Широколиственные леса
TR | UK | KK | BE | EN |широколиственные леса, широколиственные леса россии картинки
Широ́коли́ственные леса́ — разновидность лиственных лесов, образованных листопадными (летнезелёными) деревьями с широкими листовыми пластинками.
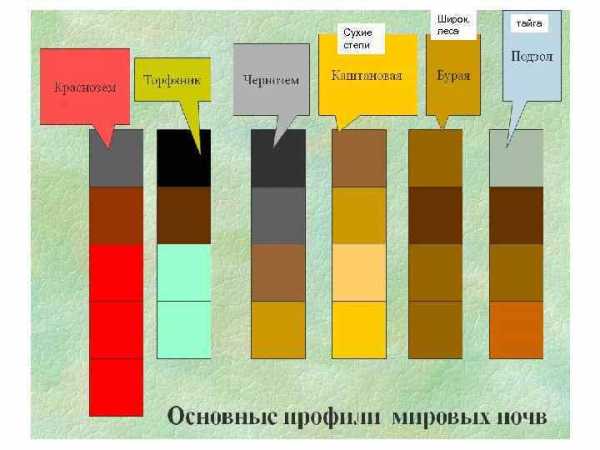
Широколиственные леса приурочены к влажным и умеренно-влажным районам умеренного климата с ослабленной континентальностью, равномерным распределением осадков в течение года и относительно высокими температурами. Почвы широколиственных лесов — обычно серые, подзолистые, тёмно-серые и бурые лесные, реже чернозёмы. Распространены в Европе, Северной Америке, Восточной Азии, на юге Чили и островах Новая Зеландия.
Содержание
- 1 Климат
- 2 Растительный мир
- 3 Животные
- 3.1 Млекопитающие
- 3.2 Птицы
- 3.3 Рептилии и амфибии
- 3.4 Рыбы
- 4 Ссылки
Климат
Умеренно теплый климат характеризуется теплым, продолжительным летом, мягкой зимой. Годовая сумма осадков, равномерно распределенных в течение года, несколько выше испарения, что существенно снижает уровень заболоченности грунтов.
Растительный мир
Древесные породы в Европе представлены главным образом буком, дубом, реже грабом, липой. Обычны также ясень, вяз, клён. В Северной Америке наиболее широко распространены кленово-буковые, дубово-гикоровые, дубовые, а в прошлом — дубово-каштановые леса. Для подлеска Восточной Европы обычны лещина, полевой и татарский клёны, черёмуха, бересклеты. В травяном покрове господствуют мезофильные эвтрофные или мезотрофные виды, в европейских лесах — пролесник, сныть, зеленчук, копытень, медуница, ясменник, осока волосистая и другие.
Широколиственным лесам свойственны весенние эфемероиды-геофиты: хохлатка, ветреница, зубянка, подснежник, пролеска, гусиный лук, которые успевают пройти цикл развития от распускания листьев до созревания семян весной, до развёртывания листьев деревьев.
Животные
Часть Голарктической зоны.
Млекопитающие
Млекопитающие широколиственных лесов Западной и Восточной Европы: дикий кабан, олени (благородный олень; в лесной зоне вне Европы встречаются такие его подвиды, как марал, вапити, изюбрь), лось, косуля, зубр, лесная белка, бурундук, бобр, рыжая полёвка, желтогорлая мышь, сони (полчок, лесная соня, садовая и орешниковая), заяц-беляк, заяц-русак, обыкновенный ёж, лесная кошка, рысь, лисица, волк, бурый медведь, барсук, горностай, лесная куница, ласка, норка европейская. Крупные животные сохранились лишь в отдельных массивах, не уничтоженных хозяйственной деятельностью человека (например, в Карпатских горах).
Фауна муссонных широколиственных лесов на Дальнем Востоке (амурско-уссурийских, китайских, корейских) особенно богата, так как туда проникают виды с юга. Из млекопитающих характерны уссурийский крот, маньчжурский заяц, пятнистый олень, горал, уссурийский черный медведь (гималайский, или белогрудый медведь), харза, енотовидная собака, амурский тигр, дальневосточный леопард, дальневосточный лесной кот.
Необычное животное широколиственных лесов Америки — скунс. По чёрной шубке скунса от головы к большому пушистому хвосту проходят две широкие белые полосы. Другой лесной обитатель, свойственный Америке, — енот, или енот-полоскун.
