Луб | это… Что такое Луб?
флоэма, сложная ткань высших растений, служащая для проведения органических веществ к различным органам. Л. выполняет также запасающую, нередко механическую и отчасти выделительную функции. В соответствии с многообразием функций для Л. характерно наличие различных элементов: ситовидных элементов, клеток запасающей и кристаллоносной лубяной паренхимы, лубяных волокон (См. Лубяные волокна) и склереид (См. Склереиды), радиальной паренхимы сердцевинных лучей. У некоторых растений в Л. имеются Млечники и смоляные ходы. Ситовидные элементы у покрытосеменных представлены ситовидными трубками. Они состоят из продольного ряда клеток (члеников), сообщающихся «ситечками» — тонкими участками в стенках со сквозными отверстиями. В протопластах ситовидных элементов рано отмирают ядра, цитоплазма становится вполне проницаемой для растворённых в воде органических веществ и минеральных солей; оболочка целлюлозная, часто с утолщением. Скорость передвижения пластических веществ по Л.
Запасающая ткань в Л. представлена клетками осевой лубяной и лучевой паренхимы. Лубяная паренхима образована удлинёнными клетками (веретеновидная паренхима) или короткими, соединёнными в вертикальный тяж (тяжевая паренхима). Клетки сердцевинных лучей образуются особыми инициальными клетками камбия (См. Камбий) и могут быть вытянуты в радиальном или в вертикальном направлении.
Лит.: Раздорский В. Ф., Анатомия растений, М., 1949; Яценко-Хмелевский А. А., Краткий курс анатомии растений, М., 1961; Эсау К., Анатомия растений, М., 1969.
P. П. Барыкина.
П. Барыкина.
Анатомическое строение луба липы (поперечный срез): с. т. — ситовидные трубки; с. п. — ситовидные пластинки; сд — содержимое ситовидных трубок; с. к. — сопровождающие клетки; л. в. — лубяные волокна; п. с. л. — первичный сердцевинный луч; в. с. л. — вторичный сердцевинный луч; к. п. — кристаллоносные паренхимные клетки; кр — кристаллы оксалата кальция; др — друзы оксалата кальция; з. п. — клетки запасающей паренхимы.
Состав, строение и функции луба
Человечество пользовалось древесиной на протяжении тысячелетий. Ее использовали для разных целей, главным образом, как источник топлива. Также древесина является отличным строительным материалом, из нее создают инструменты, оружие, мебель, тару, произведения искусства, бумагу.
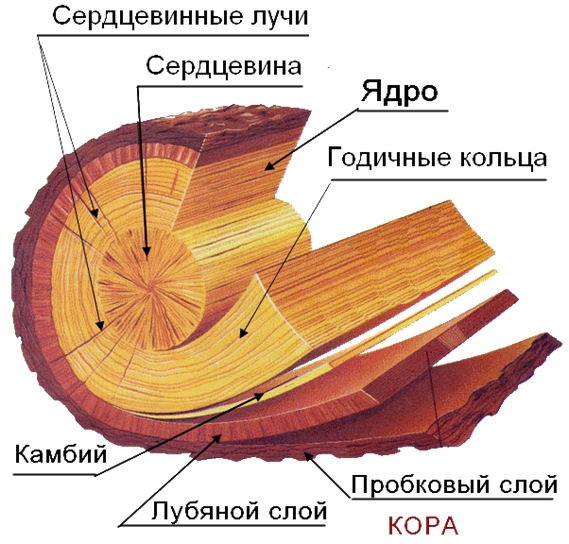
Вследствие наличия годичных колец, которые во время роста, а также в результате сезонных колебаний температуры или уровня влажности, формируют в своём стволе большинство видов деревьев, ученые могут довольно точно определить регион, в котором росло дерево. Ежегодный мониторинг изменения ширины годичных колец и анализ содержания в них некоторых изотопов элементов, дает возможность более детально изучить состояние климата и атмосферы в древние времена.
Ежегодный мониторинг изменения ширины годичных колец и анализ содержания в них некоторых изотопов элементов, дает возможность более детально изучить состояние климата и атмосферы в древние времена.
Как образуется древесина?
Древесина – это одна из составляющих сосудисто-волокнистого пучка, она противопоставлена еще одной важной части пучка, формирующейся из того же прокамбия или камбия — лубу, или флоэме. В процессе формирования сосудисто-волокнистых пучков из прокамбия возможно два варианта событий:
- все прокамбиальные клетки становятся элементами древесины и луба с образованием так называемых замкнутых пучков. Этот процесс характерен для высших споровых, однодольных и некоторых двудольных растений
- на границе между древесиной и лубом остаётся прослойка деятельной ткани, которую называют камбий. При этом формируются пучки открытые, что характерно для двудольных и голосеменных растений.
При первом варианте событий количество древесины не изменяется, и растение не может утолщаться. Если развитие идет по второму пути, то вследствие работы камбия ежегодно объем древесины увеличивается, и ствол растения медленно становится толще. У древесных пород российского региона древесина находится ближе к центру (оси) дерева, а луб — находится ближе к окружности (периферии). Ряд других растений имеет несколько иное взаимное расположение древесины и луба.
Если развитие идет по второму пути, то вследствие работы камбия ежегодно объем древесины увеличивается, и ствол растения медленно становится толще. У древесных пород российского региона древесина находится ближе к центру (оси) дерева, а луб — находится ближе к окружности (периферии). Ряд других растений имеет несколько иное взаимное расположение древесины и луба.
Именно деление клеток камбия в стебеле обеспечивает его рост в толщину. В процессе деления камбиальных клеток ? дочерних образующихся клеток отделяется в древесину, а ? – в луб. По этой причине прирост очень заметен в древесине. Камбий делится не равномерно, этот процесс зависит от сезона. В весенне – летний период деление активное, вследствие чего формируются крупные клетки, к осени деление замедляется, и формируются мелкие клетки. Зимой же камбий не делится. Таким образом, обеспечивается годичный прирост древесины, который хорошо заметен у многих деревьев, и называют его годичным кольцом. По количеству годичных колец специалисты вычисляют возраст побега и целого дерева.
Древесина имеет в своем составе уже отмершие клеточные элементы с одеревеневшими, главным образом, толстыми оболочками. Состав луба, напротив, представлен элементами живых клеток, с живой протоплазмой, клеточным соком и тонкой неодеревеневшей оболочкой. В тоже время, в лубе могут попадаться элементы мёртвые, толстостенные и одеревеневшие.
Обе составляющие сосудисто-волокнистого пучка имеют и еще одно физиологическое отличие. По древесине из земли к листьям движется сырой сок, который является водой с растворёнными в ней полезными веществами. А вот по лубу вниз течет пластический сок.
Процесс одеревенения клеточных оболочек характеризуется пропитыванием целлюлозной оболочки специальными веществами, которые объединены под общим названием лигнин. Наличие лигнина и вместе с тем одеревенение оболочки можно легко определить с помощью определенных реакций. Вследствие одеревенения, растительные оболочки растут в толщину, твердеют. В тоже время при лёгкой проницаемости для воды они утрачивают способности поглощать воду и разбухать.
Строение луба
Флоэма — то же, что и луб. Она является проводящей тканью сосудистых растений. Именно по ней осуществляется транспортировка продуктов фотосинтеза к разным частям растения, где они используются или накапливаются.
В стеблях большей части растений древесный луб находится снаружи по отношению к ксилеме, a в листьях — обращен к нижней стороне жилок листовой пластинки. Проводящие пучки корней имеют чередующиеся тяжи флоэмы и ксилемы.
Луб дерева по происхождению делится на:
- первичный, дифференциирующийся из прокамбия
- вторичный, дифференциирующийся из камбия.
Главное отличие первичной флоэмы от вторичной заключается в полном отсутствии у первой сердцевинных лучей. Однако клеточный состав и первичной, и вторичной флоэмы идентичен. В их составе присутствуют клетки различной морфологии, и выполняют разные функции:
- ситовидные элементы (клетки, трубки и клетки-спутницы). Эти элементы обеспечивают главный транспорт
- склеренхимные элементы (склереиды и волокна), отвечают за опорную функцию
- паренхимные элементы (паренхимные клетки), отвечают за ближний радиальный транспорт.


Ситовидные трубки живут совсем немного. Зачастую период их жизни не превышает 2-3 года, очень редко они доживают до 10-15 лет. Отмершие регулярно заменяют новые. Ситовидные трубки занимают немного места в лубе и чаще всего соединены в пучки. Помимо таких пучков в лубе присутствуют клетки механической ткани — лубяные волокна, а также клетки основной ткани.
Функции луба
Одной из главных функций, которые выполняет луб молодой, является флоэмный транспорт сока. Этот сок является раствором углеводов (у древесных растений — это главным образом сахароза). Углеводы – это продукты фотосинтеза, в довольно высокой концентрации — 0,2—0,7 моль/литр (примерно от 7 до 25 %). Кроме углеводов в состав сока входят и другие ассимиляты и метаболиты (аминокислоты и фитогормоны) в намного меньших количествах. Скорость транспорта достигает десятков сантиметров в час, что существенно выше скорости диффузии.
