р., Шраппе М., Де Лоренцо П., Ханн И., Де Росси Г., Феличе М. и др. Протокол лечения детей младше 1 года с острым лимфобластным лейкозом (Interfant-99): обсервационное исследование и многоцентровое рандомизированное исследование. Ланцет . 21 июля 2007 г. 370 (9583): 240-50. [Медлайн].
Маграт I, Шанта В., Адвани С., Адде М., Арья Л.С., Банавали С. и др. Лечение острого лимфобластного лейкоза в странах с ограниченными ресурсами; уроки использования единого протокола в Индии за двадцатилетний период [исправлено]. Eur J Cancer . 2005 июл. 41 (11): 1570-83. [Медлайн].
Муваккит С., Аль-Ариди С., Самра А., Сааб Р., Махфуз Р.А., Фарра С. и др. Внедрение интенсивного протокола стратифицированного по риску лечения детей и подростков с острым лимфобластным лейкозом в Ливане. Am J Hematol . 2012 Июль 87 (7): 678-83. [Медлайн].
de Labarthe A, Rousselot P, Huguet-Rigal F, Delabesse E, Witz F, Maury S, et al. Иматиниб в сочетании с индукционной или консолидирующей химиотерапией у пациентов с острым лимфобластным лейкозом de novo, положительным по филадельфийской хромосоме: результаты исследования GRAAPH-2003. Кровь . 2007 15 февраля. 109 (4): 1408-13. [Медлайн].
Dubansky AS, Boyett JM, Falletta J, Mahoney DH, Land VJ, Pullen J, et al. Изолированная тромбоцитопения у детей с острым лимфобластным лейкозом: редкое событие в исследовании группы детской онкологии. Педиатрия . 1989 декабрь 84 (6): 1068-71. [Медлайн].
Gaynon PS. Острый лимфобластный лейкоз в детстве и рецидив. Br J Haematol . 2005 декабрь 131 (5): 579-87.[Медлайн].
Ганесан П., Тулкар С., Гупта Р., Бахши С. Детский алейкемический лейкоз с гиперкальциемией и поражениями костей, имитирующими метаболическое заболевание костей. J Педиатр Метаб эндокринола . 2009 Май. 22 (5): 463-7. [Медлайн].
Kawedia JD, Kaste SC, Pei D, Panetta JC, Cai X, Cheng C и др. Фармакокинетические, фармакодинамические и фармакогенетические детерминанты остеонекроза у детей с острым лимфобластным лейкозом. Кровь .2011, 24 февраля, 117 (8): 2340-7; викторина 2556. [Medline]. [Полный текст].
te Loo DM, Kamps WA, van der Does-van den Berg A, van Wering ER, de Graaf SS. Прогностическое значение бластов в спинномозговой жидкости без плейоцитоза или травматической люмбальной пункции у детей с острым лимфобластным лейкозом: опыт Голландской группы детской онкологии. Дж Клин Онкол . 2006 20 мая. 24 (15): 2332-6. [Медлайн].
FDA одобрило применение препарата Гливек для детей с острым лимфобластным лейкозом [пресс-релиз].Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США. Доступно на http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm336868.htm. Доступ: 1 февраля 2013 г.
Lee S, Kim YJ, Min CK, Kim HJ, Eom KS, Kim DW и др. Влияние промежуточной терапии иматинибом первой линии на исход аллогенной трансплантации стволовых клеток у взрослых с впервые диагностированным острым лимфобластным лейкозом, положительным по филадельфийской хромосоме. Кровь . 2005 1 мая. 105 (9): 3449-57. [Медлайн].
Pulsipher MA, Peters C, Pui CH.Острый лимфобластный лейкоз у детей высокого риска: трансплантировать или не трансплантировать ?. Пересадка костного мозга Biol . 2011 17 января (1 приложение): S137-48. [Медлайн]. [Полный текст].