Птицы
Ястреб-тетеревятник, иволга, поползень, чиж, рябчик, тетерев-косач, глухарь, дрозд-рябинник, соловей, синица, снегирь, жаворонки, серый журавль, зяблик, кукушка, дубонос, большой пёстрый, зелёный и средний дятлы, осоед, канюк, совы, филины, голуби,аисты, вороны, галки, сороки, грачи, гуси, утки и др.; в Азии: личинкоед, белоглазка, голубая сорока, широкорот, фазан, утка-мандаринка.
Рептилии и амфибии
Рептилии: уж, гадюка, эскулапов полоз, безобидная змея медянка (неправильно считающаяся ядовитой), безногая ящерица веретеница, зелёная и живородящая ящерицы, болотная черепаха. Если рассматривать территорию России, то нельзя не упомянуть кожистую черепаху (уссурийская, или китайская черепаха).
Земноводные: квакша, прудовая, травяная и остромордая лягушки, зелёная жаба, жерлянки, гребенчатый тритон, пятнистая саламандра.
Рыбы
Сом, щука, пескарь, карповые, лососёвые и др.
Ссылки
- Широколиственные леса — статья из Большой советской энциклопедии (Проверено 21 мая 2010)
| Природные зоны | |
|---|---|
| Суша | Арктическая пустыня | Антарктическая пустыня | Тундра | Лесотундра | Хвойный лес — Тайга | Подтайга | Смешанный лес | Лиственный лес: широколиственные леса и мелколиственные леса | Лесостепь | Степь | Полупустыня | Пустыня | Саванна | Субтропический лес | Дождевой лес | Тропический лес | Переменно-влажный тропический лес | Джунгли | Сухие тропические леса: Каатинга и Мата | Влажные тропические леса: Мангры, Сельва | Альпийская | Субальпийская | Нивальная | Средиземноморская | Высотная поясность | Прибрежные и заболоченные: Плавни, Пойма, Болота, Солончаки, Марши, Болотные и заболоченные леса умеренного пояса | Пруд | Пещеры |
| Мировой океан | Супралиторальная зона | Литоральная зона: Мангры | | Сублиторальная зона | Неритическая зона | Зона открытого океана | Батиальная зона | Псевдобатиальная зона | Псевдоабиссальная зона | Талассобатиальная зона | Абиссальная зона | Ультраабиссальная зона | Коралловые рифы | Континентальный шельф | Пелагическая зона | Ложе океана | Впадины: жёлоб и рифт | Гидротермальные источники срединно-океанических хребтов | Паковый лёд | Леса бурых водорослей | Устье: дельта и эстуарий |
| Биология — География — Экология | |
широколиственные леса, широколиственные леса в россии, широколиственные леса животные, широколиственные леса климат, широколиственные леса растения, широколиственные леса россии, широколиственные леса россии картинки
Широколиственные леса Информацию О

Широколиственные леса Комментарии
Широколиственные леса
Широколиственные леса
Широколиственные леса Вы просматриваете субъект
Широколиственные леса что, Широколиственные леса кто, Широколиственные леса описание
There are excerpts from wikipedia on this article and video
www.turkaramamotoru.com
Планета Земля » Широколиственные леса
Европейские широколиственные леса — исчезающие лесные экосистемы. Всего несколько веков назад они занимали большую часть Европы и были одними из самых богатых и разнообразных на планете. В XVI — XVII вв. естественные дубовые леса произрастали на площади в несколько миллионов гектаров, а сегодня, по данным учета лесного фонда, их осталось не более 100 тысяч га. Так за несколько столетий площадь этих лесов сократилась в десятки раз. Образованные листопадными деревьями с широкими листовыми пластинками, широколиственные леса распространены в восточной части Северной Америки, Европе, Северном Китае, Японии и на Дальнем Востоке. Они занимают площадь между смешанными лесами на севере и степями, средиземноморской или субтропической растительностью на юге.
Широколиственные леса произрастают в районах с влажным и умеренно влажным климатом, для которых характерно равномерное распределение осадков (от 400 до 600 мм) в течение года и относительно высокие температуры. Средняя температура января -8…0 °C, а июля +20…+24 °С. Умеренно тёплые и влажные климатические условия, также активная деятельность почвенных организмов (бактерий, грибов, беспозвоночных) способствуют быстрому разложению листьев и накоплению гумуса. Под широколиственными лесами формируются плодородные серые лесные и бурые лесные почвы, реже чернозёмы.