Флоэмный сок двигается от органов-доноров, в которых осуществляется процесс фотосинтеза к акцепторам — органам или областям, в которых эти продукты фотосинтеза используются или откладываются на потом. Очень интенсивно потребляются ассимиляты в корневой системе, верхушках побегов, растущих листьях, репродуктивных органах. Многие растения имеют особые органы запасания — луковицы, клубни и корневища, которые выполняют роль акцепторов.
Очень интенсивно потребляются ассимиляты в корневой системе, верхушках побегов, растущих листьях, репродуктивных органах. Многие растения имеют особые органы запасания — луковицы, клубни и корневища, которые выполняют роль акцепторов.
Луб липы – это внутренний слой коры, который имеет светло- желтую окраску. Его задача – обеспечить прочность стебля. Лубяной слой довольно проблематично разорвать по ширине, однако вдоль стебля он легко распадается на тонкие волокна большой длины.
Лубяная часть стебля часто применяется в хозяйстве, к примеру, луб липы знаменит тем, что из него изготавливают рогожу и мочалки.
Отметим, что если кору на дереве по кругу перерезать до слоя древесины, то органические вещества больше не будут транспортироваться к корням, и дерево через время погибнет.
Биология рака яичников: новые возможности трансляции
1. Goff BA, et al. Диагностика рака яичников. Рак. 2000;89:2068. [PubMed] [Google Scholar]
2. Zebrowski BK, et al. Заметно повышенный уровень сосудистого эндотелиального фактора роста при злокачественном асците.
3. Мезиано С., Феррара Н., Джаффе Р. Роль фактора роста эндотелия сосудов при раке яичников: ингибирование образования асцита путем иммунонейтрализации. Являюсь. Дж. Путь. 1998;153:1249. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Numnum TM, et al. Применение бевацизумаба для паллиативного лечения симптоматического асцита у пациентов с рефрактерным раком яичников. Гинекол. Онкол. 2006; 102:425. [PubMed] [Google Scholar]
5. Берек Дж.С. В: Практическая гинекологическая онкология, 4-е изд., гл. 11 Рак яичников. Берек Дж.С., Хакер Н.Ф., редакторы. Филадельфия: Липпинкотт Уильямс и Уилкинс; 2005. стр. 443–511. [Google Scholar]
6. Armstrong DK, et al. Цисплатин и паклитаксел внутрибрюшинно при раке яичников. Н. англ. Дж. Мед. 2006; 354:34–43. [PubMed] [Академия Google]
7. Фили К.М., Уэллс М. Предвестники злокачественных новообразований эпителия яичников. Гистопатология. 2001; 38:87. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
8.
Чжан С. и др. Идентификация и характеристика клеток, инициирующих рак яичников, из первичных опухолей человека. Рак рез. 2008;68:4311–4320. [Бесплатная статья PMC] [PubMed] [Google Scholar]
В этом отчете установлен фенотип инициирующих опухоль клеток рака яичников.
9. Alvero AB, et al. Молекулярное фенотипирование стволовых клеток рака яичников человека раскрывает механизмы репарации и химиорезистентности. Клеточный цикл. 2009 г.;8:188–169. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. Jacobs IJ, et al. Клональное происхождение эпителиального рака яичников: анализ по потере гетерозиготности, мутации р53 и инактивации Х-хромосомы. J. Natl Cancer Inst. 1992; 84: 1793–1798. [PubMed] [Google Scholar]
11. Bast RC, Jr, Mills GB. В: Молекулярная основа рака. 3-е изд. Мендельсон Дж., Хоули П., Исраэль М., Грей Дж., Томпсон С., редакторы. Филадельфия: WB Saunders Co .; 2008. стр. 441–455. [Академия Google]
12. Iwabuchi H, et al. Генетический анализ доброкачественных, низкодифференцированных и высокодифференцированных опухолей яичников. Рак рез. 1995; 55: 6172–6180. [PubMed] [Google Scholar]
Iwabuchi H, et al. Генетический анализ доброкачественных, низкодифференцированных и высокодифференцированных опухолей яичников. Рак рез. 1995; 55: 6172–6180. [PubMed] [Google Scholar]
13. Risch HA, et al. Популяционные частоты мутаций BRCA1 и BRCA2 и пенетрантность рака: когортное исследование в Онтарио, Канада. J. Natl Cancer Inst. 2006; 98: 1675–1677. [PubMed] [Google Scholar]
14. Cramer DW, et al. Генитальное воздействие талька и риск рака яичников. Междунар. Дж. Рак. 1999; 81: 351–356. [PubMed] [Академия Google]
15. Мускат Ю.Э., Гунчарек М.С. Использование перинеального талька и рак яичников: критический обзор. Евро. J. Рак Prev. 2006; 62: 358–360. [Google Scholar]
16.
Колер М.Ф. и соавт. Спектр мутаций и частота аллельных делеций гена р53 при раке яичников. J. Natl Cancer Inst. 1993; 85: 1513–1519. [PubMed] [Академия Google]
В этой статье показано, что рак яичников подвергается спонтанной мутации.
17. Berchuck A, et al. Сверхэкспрессия р53 не характерна для доброкачественных и пограничных эпителиальных опухолей яичников на ранней стадии. Гинекол. Онкол. 1994;52:232–236. [PubMed] [Google Scholar]
Гинекол. Онкол. 1994;52:232–236. [PubMed] [Google Scholar]
18. Berchuck A, et al. Ген-супрессор опухоли p53 часто изменяется при гинекологическом раке. Являюсь. Дж. Обст. Гинекол. 1994; 170: 246–252. [PubMed] [Google Scholar]
19. Havrilesky L, et al. Прогностическое значение мутации р53 и гиперэкспрессии р53 при прогрессирующем эпителиальном раке яичников: исследование группы гинекологической онкологии. Дж. Клин. Онкол. 2003; 21:3814–3825. [PubMed] [Google Scholar]
20.
Холл Дж. и др. Критическая оценка р53 как прогностического маркера рака яичников. Эксп. Преподобный Мол. Мед. 2004; 12:1–20. [PubMed] [Академия Google]
Вдумчивый и тщательный обзор прогностического значения р53 при раке яичников.
21. Buller RE, et al. Фаза I / II испытания замены гена rAd/ p53 (SCH58500) при рецидивирующем раке яичников. Ген Рака Ther. 2002; 9: 553–566. [PubMed] [Google Scholar]
22. Vasey PA, et al. Испытание фазы I внутрибрюшинной инъекции E1B -55-kd-ген-делетированного аденовируса ONYX-015 (dl1520), вводившейся с 1 по 5 день каждые 3 недели у пациентов с рецидивирующим/рефрактерным эпителиальным раком яичников. Дж. Клин. Онкол. 2002; 15: 1562–1569. [PubMed] [Академия Google]
Дж. Клин. Онкол. 2002; 15: 1562–1569. [PubMed] [Академия Google]
23. Кодзима К. и др. Антагонисты MDM2 индуцируют p53-зависимый апоптоз при ОМЛ: последствия для терапии лейкемии. Кровь. 2005;106:3150–3159. [Бесплатная статья PMC] [PubMed] [Google Scholar]
24.
Ю Ю и др. Методы в энзимологии: регуляторы и эффекторы малых ГТФаз. В: Balch WE, Der C, Hall A, редакторы. Рас белки. Том. 407. Нью-Йорк: академический; 2006. стр. 455–467. [Google Scholar]
Всесторонний обзор роли DIRAS3 (ARHI) в развитии рака яичников.