Schultz KR, Bowman WP, Aledo A, Slayton WB, Sather H, Devidas M и др. Улучшение ранней бессобытийной выживаемости с иматинибом при остром лимфобластном лейкозе с положительной хромосомой в Филадельфии: исследование детской онкологической группы. Дж Клин Онкол . 2009, 1. 27 (31): 5175-81. [Медлайн].[Полный текст].
Schrappe M, Hunger SP, Pui CH, Saha V, Gaynon PS, Baruchel A, et al. Исходы после неудачной индукции при остром лимфобластном лейкозе у детей. N Engl J Med . 2012 Апрель 12, 366 (15): 1371-81. [Медлайн]. [Полный текст].
Buitenkamp TD, Израэли S, Zimmermann M, Forestier E, Heerema NA, van den Heuvel-Eibrink MM, et al. Острый лимфобластный лейкоз у детей с синдромом Дауна: ретроспективный анализ группы исследования Понте ди Леньо. Кровь . 2014 2 января 123 (1): 70-7. [Медлайн]. [Полный текст].
Bhojwani D, Pui CH. Рецидив острого лимфобластного лейкоза в детском возрасте. Ланцет Онкол . 2013 май. 14 (6): e205-17. [Медлайн].
Маллиган К.Г., Филлипс Л.А., Су X, Ма Дж., Миллер С.Б., Шуртлефф С.А. и др. Геномный анализ клонального происхождения рецидивирующего острого лимфобластного лейкоза. Наука . 28 ноября 2008 г. 322 (5906): 1377-80. [Медлайн]. [Полный текст].
Блинцито (блинатумомаб) [вкладыш в упаковке]. Таузенд-Окс, Калифорния: Amgen Inc., декабрь 2014 г. Доступно на [Полный текст].
Schlegel P, Lang P, Zugmaier G, Ebinger M, Kreyenberg H, Witte KE, et al. Педиатрический посттрансплантационный рецидивирующий / рефрактерный острый лимфобластный лейкоз, предшественник B-предшественника, демонстрирует стойкую ремиссию при терапии блинатумомабом, биспецифическим антителом, взаимодействующим с Т-клетками. Haematologica . 2014 Июль 99 (7): 1212-9. [Медлайн].[Полный текст].
Паркер С., Уотерс Р., Лейтон С., Хэнкок Дж., Саттон Р., Мурман А.В. и др. Влияние митоксантрона на исходы у детей с первым рецидивом острого лимфобластного лейкоза (ALL R3): открытое рандомизированное исследование. Ланцет . 2010 декабрь 11, 376 (9757): 2009-17. [Медлайн]. [Полный текст].
Campana D, Leung W. Клиническое значение минимальной остаточной болезни у пациентов с острым лейкозом, перенесших трансплантацию гемопоэтических стволовых клеток. Br J Haematol . 2013 Июль 162 (2): 147-61. [Медлайн].
Одобрение FDA дает первую генную терапию в Соединенных Штатах. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США . 30 августа 2017 г. Доступно по адресу https://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm574058.htm.
Eapen M, Raetz E, Zhang MJ и др. совместное исследование Детской онкологической группы и Центра международных исследований по трансплантации крови и костного мозга. Кровь . 15 июня. 107 (12): 4961-7.
Cheok MH, Evans WE. Острый лимфобластный лейкоз: модель фармакогеномики противораковой терапии. Нат Рев Рак . 2006 Февраль 6 (2): 117-29. [Медлайн].
Джонс Л.К., Саха В. Филадельфийский положительный острый лимфобластный лейкоз у детей. Br J Haematol . 2005 августа 130 (4): 489-500. [Медлайн].
Brentjens RJ, Curran KJ. Новые клеточные методы лечения лейкемии: Т-клетки, модифицированные CAR, нацелены на антиген CD19. Hematology Am Soc Hematol Educational Program . 2012. 2012: 143–51. [Медлайн].