Верхний ярус в этих лесах занимают дуб, бук, граб и липа. В Европе встречаются ясень, ильм, клён, вяз. Подлесок образован кустарниками — лещиной, бересклетом бородавчатым, жимолостью лесной. В густом и высоком травяном покрове европейских широколиственных лесов господствуют сныть, зеленчук, копытень, медуница, ясменник, осока волосистая, весенние эфемероиды: хохлатки, ветреница, подснежник, пролеска, гусиный лук и др. В Северной Америке в этой зоне произрастают виды дубов, характерные только для этого материка. В широколиственных лесах Южного полушария преобладает южный бук.
Современные широколиственные и хвойно-широколиственные леса сформировались пять-семь тысячелетий назад, когда на планете наступило потепление и широколиственные породы деревьев смогли далеко продвинуться на север. В последующие тысячелетия климат становился более холодным и зона широколиственных лесов постепенно уменьшалась. Поскольку под этими лесами сформировались наиболее плодородные из всей лесной зоны почвы, леса интенсивно вырубались, а их место занимала пашня. Кроме того, дуб, обладающий очень прочной древесиной, широко использовался в строительстве.
Правление Петра I стало для России временем создания парусного флота. «Задумка царская» требовала большого количества высококачественной древесины, поэтому так называемые корабельные рощи строго охранялись. Леса, не входившие в охраняемые территории, жители лесной и лесостепной зоны активно вырубали под пашни и луга. В середине XIX в. закончилась эпоха парусного флота, корабельные рощи перестали охранять, и леса начали сводить ещё интенсивнее.
К началу XX в. сохранились лишь фрагменты некогда единого и обширного пояса широколиственных лесов. Уже тогда пытались выращивать новые дубы, но это оказалось делом непростым: молодые дубовые рощицы гибли из-за частых и сильных засух. Исследования, проведённые под руководством великого русского географа В.В. Докучаева, показали, что эти бедствия были связаны с широкомасштабным сведением лесов и, как следствие, изменениями гидрологического режима и климата территории.
Тем не менее и в XX столетии оставшиеся дубравы интенсивно вырубались. Насекомые-вредители и холодные зимы конца века привели к тому, что вымирание естественных дубовых лесов стало неизбежным.
Сегодня на некоторых участках, где раньше произрастали широколиственные леса, распространились вторичные леса и искусственные насаждения, в которых преобладают хвойные породы деревьев. Восстановить структуру и динамику естественно-природных дубовых лесов не только России, но и всей Европе (где они испытали еще более сильное антропогенное влияние) уже вряд ли удастся.
zemlj.ru
3. Зона широколиственных лесов.
В структуре и функции геосистем широколиственных лесов следует отметить некоторые специфические черты: разорванность ареала на Евроазиатском континенте, приуроченность к зонам с умеренно континентальным климатом, высокая требовательность растительности к почвенному плодородию, значительно большое флористическое богатство и разнообразие животного населения по сравнению с хвойными лесами, присутствие в травяном покрове растений с широкой листовой пластинкой (“спутники дубрав”), слабое семенное воспроизводство травянистых растений в связи с сильным летним затенением, развитие мощной лесной подстилки и слабым промерзанием почвы, большая роль гетеротроф-комплекса в функции геосистем (в естественном возобновлении деревьев и кустарников и их расселении, в повреждении различных органов растений, во влияние на разложение и гумификацию лесной подстилки, в изменении состава, влажности и аэрированности почвы).
В силу исторических, геологических и экологических причин широколиственные леса подвергались значительно большему воздействию со стороны человека, чем хвойные. Наиболее распространенные формы антропогенного воздействия – вырубка леса, пожары, выпас, и как следствие этого, сведение широколиственных лесов на больших площадях, смена коренных лесов производными. Существенное изменение структуры и видового состава, изменение гидрологического режима, усиление эрозионной и селевой деятельности.
4. Зона лесостепи.
Особенности лесостепи – комплексность растительного покрова, в котором в качестве зональных элементов выступают такие контрастные геосистемы, как лес и степь, близкие к единице соотношения тепла и влаги, неустойчивость увлажнения, проявляющаяся в наличии частых бездождевых летних периодов и засушливых лет, разорванность ареала, что приводит к необходимости выделения европейской и азиатской лесостепи, отличающихся друг от друга количеством тепла и влаги, рельефом, почвами и растительностью, сильное антропогенное изменение природы, отсутствие ненарушенных территорий.