25. Cvetkovic D, et al. Измененная экспрессия и потеря гетерозиготности Ген LOT1 при раке яичников. Гинекол. Онкол. 2004; 95: 449–455. [PubMed] [Google Scholar]
26. Feng W, et al. Импринтированные гены-супрессоры опухолей ARHI и PEG3 чаще всего подавляются при раке яичников человека из-за потери гетерозиготности и метилирования промотора. Рак. 2008; 112:1489–1502. [PubMed] [Google Scholar]
27. Chen MY, et al.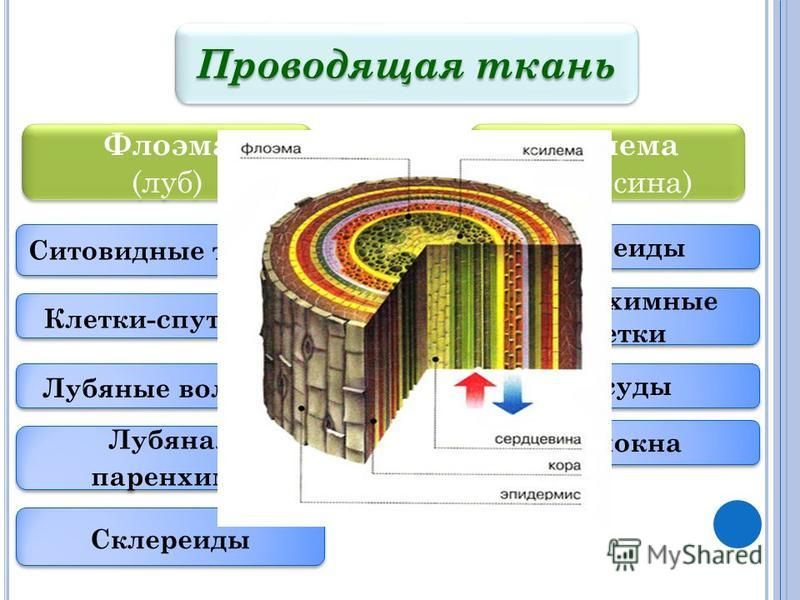 Синергетическое ингибирование роста клеток рака яичников деметилирующими агентами и ингибиторами гистондеацетилазы. проц. амер. доц. Рак рез. 2007;681 [Google Академия]
Синергетическое ингибирование роста клеток рака яичников деметилирующими агентами и ингибиторами гистондеацетилазы. проц. амер. доц. Рак рез. 2007;681 [Google Академия]
28. Mackay H, et al. Испытание фазы II ингибитора гистондеацетилазы белиностата (PSC101) у пациентов с резистентными к платине эпителиальными опухолями яичников и микропапиллярными/пограничными (LMP) опухолями яичников. Испытание консорциума фазы II PMH. Дж. Клин. Онкол. 2008;26 Дополнение:5518. [Google Scholar]
29. Balch C, et al. Эпигенетика лекарственной устойчивости и ресенсибилизации рака яичников. Являюсь. Дж. Обст. Гинекол. 2004; 191:1552–1572. [PubMed] [Google Scholar]
30. Bast RC, et al. Исследование фазы IIa последовательного режима с использованием азацитидина для устранения резистентности платины к карбоплатину у пациентов с резистентным или рефрактерным к платине эпителиальным раком яичников. Дж. Клин. Онкол. 2008;26 Дополнение:3500. [Академия Google]
31. Рубин С.К., и соавт. BRCA1, BRCA2, и наследственные неполипозные мутации гена колоректального рака в невыбранной популяции рака яичников: связь с семейным анамнезом и значение для генетического тестирования. Являюсь. Дж. Обст. Гинекол. 1998; 178: 670–677. [PubMed] [Google Scholar]
BRCA1, BRCA2, и наследственные неполипозные мутации гена колоректального рака в невыбранной популяции рака яичников: связь с семейным анамнезом и значение для генетического тестирования. Являюсь. Дж. Обст. Гинекол. 1998; 178: 670–677. [PubMed] [Google Scholar]
32. Lancaster JM, et al. Мутации BRCA2 при первичном раке молочной железы и яичников. Природа Жене. 1996; 13: 238–240. [PubMed] [Google Scholar]
33. Бойд Дж. В: Рак яичников 5. Шарп Ф., Блэкетт Т., Берек Дж., Баст Р., редакторы. Оксфорд: Isis Medical Media; 1998. С. 3–16. [Google Scholar]
34. Chetrit A, Hirsh-Yechezkel G, Ben-David Y, Lubin F, Friedman E. Влияние мутаций BRCA 1/2 на долгосрочную выживаемость пациентов с раком яичников: национальный израильский исследование рака яичников. Дж. Клин. Онкол. 2008; 26:20–25. [PubMed] [Google Scholar]
35. Moynahan ME, et al. Направленная гомологией репарация ДНК, устойчивость к митомицину-с и стабильность хромосом восстанавливаются при коррекции мутации Brca1 . Рак рез. 2001;61:4842–4850. [PubMed] [Академия Google]
Рак рез. 2001;61:4842–4850. [PubMed] [Академия Google]
36. Народ С.А., Фоулкс В.Д. BRCA1 и BRCA2 , 1994 г. и позже. Природа Преподобный Рак. 2004; 4: 665–676. [PubMed] [Google Scholar]
37. Edwards SL, et al. Устойчивость к терапии обусловлена внутригенной делецией BRCA2 . Природа. 2008; 451:1111–1115. [PubMed] [Google Scholar]
38. Sakai W, et al. Вторичные мутации как механизм резистентности к цисплатину в BRCA2 -мутированных раках. Природа. 2008; 451:1116–1120. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Дрю И., Калверт Х. Потенциал ингибиторов PARP при генетическом раке груди и яичников. Анна. Академик Нью-Йорка науч. 2008; 1138: 126–145. [PubMed] [Google Scholar]
40.
Яп Т.А., Карден К.Т., Кэй С.Б. Помимо химиотерапии: таргетная терапия рака яичников. Природа Преподобный Рак. 2009; 9: 167–181. [PubMed] [Академия Google]
Тщательный и актуальный обзор молекулярной терапии рака яичников.
41. Hennessey B, et al. Статус BRCA при раке яичников. проц. амер. соц. клин. Онкол. (в печати) [Google Scholar]
Hennessey B, et al. Статус BRCA при раке яичников. проц. амер. соц. клин. Онкол. (в печати) [Google Scholar]
42. Umayahara K, et al. В: Рак яичников 5. Шарп Ф., Блэкетт Т., Берек Дж., Баст Р., редакторы. Оксфорд: Isis Medical Media; 1998. С. 17–23. [Google Scholar]
43.
Джазаери А.А. и соавт. Профили экспрессии генов BRCA1 -сцепленного, BRCA2 -сцепленного и спорадического рака яичников. J. Natl Cancer Inst. 2002; 13: 990–1000. [PubMed] [Академия Google]
Эта провокационная статья предполагает, что спорадический рак яичников является либо BRCA1, либо BRCA2-подобным.
44. Eder AM, et al. Атипичная PKC ι способствует плохому прогнозу из-за потери апикально-базальной полярности и гиперэкспрессии циклина E при раке яичников. проц. Натл акад. США. 2005;102:12519–12524. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Zhang L, et al. МикроРНК демонстрируют высокую частоту геномных изменений при раке человека. проц. Натл акад.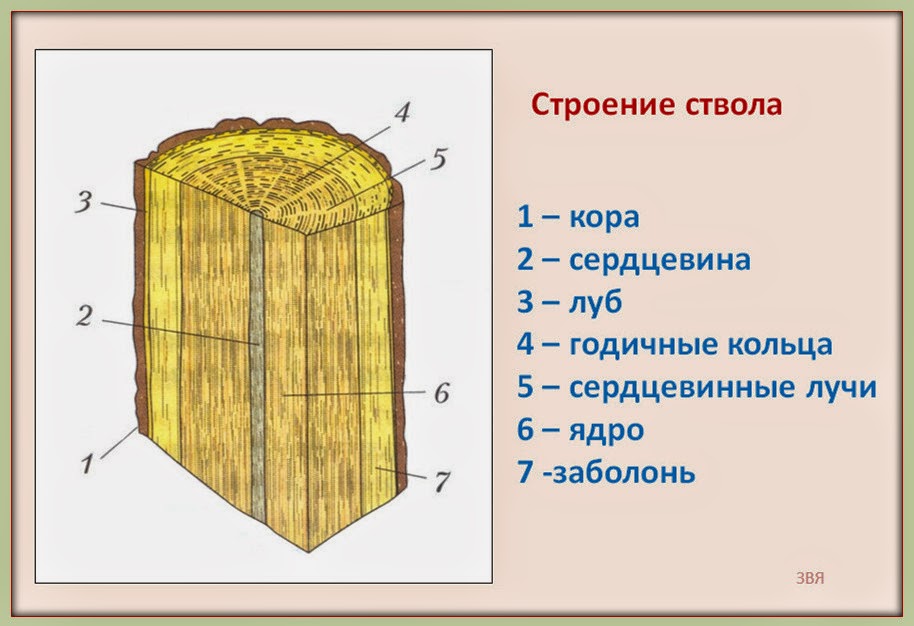 науч. США. 2003; 103:9136–9141. [Бесплатная статья PMC] [PubMed] [Google Scholar]
науч. США. 2003; 103:9136–9141. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Tangir J, et al. Частая микросателлитная нестабильность при эпителиальных пограничных опухолях яичников. Рак рез. 1996; 56: 2501–2505. [PubMed] [Google Scholar]
47. Rodabaugh KJ, et al. Детальное картирование делеций хромосомных изменений 9p и p16 в пограничных и инвазивных эпителиальных опухолях яичников человека. Онкоген. 1995;11:1249–1254. [PubMed] [Google Scholar]
48. Berchuck A, et al. Сверхэкспрессия р53 не характерна для доброкачественных и пограничных эпителиальных опухолей яичников на ранней стадии. Гинекол. Онкол. 1994; 52: 232–236. [PubMed] [Google Scholar]
49. Iwabuchi H, et al. Генетический анализ доброкачественных, низкодифференцированных и высокодифференцированных опухолей яичников. Рак рез. 1995; 55: 6172–6180. [PubMed] [Google Scholar]
50. Abu-Jawdeh GM, et al. Экспрессия рецепторов эстрогена является общей чертой пограничных опухолей яичников. Гинекол. Онкол. 1996;60:301–307. [PubMed] [Google Scholar]
Гинекол. Онкол. 1996;60:301–307. [PubMed] [Google Scholar]
51.