CAR Т-клетки: разработка иммунных клеток пациентов для лечения их рака. Национальный институт рака. Доступно по адресу https://www.cancer.gov/about-cancer/treatment/research/car-t-cells#option. 7 августа 2017 г .; Доступ: 30 августа 2017 г.
Смит М., Гудман Б. FDA одобрило первое в своем роде лечение рака. Медицинские новости Medscape. Medscape Medical News .30 августа 2017 г. Доступно по адресу: http://www.webmd.com/cancer/news/20170830/fda-approves-breakthrough-cancer-treatment?ecd=wnl_nal_083017&ctr=wnl-nal-083017_nsl-ld-stry_1&mb=jt7Y3G4gtU2VE3GcIFTYV2 % 40qU% 3d.
Kymriah (tisagenlecleucel) [вкладыш в упаковке]. Восточный Ганновер, Нью-Джерси: Novartis Pharmaceuticals Corporation. Август 2017 г. Доступно в [Полный текст].
Mullighan CG, Su X, Zhang J, Radtke I., Phillips LA, Miller CB, et al.Удаление IKZF1 и прогноз при остром лимфобластном лейкозе. N Engl J Med . 2009 29 января. 360 (5): 470-80. [Медлайн]. [Полный текст].
Mullighan CG, Zhang J, Harvey RC, Collins-Underwood JR, Schulman BA, Phillips LA, et al. Мутации JAK при остром лимфобластном лейкозе у детей высокого риска. Proc Natl Acad Sci U S A . 2009 г. 9 июня. 106 (23): 9414-8. [Медлайн]. [Полный текст].
Fuster JL, Bermúdez M, Galera A, Llinares ME, Calle D, Ortuño FJ.Мезилат иматиниба в сочетании с химиотерапией у четырех детей с de novo и поздней стадией филадельфийского хромосомно-положительного острого лимфобластного лейкоза. Haematologica . 2007 декабрь 92 (12): 1723-4. [Медлайн].
Hackethal V. TKI могут стать новым вариантом лечения для ВСЕХ подтипов. Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/831525. Доступ: 24 сентября 2014 г.
Робертс К.Г., Ли Ю., Пейн-Тернер Д., Харви Р.С., Ян Ю.Л., Пей Д. и др.Целевые поражения, активирующие киназу, при Ph-подобном остром лимфобластном лейкозе. N Engl J Med . 2014 11 сентября. 371 (11): 1005-15. [Медлайн]. [Полный текст].
Боггс В. Неврологические изменения происходят спустя долгое время после детской лимфоидной злокачественной опухоли. Медицинские новости Medscape. Доступно на http://www.medscape.com/viewarticle/810241. Дата обращения: 11 сентября 2013 г.
Gordijn MS, Gemke RJ, van Dalen EC, Rotteveel J, Kaspers GJ. Подавление оси гипоталамус-гипофиз-надпочечники (HPA) после лечения глюкокортикоидной терапией острого лимфобластного лейкоза у детей. Кокрановская база данных Syst Rev . 2012 16 мая. 5: CD008727. [Медлайн].
Henze G, v Stackelberg A, Eckert C. ALL-REZ BFM — последовательные испытания для детей с рецидивирующим острым лимфобластным лейкозом. Клин Падиатр . 2013 май. 225 Приложение 1: S73-8. [Медлайн].
Hijiya N, Barry E, Arceci RJ. Клофарабин при остром лейкозе у детей: текущие результаты и проблемы. Рак крови у детей . 2012 Сентябрь 59 (3): 417-22.[Медлайн].
Хилл Ф.Г., Ричардс С., Гибсон Б., Ханн И., Лиллейман Дж., Кинси С. и др. Успешное лечение без краниальной лучевой терапии детей, получающих усиленную химиотерапию острого лимфобластного лейкоза: результаты стратифицированного по риску рандомизированного исследования лечения центральной нервной системы MRC UKALL XI (ISRC TN 16757172). Br J Haematol . 2004, январь, 124 (1): 33-46. [Медлайн].