5. Зона степи.
Много общего с лесостепью. Из особенностей степной зоны можно отметить преобладание на ее территории травянистых растений (в основном сельскохозяйственных посевов), засушливость и неустойчивость увлажнения, выраженные еще более резко, чем в лесостепи, сильное антропогенное изменение природы (распаханность степей более 50%, в отдельных районах 80-90%).
Большие площади посевов в степной зоне подвергаются искусственному орошению. При этом возникает потенциальная опасность засоления почв и других изменений на больших территориях. За этими процессами необходим постоянный контроль.
6. Зона пустынь.
Геосистемы пустынь, особенно песчаных, относятся к наиболее неустойчивым и легкоранимым при антропогенном воздействии. Пустыни Средней Азии и Казахстана весьма разнообразны, причем каждый из типов пустынь требует своей организации, объема и методики слежения за состоянием и динамикой геосистем.
Важным объектом ГСМ в песчаных пустынях становятся зоны влияния крупных оросительных каналов – под их влиянием в результате фильтрации (каналы проложены в естественном грунте без гидроизоляции) произошли и продолжают происходить коренные изменения геосистем. вдоль каналов в полосе 5 – 30 км шириной развиваются комплексы гидроморфного типа, отличающиеся растительностью, соответствующим животным населением, особыми гидрогеологическими условиями.
Литература:
1. Пономаренко О.И., Ботвинкина М.А. Методы контроля природных объктов и мо-ниторинг окружающей среды: учебно-методическое пособие. – Алматы, 2011.
2. Горшков М.В. Экологический мониторинг. Учебное пособие. – Владивосток: Изд-во ТГЭУ, 2010.
studfiles.net
ШИРОКОЛИСТВЕННЫЕ ЛЕСА зона неморальных лесов подтаежные леса
 ШИРОКОЛИСТВЕННЫЕ ЛЕСА (зона неморальных лесов)
ШИРОКОЛИСТВЕННЫЕ ЛЕСА (зона неморальных лесов)
 подтаежные леса По видовому составу аппалачские леса – одни из богатейших на Земле. Больше всего в них американских видов дубов, вместе с ними распространены каштан, бук, ясень, липа, платан. Преобладают высокие деревья с мощной раскидистой кроной, часто обвитые вьющимися растениями – виноградом или плющом. В южной части зоны встречаются такие древние теплолюбивые виды, как гикори, магнолия, тюльпанное дерево и ликвидамбар.
подтаежные леса По видовому составу аппалачские леса – одни из богатейших на Земле. Больше всего в них американских видов дубов, вместе с ними распространены каштан, бук, ясень, липа, платан. Преобладают высокие деревья с мощной раскидистой кроной, часто обвитые вьющимися растениями – виноградом или плющом. В южной части зоны встречаются такие древние теплолюбивые виды, как гикори, магнолия, тюльпанное дерево и ликвидамбар.
 Широкий пояс хвойно-широколиственных насаждений образует переходную зону между бореальными (тайга) и неморальными (чисто лиственными) лесами в Европе и на Дальнем Востоке
Широкий пояс хвойно-широколиственных насаждений образует переходную зону между бореальными (тайга) и неморальными (чисто лиственными) лесами в Европе и на Дальнем Востоке
 По сравнению с тайгой широколиственные леса занимают вдвое меньшую площадь и почти вдвое превосходят ее по удельной продуктивности. Поэтому суммарная годовая продуктивность в этих природных зонах примерно одинакова
По сравнению с тайгой широколиственные леса занимают вдвое меньшую площадь и почти вдвое превосходят ее по удельной продуктивности. Поэтому суммарная годовая продуктивность в этих природных зонах примерно одинакова
 Сухие степи Широк. леса тайга
Сухие степи Широк. леса тайга
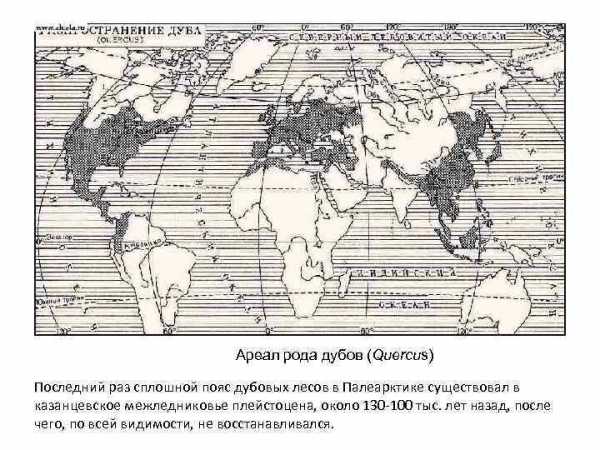 Ареал рода дубов (Quercus) Последний раз сплошной пояс дубовых лесов в Палеарктике существовал в казанцевское межледниковье плейстоцена, около 130 -100 тыс. лет назад, после чего, по всей видимости, не восстанавливался.