Лю Дж. и др. Генетически определенная модель рака яичников человека. Рак рез. 2004; 64: 1655–1663. [PubMed] [Академия Google]
В этой статье показано, что фенотип яичников можно индуцировать в ксенотрансплантатах путем трансфекции нормальных эпителиальных клеток яичников человека Т-антигеном SV40, теломеразой и мутантным Ras.
52. Cheng KW, et al. Новая роль Rab GTPases в раке и заболеваниях человека. Рак рез. 2005;65:2516–2519. [PubMed] [Академия Google]
53. Gautschi O, et al. Киназы Aurora как мишени противораковых препаратов. клин. Рак рез. 2008; 14:1639–1648. [PubMed] [Google Scholar]
54. Li K, et al. Модуляция передачи сигналов Notch антителами, специфичными к внеклеточной регуляторной области Notch4. Дж. Биол. хим. 2008; 283:8046–8054. [PubMed] [Google Scholar]
55. Schilder RJ, et al. Исследование фазы II гефитиниба у пациентов с рецидивирующей или персистирующей карциномой яичников или первичной перитонеальной карциномой и оценка мутаций рецептора эпидермального фактора роста и иммуногистохимической экспрессии: групповое исследование гинекологической онкологии. клин. Рак рез. 2005;11:5539–5548. [PubMed] [Google Scholar]
клин. Рак рез. 2005;11:5539–5548. [PubMed] [Google Scholar]
56. Gordon AN, et al. Эффективность и безопасность эрлотиниба гидрохлорида, ингибитора тирозинкиназы рецептора эпидермального фактора роста (HER1/EGFR), у пациентов с распространенным раком яичников: результаты многоцентрового исследования фазы II. Междунар. Дж. Гинекол. Рак. 2005; 15: 785–792. [PubMed] [Google Scholar]
57. Heinemann V, Stintzing S, Kirchner T, Boeck S, Jung A. Клиническая значимость EGFR- и KRAS-статуса у пациентов с колоректальным раком, получавших моноклональные антитела, направленные против EGFR. Лечение рака. Версия 2009 г.;35:262–271. [PubMed] [Google Scholar]
58. Bookman MA, et al. Оценка моноклонального гуманизированного антитела к HER2, трастузумаба, у пациентов с рецидивирующей или рефрактерной карциномой яичников или первичной перитонеальной карциномой со сверхэкспрессией HER2: исследование фазы II группы гинекологической онкологии. Дж. Клин. Онкол. 2003; 21: 283–290. [PubMed] [Google Scholar]
59. Hu L, Hofmann J, Lu Y, Mills GB, Jaffe RB. Ингибирование фосфатидилинозитол-3′-киназы повышает эффективность паклитаксела в in vitro и in vivo модели рака яичников. Рак рез. 2002; 62: 1087–1092. [PubMed] [Google Scholar]
Hu L, Hofmann J, Lu Y, Mills GB, Jaffe RB. Ингибирование фосфатидилинозитол-3′-киназы повышает эффективность паклитаксела в in vitro и in vivo модели рака яичников. Рак рез. 2002; 62: 1087–1092. [PubMed] [Google Scholar]
60. Raynaud FL, et al. Фармакологическая характеристика мощного ингибитора фосфатидилинозитид-3-киназ класса I. Рак рез. 2007; 67: 5840–5850. [PubMed] [Google Scholar]
61. Rosen DG, et al. Роль конститутивно активного преобразователя сигнала и активатора транскрипции 3 в онкогенезе и прогнозе яичников. Рак. 2006; 107: 2730–2740. [PubMed] [Академия Google]
62. Burke WM, et al. Ингибирование конститутивно активного Stat3 подавляет рост клеток рака яичников и молочной железы человека. Онкоген. 2001; 20:7925–7934. [PubMed] [Google Scholar]
63. Duan Z, et al. 8-бензил-4-оксо-8-азабицикло[3.2.1]окт-2-ен-6,7-дикарбоновая кислота (SD-1008), новый ингибитор янус-киназы 2, повышает чувствительность к химиотерапии клеток рака яичников человека. Мол. Фармакол. 2007; 72: 1137–1145. [PubMed] [Google Scholar]
Мол. Фармакол. 2007; 72: 1137–1145. [PubMed] [Google Scholar]
64. McMurray JS. Новый низкомолекулярный ингибитор Stat 3. хим. биол. 2006; 13:1123–1124. [PubMed] [Академия Google]
65. Murph M, et al. О пауках и крабах: появление лизофосфолипидов и путей их метаболизма в качестве мишеней для терапии рака. клин. Рак рез. 2006; 12:6598–6602. [PubMed] [Google Scholar]
66. Beck HP, et al. Открытие мощных антагонистов LPA2 (EDG4) в качестве потенциальных противоопухолевых средств. биоорг. Мед. хим. лат. 2008;18:1037–1041. [PubMed] [Google Scholar]
67. Lin YG, et al. Куркумин ингибирует рост опухоли и ангиогенез при раке яичников, воздействуя на путь ядерного фактора-κB. клин. Рак рез. 2007; 13:3423–3430. [PubMed] [Академия Google]
68. Samanta AK, Huang HJ, Bast RC, Jr, Liao W. Сверхэкспрессия MEKK3 придает устойчивость к апоптозу посредством активации NF κB. Дж. Биол. хим. 2004; 279:7576–7583. [PubMed] [Google Scholar]
69. Häcker H, Karin M. Регуляция и функция IKK и киназ, связанных с IKK. Наука СТКЭ. 2006;357:re 13. [PubMed] [Google Scholar]
Регуляция и функция IKK и киназ, связанных с IKK. Наука СТКЭ. 2006;357:re 13. [PubMed] [Google Scholar]
70. Yang J, et al. Существенная роль MEKK3 в TNF-индуцированной активации NF-κB. Натура Иммунол. 2001; 2: 620–624. [PubMed] [Академия Google]
71. Карин М. Ядерный фактор κB в развитии и прогрессировании рака. Природа. 2006; 441:431–436. [PubMed] [Google Scholar]
72. См. HT, Kavanagh JJ, Hu W, Bast RC., Jr Таргетная терапия эпителиального рака яичников: текущее состояние и перспективы на будущее. Междунар. Дж. Гинекол. Рак. 2004; 13: 701–734. [PubMed] [Google Scholar]
73. Suh DS, Yoon MS, Choi KU, Kim JY. Значение сверхэкспрессии E2F-1 при раке яичников. Междунар. Дж. Гинекол. Рак. 2008; 18: 492–498. [PubMed] [Академия Google]
74. Reimer D, et al. Экспрессия семейства факторов транскрипции E2 и ее клиническое значение при раке яичников. Анна. Академик Нью-Йорка науч. 2006; 1091: 270–286. [PubMed] [Google Scholar]
75. Berchuck A, et al. Регуляция роста нормальных эпителиальных клеток яичников и клеточных линий рака яичников с помощью трансформирующего фактора роста-β Am. Дж. Обст. Гинекол. 1992; 166: 676–684. [PubMed] [Google Scholar]
Регуляция роста нормальных эпителиальных клеток яичников и клеточных линий рака яичников с помощью трансформирующего фактора роста-β Am. Дж. Обст. Гинекол. 1992; 166: 676–684. [PubMed] [Google Scholar]
76. Sunde JS, et al. Профилирование экспрессии идентифицирует измененную экспрессию генов, которые способствуют ингибированию передачи сигналов трансформирующего фактора роста-β при раке яичников. Рак рез. 2006;66:8404–8412. [PubMed] [Академия Google]
77. Фуллер А.Ф., младший, Гай С., Будзик Г.П., Донахью П.К. Мюллеров ингибирующее вещество ингибирует рост колоний клеточной линии карциномы яичника человека. Дж. Клин. Эндокринол. Метабол. 1982; 54: 1051–1055. [PubMed] [Google Scholar]
78. Szotek PP, et al. Боковая популяция рака яичников определяет клетки с характеристиками, подобными стволовым клеткам, и чувствительностью к мюллеровским ингибирующим веществам. проц. Натл акад. науч. США. 2006; 103:11154–11159. [Бесплатная статья PMC] [PubMed] [Google Scholar]
79. Пьеретти-Ванмарке Р. и соавт. Мюллеровское ингибирующее вещество увеличивает субклинические дозы химиотерапевтических агентов для ингибирования рака яичников человека и мыши. проц. Натл акад. науч. США. 2006; 103:17426–17431. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Пьеретти-Ванмарке Р. и соавт. Мюллеровское ингибирующее вещество увеличивает субклинические дозы химиотерапевтических агентов для ингибирования рака яичников человека и мыши. проц. Натл акад. науч. США. 2006; 103:17426–17431. [Бесплатная статья PMC] [PubMed] [Google Scholar]
80. Reed J, et al. Значение экспрессии белка рецептора Fas при эпителиальном раке яичников. Гум. Патол. 2005; 36: 971–976. [PubMed] [Google Scholar]
81. Kar R, et al. Роль регуляторов апоптоза в эпителиальном раке яичников человека. Рак биол. тер. 2007; 6: 1101–1105. [PubMed] [Академия Google]
82. Schuyer M, et al. Снижение экспрессии BAX связано с плохим прогнозом у больных эпителиальным раком яичников: многофакторный анализ TP53, p21, BAX и BCL-2 . бр. Дж. Рак. 2001; 85: 1359–1367. [Бесплатная статья PMC] [PubMed] [Google Scholar]
83. Lancaster JM, et al. Высокая экспрессия лиганда фактора некроза опухоли, индуцирующего апоптоз, связана с благоприятной выживаемостью при раке яичников. клин. Рак рез. 2003; 9: 762–766. [PubMed] [Академия Google]
клин. Рак рез. 2003; 9: 762–766. [PubMed] [Академия Google]
84. De la Torre FJ, et al. Апоптоз в эпителиальных опухолях яичников: прогностическое значение клинических и гистопатологических факторов и его связь с иммуногистохимической экспрессией регуляторных белков апоптоза (p53, bcl-2 и bax) Eur. Дж. Обст. Гинекол. Воспр. биол. 2007; 130:121–128. [PubMed] [Google Scholar]
85. Liang XH, et al. Индукция аутофагии и ингибирование онкогенеза беклином 1 . Природа. 1999; 402: 672–676. [PubMed] [Академия Google]
86.