Хонг Д., Гупта Р., Анклифф П., Атцбергер А., Браун Дж., Сонеджи С. и др.Инициирующие и распространяющие рак клетки при детской лейкемии, связанной с TEL-AML1. Наука . 2008 18 января. 319 (5861): 336-9. [Медлайн].
Ландье В., Бхатиа С., Эшелман Д.А., Форте К.Дж., Суини Т., Хестер А.Л. и др. Разработка основанных на оценке рисков руководящих принципов для детей, переживших рак: Долгосрочные последующие рекомендации Детской онкологической группы Комитета по поздним эффектам детской онкологической группы и сестринской дисциплины. Дж Клин Онкол . 2004 15 декабря.22 (24): 4979-90. [Медлайн].
le Viseur C, Hotfilder M, Bomken S, Wilson K, Röttgers S, Schrauder A, et al. При остром лимфобластном лейкозе у детей бласты на разных стадиях иммунофенотипического созревания обладают свойствами стволовых клеток. Раковые клетки . 2008 июл 8. 14 (1): 47-58. [Медлайн]. [Полный текст].
Mulcahy N. FDA одобрило иматиниб для лечения ВСЕХ педиатрических препаратов. Медицинские новости Medscape. 25 января 2013 года. Medscape Medical News. Доступно по адресу http: // www.medscape.com/viewarticle/778062. Доступ: 1 февраля 2013 г.
Натан П.С., Василевски-Маскер К., Янцен, Л.А. Отдаленные результаты у лиц, переживших острый лимфобластный лейкоз в детском возрасте. Гематол Онкол Клин Норт Ам . 2009 октября, 23 (5): 1065-82, vi-vii. [Медлайн].
Pui CH, Campana D, Pei D, Bowman WP, Sandlund JT, Kaste SC, et al. Лечение острого лимфобластного лейкоза у детей без облучения черепа. N Engl J Med .2009, 25 июня. 360 (26): 2730-41. [Медлайн]. [Полный текст].
Pui CH, Carroll WL, Meshinchi S, Arceci RJ. Биология, стратификация риска и терапия острых лейкозов у детей: обновленная информация. Дж Клин Онкол . 2011 10 февраля. 29 (5): 551-65. [Медлайн]. [Полный текст].
Schuitema I, Deprez S, Van Hecke W., Daams M, Uyttebroeck A, Sunaert S, et al. Ускоренное старение, снижение целостности белого вещества и связанная с ним нейропсихологическая дисфункция через 25 лет после детских лимфоидных злокачественных новообразований. Дж Клин Онкол . 2013 20 сентября. 31 (27): 3378-88. [Медлайн].
Schultz KR, Pullen DJ, Sather HN, Shuster JJ, Devidas M, Borowitz MJ, et al. Классификация острого лимфобластного лейкоза детского предшественника В-предшественника на основе риска и реакции: комбинированный анализ прогностических маркеров из группы детской онкологии (POG) и детской онкологической группы (CCG). Кровь . 2007 г., 1 февраля. 109 (3): 926-35. [Медлайн]. [Полный текст].
Teachey DT, Hunger SP.Прогнозирование риска рецидива при остром лимфобластном лейкозе у детей. Br J Haematol . 2013 Сентябрь 162 (5): 606-20. [Медлайн].
Томас Д.А., Фадерл С., Кортес Дж., О’Брайен С., Джайлс Ф.Дж., Корнблау С.М. и др. Лечение острого лимфоцитарного лейкоза с положительной хромосомой в Филадельфии с помощью гипер-CVAD и мезилата иматиниба. Кровь . 2004 15 июня. 103 (12): 4396-407. [Медлайн].
Гор Л., Кернс П.Р., де Мартино М.Л., Ли, Де Соуза, Калифорния, Бертран И. и др.Дазатиниб у педиатрических пациентов с хроническим миелоидным лейкозом в хронической фазе: результаты исследования фазы II. Дж Клин Онкол . 1 мая 2018 г. 36 (13): 1330-1338. [Медлайн].