Ареал рода дубов (Quercus) Последний раз сплошной пояс дубовых лесов в Палеарктике существовал в казанцевское межледниковье плейстоцена, около 130 -100 тыс. лет назад, после чего, по всей видимости, не восстанавливался.
 В Северной Америке произрастает около 40 видов дубов, в Европе, Малой Азии и Северной Африке – около 20 видов
В Северной Америке произрастает около 40 видов дубов, в Европе, Малой Азии и Северной Африке – около 20 видов
 Южной границей бореальной зоны в Европе можно считать северную границу распространения дуба черешчатого.
Южной границей бореальной зоны в Европе можно считать северную границу распространения дуба черешчатого.


 Орех грецкий Орех манчьжурский
Орех грецкий Орех манчьжурский
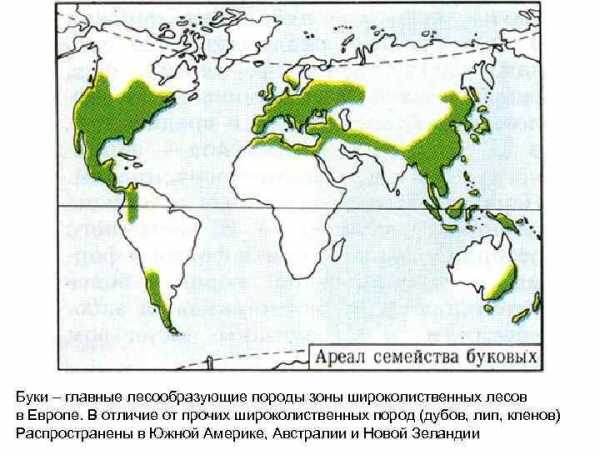 Буки – главные лесообразующие породы зоны широколиственных лесов в Европе. В отличие от прочих широколиственных пород (дубов, лип, кленов) Распространены в Южной Америке, Австралии и Новой Зеландии
Буки – главные лесообразующие породы зоны широколиственных лесов в Европе. В отличие от прочих широколиственных пород (дубов, лип, кленов) Распространены в Южной Америке, Австралии и Новой Зеландии
 Вечнозеленые буковые леса восточной Австралии, Новой Зеландии и на западных склонах Анд относятся к экосистемам влажных лесов умеренного и субтропического пояса
Вечнозеленые буковые леса восточной Австралии, Новой Зеландии и на западных склонах Анд относятся к экосистемам влажных лесов умеренного и субтропического пояса

 В Северной Америке пограничная полоса между широколиственными лесами и степями (прериями) сформирована саванноподобными ландшафтами, где доминируют разные виды дубов (дубовые саванны)
В Северной Америке пограничная полоса между широколиственными лесами и степями (прериями) сформирована саванноподобными ландшафтами, где доминируют разные виды дубов (дубовые саванны)
 Спелый дубовый лес
Спелый дубовый лес

 Старейший дуб – главная достопримечательность Шервудского леса в Нотингеме, Англии. Согласно легенде это величественное дерево служило местом ночлега для героя английского фольклора – Робин Гуда и его шайки. Возраст дерева насчитывает около 800 − 1000 лет.
Старейший дуб – главная достопримечательность Шервудского леса в Нотингеме, Англии. Согласно легенде это величественное дерево служило местом ночлега для героя английского фольклора – Робин Гуда и его шайки. Возраст дерева насчитывает около 800 − 1000 лет.