Лу Зи и др. Новый ген-супрессор опухоли ARHI индуцирует аутофагию и покой опухоли в ксенотрансплантатах рака яичников. Дж. Клин. Вкладывать деньги. 2008; 118:3917–3929. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Первоначальный отчет о связи аутофагии и покоя опухоли.
87. Ren J, et al. Лизофосфатидная кислота конститутивно вырабатывается мезотелиальными клетками брюшины человека и усиливает адгезию, миграцию и инвазию клеток рака яичников. Рак рез. 2006;15:3006–3014. [PubMed] [Академия Google]
Рак рез. 2006;15:3006–3014. [PubMed] [Академия Google]
88. Фишман Д.А. и соавт. Лизофосфатидная кислота способствует активации матриксной металлопротеиназы (ММП) и ММП-зависимой инвазии в клетки рака яичников. Рак рез. 2001; 1:3194–3199. [PubMed] [Google Scholar]
89. Fang X, et al. Механизмы продукции цитокинов, индуцированной лизофосфатидной кислотой, в клетках рака яичников. Дж. Биол. хим. 2004; 279:9653–9661. [PubMed] [Google Scholar]
90.
Суд А.К. и соавт. Опосредованная гормонами стресса инвазия клеток рака яичников. клин. Рак рез. 2006;15:369–375. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Этот отчет связывает стресс с ростом рака яичников через физиологические механизмы.
91. Барболина М.В., и соавт. Микроокружающая регуляция активности матриксной металлопротеиназы мембранного типа 1 в клетках карциномы яичника посредством индуцированной коллагеном экспрессии EGR1. Дж. Биол. хим. 2007; 16:4924–4931. [PubMed] [Google Scholar]
92. Cai KQ, et al. Выраженная экспрессия металлопротеиназ на ранних стадиях онкогенеза яичников. Мол. Карциног. 2007; 46: 130–143. [PubMed] [Академия Google]
Cai KQ, et al. Выраженная экспрессия металлопротеиназ на ранних стадиях онкогенеза яичников. Мол. Карциног. 2007; 46: 130–143. [PubMed] [Академия Google]
93. Prezas P, et al. Сверхэкспрессия генов калликреина тканей человека KLK4, 5, 6 и 7 увеличивает злокачественный фенотип клеток рака яичников. биол. хим. 2006; 387: 807–811. [PubMed] [Google Scholar]
94. Paliouras M, et al. Калликреины тканей человека: семейство биомаркеров рака. Рак Летт. 2007; 28: 61–79. [PubMed] [Google Scholar]
95. Yin BWT, Lloyd KO. Молекулярное клонирование антигена рака яичников CA125: идентификация нового муцина, MUC16. Дж. Биол. хим. 2001; 276: 27371–27375. [PubMed] [Академия Google]
96. Gubbels JA, et al. Связывание мезотелина с MUC16 представляет собой высокоаффинное, зависимое от N-гликанов взаимодействие, которое способствует перитонеальному метастазированию опухолей яичников. Мол. Рак. 2006; 5:50. [Статья бесплатно PMC] [PubMed] [Google Scholar]
97. Rump A, Morikawa Y, Tanaka M. Связывание антигена рака яичников CA125/MUC16 с мезотелином опосредует клеточную адгезию. Дж. Биол. хим. 2004; 279:9190–9198. [PubMed] [Google Scholar]
Rump A, Morikawa Y, Tanaka M. Связывание антигена рака яичников CA125/MUC16 с мезотелином опосредует клеточную адгезию. Дж. Биол. хим. 2004; 279:9190–9198. [PubMed] [Google Scholar]
98. Cannistra SA, et al. Экспрессия варианта CD44 является общей чертой эпителиального рака яичников: отсутствие связи со стандартными прогностическими факторами. Дж. Клин. Онкол. 1995;13:1912–1921. [PubMed] [Google Scholar]
99. Strobel T, Swanson L, Cannistra SA. In vivo ингибирование CD44 ограничивает внутрибрюшное распространение ксенотрансплантата рака яичников человека у голых мышей: новая роль CD44 в процессе перитонеальной имплантации. Рак рез. 1997; 57: 1228–1232. [PubMed] [Google Scholar]
100. Yoneda J, et al. Экспрессия генов, связанных с ангиогенезом, и прогрессирование рака яичников человека у голых мышей. J. Natl Cancer Inst. 1998; 90: 447–454. [PubMed] [Академия Google]
101.
Биррер М.Дж. и соавт. Сравнительный анализ геномной гибридизации на основе полногеномных олигонуклеотидов идентифицировал фактор роста фибробластов 1 как прогностический маркер для серозных аденокарцином яичников на поздних стадиях. Дж. Клин Онкол. 2007; 1: 2281–2287. [PubMed] [Академия Google]
Недавний сравнительный анализ геномной гибридизации указывает на важность FGF1 в патогенезе рака яичников.
Дж. Клин Онкол. 2007; 1: 2281–2287. [PubMed] [Академия Google]
Недавний сравнительный анализ геномной гибридизации указывает на важность FGF1 в патогенезе рака яичников.
102. Монк Б.Дж. и др. Спасательная терапия на основе бевацизумаба (rhuMAB VEGF) после нескольких предшествующих цитотоксических режимов при прогрессирующем рефрактерном эпителиальном раке яичников. Гинекол. Онкол. 2006; 102: 140–144. [PubMed] [Академия Google]
103. Камат А.А., и соавт. Метрономная химиотерапия повышает эффективность антиваскулярной терапии при раке яичников. Рак рез. 2007; 67: 281–288. [PubMed] [Google Scholar]
104. Lu C, et al. Влияние созревания сосудов на антиангиогенную терапию при раке яичников. Являюсь. Дж. Обст. Гинекол. 2008;198:477.e1–477.e9. [Бесплатная статья PMC] [PubMed] [Google Scholar]
105. Lin YG, et al. Сверхэкспрессия EphA2 связана с ангиогенезом при раке яичников. Рак. 2007; 109: 332–340. [PubMed] [Академия Google]
106. Landen CN, et al.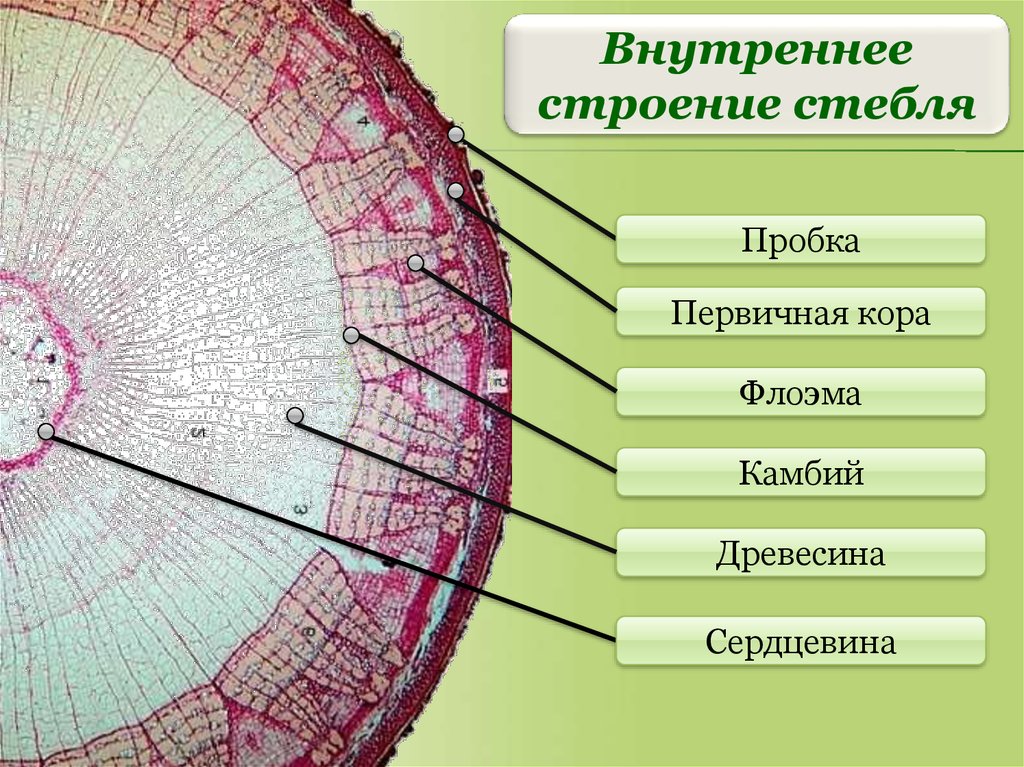 Эффективность и противососудистые эффекты снижения EphA2 с помощью агонистического антитела при раке яичников. J. Natl Cancer Inst. 2006; 98: 1558–1570. [PubMed] [Google Scholar]
Эффективность и противососудистые эффекты снижения EphA2 с помощью агонистического антитела при раке яичников. J. Natl Cancer Inst. 2006; 98: 1558–1570. [PubMed] [Google Scholar]
107.
Ланден К.Н. и др. Внутрибрюшинная доставка липосомальной миРНК для терапии распространенного рака яичников. Рак биол. тер. 2006; 5: 1708–1713. [PubMed] [Академия Google]
В этой статье показано, что нейтральные липосомы обеспечивают эффективную доставку siRNA к ксенотрансплантатам рака яичников человека.
108. Mancuso MR, et al. Быстрое возобновление роста сосудов в опухолях после отмены ингибирования VEGF. Дж. Клин. Вкладывать деньги. 2006;116:2610–2621. [Бесплатная статья PMC] [PubMed] [Google Scholar]
109. Yang G, et al. Онкоген 1 (Gro-1), регулируемый ростом хемокинов, связывает передачу сигналов RAS со старением стромальных фибробластов и онкогенезом яичников. проц. Натл акад. науч. США. 2006; 31:16472–16477. [Бесплатная статья PMC] [PubMed] [Google Scholar]
110. Milliken D, et al. Анализ хемокинов и экспрессии хемокиновых рецепторов при асците рака яичников. клин. Рак рез. 2002; 8: 1108–1114. [PubMed] [Академия Google]
Milliken D, et al. Анализ хемокинов и экспрессии хемокиновых рецепторов при асците рака яичников. клин. Рак рез. 2002; 8: 1108–1114. [PubMed] [Академия Google]
111.
Чжан Л. и др. Внутриопухолевые Т-клетки, рецидив и выживаемость при эпителиальном раке яичников. Н. англ. Дж. Мед. 2003; 16: 203–213. [PubMed] [Академия Google]
Тщательное исследование, подтверждающее прогностическое значение Т-клеточной инфильтрации при раке яичников.
112. Sato E, et al. Интраэпителиальные лимфоциты CD8 + , инфильтрирующие опухоль, и высокое соотношение CD8 + / регуляторные Т-клетки связаны с благоприятным прогнозом при раке яичников. проц. Натл акад. науч. США. 2005; 102:18538–18543. [Бесплатная статья PMC] [PubMed] [Google Scholar]
113. Jiang YP, et al. Экспрессия хемокина CXCL12 и его рецептора CXCR4 при эпителиальном раке яичников человека: независимый прогностический фактор прогрессирования опухоли. Гинекол. Онкол. 2006; 103: 226–233. [PubMed] [Google Scholar]
114. Kryczek I, et al. CXCL12 и фактор роста эндотелия сосудов синергически индуцируют неоангиогенез при раке яичников человека. Рак рез. 2005; 65: 465–472. [PubMed] [Google Scholar]
Kryczek I, et al. CXCL12 и фактор роста эндотелия сосудов синергически индуцируют неоангиогенез при раке яичников человека. Рак рез. 2005; 65: 465–472. [PubMed] [Google Scholar]
115. Curiel TJ, et al. Субпопуляции дендритных клеток по-разному регулируют ангиогенез при раке яичников человека. Рак рез. 2004; 64: 5535–5538. [PubMed] [Академия Google]
116. Kajiyana H, et al. Участие оси SDF-1α/CXCR4 в усиленном перитонеальном метастазировании эпителиального рака яичников. Междунар. Дж. Рак. 2008; 122:91–99. [PubMed] [Google Scholar]
117. Szosarek PW, et al. Экспрессия и регуляция фактора некроза опухоли α в нормальном и злокачественном эпителии яичников. Мол. Рак Тер. 2006; 5: 382–390. [PubMed] [Google Scholar]
118. Kulbe H, et al. Воспалительный цитокин фактор некроза опухоли-α генерирует аутокринную опухоль-стимулирующую сеть в эпителиальных раковых клетках яичников. Рак рез. 2007; 67: 585–59.2. [Бесплатная статья PMC] [PubMed] [Google Scholar]
119. Madhusdan S, et al. Изучение этанерцепта, ингибитора некроза опухоли α, при рецидивирующем раке яичников. Дж. Клин. Онкол. 2005; 23: 5950–5959. [PubMed] [Google Scholar]
Madhusdan S, et al. Изучение этанерцепта, ингибитора некроза опухоли α, при рецидивирующем раке яичников. Дж. Клин. Онкол. 2005; 23: 5950–5959. [PubMed] [Google Scholar]
120.
Растин Г.Дж.С. и др. Использование CA-125 в оценке клинических испытаний новых терапевтических препаратов для лечения рака яичников. клин. Рак рез. 2004; 10:3919–3926. [PubMed] [Академия Google]
Недавний обзор применения CA125 в клинических испытаниях.
121.
Менон У и др. Чувствительность и специфичность мультимодального и ультразвукового скрининга рака яичников и распределение по стадиям выявленного рака: результаты скрининга распространенности Совместного исследования скрининга рака яичников в Великобритании (UKCTOCS) Lancet Oncol. 2009; 10: 327–340. [PubMed] [Академия Google]
Первоначальные данные этого исследования позволяют предположить, что сочетание CA125 и трансвагинального УЗИ будет эффективной стратегией раннего выявления рака яичников.
122. Das PM, Bast RC., Jr Раннее выявление рака яичников. Биомаркеры Мед. 2008;2:291–303. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Биомаркеры Мед. 2008;2:291–303. [Бесплатная статья PMC] [PubMed] [Google Scholar]
123. Bouchard D, et al. Белки с мотивами сывороточно-кислых белков и рак. Ланцет Онкол. 2006; 7: 167–174. [PubMed] [Google Scholar]
124. Lu KH, et al. Отбор потенциальных маркеров эпителиального рака яичников с помощью массивов экспрессии генов и рекурсивного анализа разделения спуска. клин. Рак рез. 2004; 10:3291–3300. [PubMed] [Google Scholar]
125. Clarke CH, et al. Панель протеомных маркеров повышает чувствительность CA125 для выявления эпителиального рака яичников I стадии. Дж. Клин. Онкол. 2008;26 Дополнение:5542. [Академия Google]
126. Bast RC, et al. Оптимизация двухэтапной стратегии раннего выявления рака яичников. Встреча NCI по трансляционной науке 300, № 292. Национальный институт рака. 2008 [Google Scholar]
127. Shridhar V, et al. Генетический анализ ранних и поздних стадий опухолей яичников. Рак рез. 2001; 61: 5895–5904. [PubMed] [Google Scholar]
128. Маркес Р.Т. и др. Паттерны экспрессии генов при различных гистотипах эпителиального рака яичников коррелируют с таковыми в нормальных фаллопиевых трубах, эндометрии и толстой кишке. клин. Рак рез. 2005;11:6116. [PubMed] [Академия Google]
Это исследование показало, что профили экспрессии генов различных гистотипов рака яичников коррелируют с их морфологическими аналогами в нормальных тканях.
Маркес Р.Т. и др. Паттерны экспрессии генов при различных гистотипах эпителиального рака яичников коррелируют с таковыми в нормальных фаллопиевых трубах, эндометрии и толстой кишке. клин. Рак рез. 2005;11:6116. [PubMed] [Академия Google]
Это исследование показало, что профили экспрессии генов различных гистотипов рака яичников коррелируют с их морфологическими аналогами в нормальных тканях.
129.
Ченг В. и др. Линейная неверность эпителиального рака яичников контролируется генами HOX , которые определяют региональную идентичность в репродуктивном тракте. Природа Мед. 2005; 11:531. [PubMed] [Академия Google]
Авторы приводят убедительный аргумент, что гены HOX играют роль в определении гистотипов рака яичников.
130. Tothill RW, et al. Новые молекулярные подтипы серозного и эндометриоидного рака яичников связаны с клиническим исходом. клин. Рак рез. 2008;14:5198–5208. [PubMed] [Google Scholar]
131. Schwartz DR, et al. Экспрессия генов при раке яичников отражает как морфологию, так и биологическое поведение, отличая светлоклеточный рак от других карцином яичников с плохим прогнозом. Рак рез. 2002; 63: 4722–4729. [PubMed] [Google Scholar]
Рак рез. 2002; 63: 4722–4729. [PubMed] [Google Scholar]
132.
Курман Р.Дж., Ши Л.Э.М. Патогенез рака яичников: уроки морфологии и биологии и их клиническое значение. Междунар. Дж. Гинекол. Патол. 2008; 27: 151–160. [Бесплатная статья PMC] [PubMed] [Google Scholar]
В этом обзоре обобщены данные о раке яичников I и II типов.
133.
Баст Р.С., младший, и др. Радиоиммуноанализ с использованием моноклональных антител для мониторинга течения эпителиального рака яичников. Новый англ. Дж. Мед. 1983; 309: 883–887. [PubMed] [Академия Google]
Это исходный отчет об анализе CA125.
134. Spaeth EL, et al. Переход мезенхимальных стволовых клеток в ассоциированные с опухолью фибробласты способствует расширению фиброваскулярной сети и прогрессированию опухоли. ПЛОС ОДИН. 2009;4:e4992. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Учебные советы для занятий по биологии — Листы с рекомендациями
Учебные советы для занятий по биологии
Учеба на уроках биологии сильно отличается от подготовки к урокам истории или английского языка. Стратегии, которые хорошо работали в этих классах, могут не сработать здесь. Ниже приведены стратегии обучения, предназначенные для учащихся на уроках биологии. У вас, вероятно, не будет времени попробовать все эти стратегии, но выберите несколько, которые, по вашему мнению, могут помочь, и попробуйте их. Ключ в том, чтобы найти как можно больше различных способов работы с информацией, которую вам дают.
Стратегии, которые хорошо работали в этих классах, могут не сработать здесь. Ниже приведены стратегии обучения, предназначенные для учащихся на уроках биологии. У вас, вероятно, не будет времени попробовать все эти стратегии, но выберите несколько, которые, по вашему мнению, могут помочь, и попробуйте их. Ключ в том, чтобы найти как можно больше различных способов работы с информацией, которую вам дают.
Перед занятием
- Очень важно, чтобы вы прочитали текст перед занятием; но то, КАК вы читаете книгу, имеет значение.
- Если у вас нет времени прочитать всю главу, хотя бы посмотрите картинки (и прочитайте подписи). Биология — это визуальный предмет, и многие понятия лучше всего объясняются в виде картинок.
- Не пытайтесь запомнить всю главу. Многие преподаватели не будут использовать ВСЮ информацию в тексте, а добавят информацию, которой нет в тексте. Преподаватель сообщит вам, какие части главы наиболее актуальны.
- Составьте список всех слов в главе, которые вы не понимаете. Очень вероятно, что это термины, которые учитель будет объяснять на уроке. Если вы уже записали эти слова – у вас будет фора, чтобы делать заметки в классе. Кроме того, вы с большей вероятностью поймете лекцию, если будете иметь представление о том, какие термины будут важны (даже если вы не знаете, что они означают)
- Если картинка используется несколько раз в одной и той же главе — наверное, это важно. Вы можете сделать копию фотографии и принести ее с собой в класс. Или, может быть, найти похожую картинку в Интернете и распечатать ее. Подписать изображение может быть проще, чем пытаться нарисовать его в своих заметках. Однако эта стратегия может оказаться неподходящей – это зависит от класса и преподавателя.
- Если у преподавателя есть онлайн-заметки, контрольные листы или другая информация, прочтите их перед занятием. Также не забудьте взять их с собой на урок.
 Очень вероятно, что это термины, которые учитель будет объяснять на уроке. Если вы уже записали эти слова – у вас будет фора, чтобы делать заметки в классе. Кроме того, вы с большей вероятностью поймете лекцию, если будете иметь представление о том, какие термины будут важны (даже если вы не знаете, что они означают)
Очень вероятно, что это термины, которые учитель будет объяснять на уроке. Если вы уже записали эти слова – у вас будет фора, чтобы делать заметки в классе. Кроме того, вы с большей вероятностью поймете лекцию, если будете иметь представление о том, какие термины будут важны (даже если вы не знаете, что они означают)
Во время занятий
- Приготовьтесь! Принесите свою книгу, а также любые наборы заметок / обзорные листы, которые предоставляет преподаватель. Часто вы можете сэкономить время, если будете делать заметки в книге или в наборах заметок, а не в классе. Одна из наиболее распространенных трудностей на уроках естествознания заключается в том, что трудно не отставать от скорости лекций. Если у вас есть картинки из книги (или ксерокопии), вы сможете делать заметки намного быстрее.
- Если вы делаете заметки в своей книге или где-то еще – обязательно включите это в свои заметки. Напишите что-нибудь вроде «см. рис. 3.5» в своих заметках, чтобы знать, когда смотреть на картинки в книге.
- Не пытайтесь записывать все, что говорит инструктор. Это один из самых сложных аспектов жизни студента – понять, что важно в лекции. Это требует практики. Несколько советов по ведению заметок:
- Если инструктор напишет это на доске — запишите в свои заметки.
- Если инструктор говорит «это важно» или что-то подобное — пишите в заметки.
- Если инструктор ссылается на что-то в книге – запишите номер страницы или рисунка, чтобы вернуться к ним позже.
- Разработайте систему для заметок. Существуют официальные методы ведения заметок – или вы можете разработать свои собственные. Если есть наборы слов, которые часто используются на уроке – придумайте для них символ, который вы запомните. (Вы можете составить список этих символов на случай, если забудете.)
- Вы можете попробовать поработать с системой ведения заметок, посмотрев телепередачу или фильм и делая заметки.
- Если вы что-то пропустили во время лекции:
- Попросите инструктора повторить информацию.
- Оставьте место и продолжайте — вернитесь и заполните то, что вы пропустили позже.
- Не пропустите следующий пункт, потому что вы спрашивали своего соседа о последнем пункте.
- Если что-то непонятно на уроке – СПРАШИВАЙТЕ!
- Большинство преподавателей не будут возражать против того, чтобы вы попросили разъяснений в классе. Часто неловко быть тем, кто задает вопросы, но ваши сокурсники будут рады, что вы это сделали. (Вероятно, у них есть один и тот же вопрос, и они просто постеснялись его задать.)
- Если вы задаете вопросы, вы не выглядите глупым учеником – вы выглядите заинтересованным учеником.
- Большинство преподавателей не будут возражать против того, чтобы вы попросили разъяснений в классе. Часто неловко быть тем, кто задает вопросы, но ваши сокурсники будут рады, что вы это сделали.
- Если у вас есть время, вы можете записать лекции. Таким образом, если вы что-то пропустили, вы всегда можете вернуться к этому. Вы также можете прослушать всю лекцию еще раз. Некоторые учащиеся слушают их по дороге на работу – это делает время, проведенное в пробке, более продуктивным. Или вы можете слушать их во время уборки дома или во время пробежки. Ключом к этой стратегии является повторение – оно может не сработать с первого раза, но в конечном итоге оно сработает.
 Часто вы можете сэкономить время, если будете делать заметки в книге или в наборах заметок, а не в классе. Одна из наиболее распространенных трудностей на уроках естествознания заключается в том, что трудно не отставать от скорости лекций. Если у вас есть картинки из книги (или ксерокопии), вы сможете делать заметки намного быстрее.
Часто вы можете сэкономить время, если будете делать заметки в книге или в наборах заметок, а не в классе. Одна из наиболее распространенных трудностей на уроках естествознания заключается в том, что трудно не отставать от скорости лекций. Если у вас есть картинки из книги (или ксерокопии), вы сможете делать заметки намного быстрее.
 (Вероятно, у них есть один и тот же вопрос, и они просто постеснялись его задать.)
(Вероятно, у них есть один и тот же вопрос, и они просто постеснялись его задать.)
В лабораторном классе
- Прочтите лабораторную работу за этот день и убедитесь, что вы полностью понимаете, что вы делаете в классе. На большинстве лабораторных занятий от вас ожидают самостоятельности при выполнении заданий, поэтому вам нужно понимать, что от вас ожидают в этот день. В качестве дополнительного бонуса, если вы организованы и у вас есть план – вы даже можете закончить раньше. С другой стороны, если вы потратите лабораторное время на выяснение того, что вы должны делать, вы можете не закончить лабораторную работу.
- Разделяй и властвуй. У вас может не хватить времени, чтобы выполнить все части эксперимента самостоятельно. Частью навыков, которым должны научить лабораторные занятия, является совместная работа. Однако убедитесь, что вы понимаете все части эксперимента, даже если на самом деле вы его не проводили.
 В качестве дополнительного бонуса, если вы организованы и у вас есть план – вы даже можете закончить раньше. С другой стороны, если вы потратите лабораторное время на выяснение того, что вы должны делать, вы можете не закончить лабораторную работу.
В качестве дополнительного бонуса, если вы организованы и у вас есть план – вы даже можете закончить раньше. С другой стороны, если вы потратите лабораторное время на выяснение того, что вы должны делать, вы можете не закончить лабораторную работу.
После занятий
- Перепишите свои записи. Это отнимает много времени, но делает две вещи. Это дает вам возможность просмотреть то, что вы прошли в классе, и убедиться, что вы ничего не пропустили. Это также дает вам хорошо организованный набор заметок для подготовки к тесту.
- Эта стратегия работает лучше всего, если записи повторяются вскоре после урока.
- Используйте предоставленные ресурсы! Факультет биологии приложил немало усилий, чтобы предоставить своим студентам учебную комнату (LS 126). Пожалуйста, используйте это. В этой комнате вам могут быть доступны модели, практические тесты или компьютерные ресурсы. Есть также репетиторы, которые могут помочь ответить на вопросы.
- Найдите онлайн-ресурсы, содержащие ту же информацию. Попробуйте ввести в поисковик тему и слово «учебник» или «викторина». Я рекомендую использовать google.com — это очень всеобъемлющая поисковая система. В Google также есть библиотека изображений, в которой вы можете искать изображения бактерий, мышц, цветов или всего, что вы ищете.
- Если у вас есть вопросы, задайте их инструктору в рабочее время. Преподаватели НИКОГДА не возражают против того, что вы задаете им вопросы (относящиеся к классу).
 Пожалуйста, используйте это. В этой комнате вам могут быть доступны модели, практические тесты или компьютерные ресурсы. Есть также репетиторы, которые могут помочь ответить на вопросы.
Пожалуйста, используйте это. В этой комнате вам могут быть доступны модели, практические тесты или компьютерные ресурсы. Есть также репетиторы, которые могут помочь ответить на вопросы.
Подготовка к тесту
- Напишите свой собственный тест. Если бы у вас было 20 (или 50, или 100) вопросов, которые вы могли бы задать об этой информации – что бы вы спросили? Какие темы самые важные? Как бы вы задали вопросы по каждой из этих тем? Знать, что будет на тесте, — сложный навык, но с практикой вы сможете понять это.
- Найдите учебную группу. Этот метод не работает для всех, но он может быть очень эффективным. Обсудите тему и убедитесь, что все ее понимают, или задайте друг другу вопросы. Это отличный ресурс, чтобы убедиться, что вы правильно понимаете концепции.

Стратегии для тестов на запоминание
Приведенные ниже стратегии особенно полезны для классов или тем, которые имеют много словарного запаса или названий.
- Карточки, старые любимые. Ключ к карточкам — писать ОЧЕНЬ МАЛЕНЬКО на карточке. Некоторые ученики даже разрезают карточки пополам, чтобы не писать на них слишком много.
- Несколько советов, как сделать хорошие карточки:
- На одной стороне карточки должно быть одно словарное слово, а на другой стороне должно быть определение или изображение
- В качестве альтернативы вы можете написать один вопрос на лицевой стороне и ответ на обратной стороне.
- Несколько советов по использованию карточек:
- Обязательно изучайте карточки в обоих направлениях (смотрите на слово и произносите определение, смотрите на определение и произносите слово).
- Держите их при себе. Изучайте их, пока стоите в очереди или пока выключается микроволновка. Ключевым моментом здесь является то, что немного чаще учиться лучше, чем много учиться в течение короткого времени.
- Сложите стопки из карточек. Куча информации, которую вы знаете, и куча информации, которую вы забыли. Затем возьмите вторую стопку и снова пройдите ее – и снова сложите карты в две стопки. Так вы потратите больше времени на изучение карт, с которыми у вас возникают трудности.
- Обязательно изучайте карточки в обоих направлениях (смотрите на слово и произносите определение, смотрите на определение и произносите слово).
- Несколько советов, как сделать хорошие карточки:
- Скопируйте картинки из книги (и удалите этикетки). Сделайте примерно пять копий каждой картинки. Развесьте эти картинки по всему дому (с этим придется столкнуться вашей семье!). Проходя мимо каждой картинки, подпишите что-нибудь на ней. В следующий раз, когда будете проходить мимо, назовите что-нибудь другое. Вскоре вы пометите большинство предметов на картинке. Когда у вас останется одна или две – если вы их знаете, назовите их – если вы их не знаете, изучите их.
- Возьмите книжку-раскраску. На рынке есть несколько очень хороших раскрасок. У них есть книжки-раскраски для большинства тем, которые вы будете изучать. Это отличный ресурс и хороший способ изучить определенный материал. Опять же, ключ понемногу. Раскрасьте одну страницу за обедом, одну во время просмотра телевизора после ужина и т. д.
- Если вы не можете позволить себе книжку-раскраску (они могут быть немного дорогими), скопируйте или обведите картинки из учебника, а затем раскрасьте и подпишите их. (Сделайте несколько копий этих изображений и удалите этикетки.)
- Ищите онлайн-викторины.
- Обязательно используйте время открытой лаборатории, если оно предусмотрено. Картинки красивые, но не такие, как настоящие. Используйте эти стратегии изучения, когда вы держите и смотрите на трехмерные модели/образцы:
- Просмотрите список терминов и найдите их все в объекте.
- Не глядя в список, укажите и назовите все части объекта, которые вам нужно знать. Затем проверьте список.
- Не глядя в список, укажите и запишите названия всех частей предмета, которые вам нужно знать. Затем проверьте список. Помните, что на тесте вам придется писать (и произносить слово по буквам), а не произносить его вслух.
- Попросите кого-нибудь указать на деталь, и вы напишите ее на листе бумаги. Это лучший способ обучения, потому что именно так устроено большинство практических занятий в лаборатории. Если вы не можете найти другого студента, чтобы проверить вас, спросите репетиторов или вашего инструктора.


 Затем проверьте список.
Затем проверьте список.
Стратегии изучения концептуальных тестов
Эти стратегии лучше всего подходят для занятий и тем, посвященных процессам.
- Ниже приводится метод, который поможет вам изучить сложный процесс (например, если вам нужно изучить все этапы аэробного дыхания). Когда вы освоитесь с одним из этих шагов, переходите к следующему уровню сложности:
- Нарисуйте, обведите или скопируйте изображение процесса из своей книги (удалите этикетку). Напишите словарные слова, относящиеся к процессу, на другом листе бумаги. Уметь поставить правильный термин в нужном месте на картинке.
- Теперь попробуйте подписать картинку без терминов перед вами.
- Теперь посмотрите на список словарных слов. Расставь слова по порядку и нарисуй картинку. Обозначьте цветом различные этапы процесса.
- Теперь возьмите чистый лист бумаги и нарисуйте картинку с нуля, без слов перед собой. Подпишите нарисованную вами картинку.
- Теперь возьмите чистый лист бумаги и нарисуйте процесс в обратном порядке. (Эй, почему бы и нет!)
- Нарисуйте, обведите или скопируйте изображение процесса из своей книги (удалите этикетку).
- Выполните вышеуказанный процесс, но вместо того, чтобы рисовать картинку, напишите абзац, объясняющий процесс.
- Вы также можете взять все части процесса и записать их на карточках для заметок. Теперь используйте карточки для заметок, чтобы объяснить процесс.
- Или можно купить пластилин и построить модель процесса. (На самом деле не имеет значения, как вы это сделаете, но вам нужно взять процесс и разбить его на более мелкие части, которые вы сможете понять и собрать вместе.
 Напишите словарные слова, относящиеся к процессу, на другом листе бумаги. Уметь поставить правильный термин в нужном месте на картинке.
Напишите словарные слова, относящиеся к процессу, на другом листе бумаги. Уметь поставить правильный термин в нужном месте на картинке.