 Дубняк из дуба монгольского в Южном Приморье
Дубняк из дуба монгольского в Южном Приморье
 Дубняк из дуба зубчатого в Лазовском заповеднике
Дубняк из дуба зубчатого в Лазовском заповеднике
 Липовый лес
Липовый лес
 Буковый лес
Буковый лес

 Буковый лес
Буковый лес



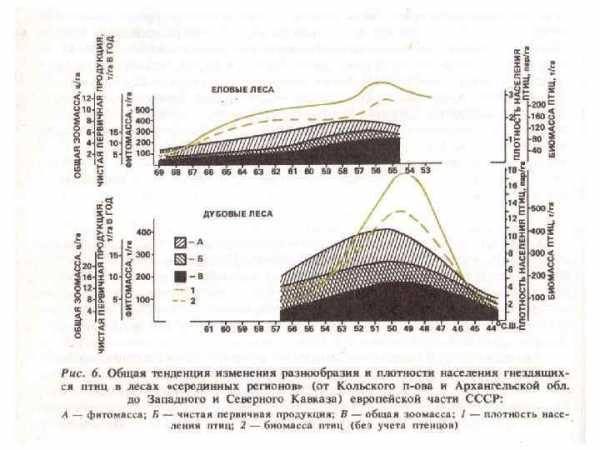
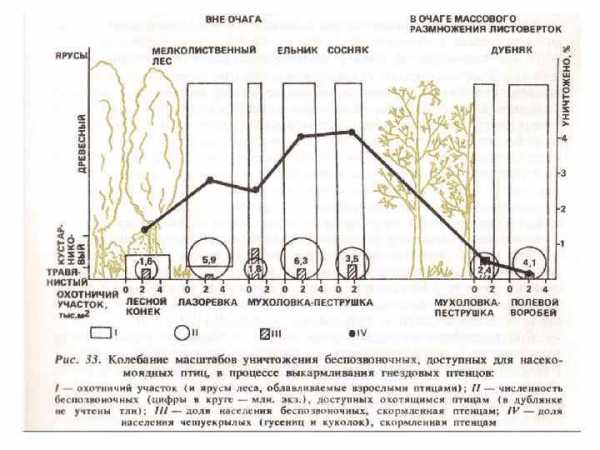
 1. Массовые листогрызущие насекомые ведут преимущественно открытый образ жизни. 2. Во взрослой фазе не питаются или используют воду и нектар цветков. Процессы жизнедеятельности у взрослых насекомых протекают за счет резервных питательных веществ, накопленных личинкой. 3. Насекомым данной группы свойственна высокая потенциальная плодовитость. Кучная откладка яиц ведет к концентрации личинок и образованию очагов. 4. Дальность и быстрота полета у листогрызущих насекомых значительно выше, чем у других групп вредителей. Большинство гусениц способны к активным переползаниям. Летными способностями взрослых особей и миграцией гусениц обусловливается расширение очагов вспышек. 5. Массовые листогрызущие насекомые характеризуются изменчивостью организма, которая проявляется у одного и того же вида в различной величине особей, их массе, окраске, плодовитости. 6. Численность насекомых в насаждениях по годам непостоянна и колеблется в очень широких пределах. Приведенные биологические особенности хвое-листогрызущих насекомых обусловливают резкие колебания их численности и возникновение вспышек массового размножения. 7. Фоновый уровень повреждения листвы в лесах умеренного пояса составляет около 7 %.
1. Массовые листогрызущие насекомые ведут преимущественно открытый образ жизни. 2. Во взрослой фазе не питаются или используют воду и нектар цветков. Процессы жизнедеятельности у взрослых насекомых протекают за счет резервных питательных веществ, накопленных личинкой. 3. Насекомым данной группы свойственна высокая потенциальная плодовитость. Кучная откладка яиц ведет к концентрации личинок и образованию очагов. 4. Дальность и быстрота полета у листогрызущих насекомых значительно выше, чем у других групп вредителей. Большинство гусениц способны к активным переползаниям. Летными способностями взрослых особей и миграцией гусениц обусловливается расширение очагов вспышек. 5. Массовые листогрызущие насекомые характеризуются изменчивостью организма, которая проявляется у одного и того же вида в различной величине особей, их массе, окраске, плодовитости. 6. Численность насекомых в насаждениях по годам непостоянна и колеблется в очень широких пределах. Приведенные биологические особенности хвое-листогрызущих насекомых обусловливают резкие колебания их численности и возникновение вспышек массового размножения. 7. Фоновый уровень повреждения листвы в лесах умеренного пояса составляет около 7 %.
 Дубрава, объеденная дубовым шелкопрядом.
Дубрава, объеденная дубовым шелкопрядом.
 Дубрава, объеденная дубовой листоверткой.
Дубрава, объеденная дубовой листоверткой.


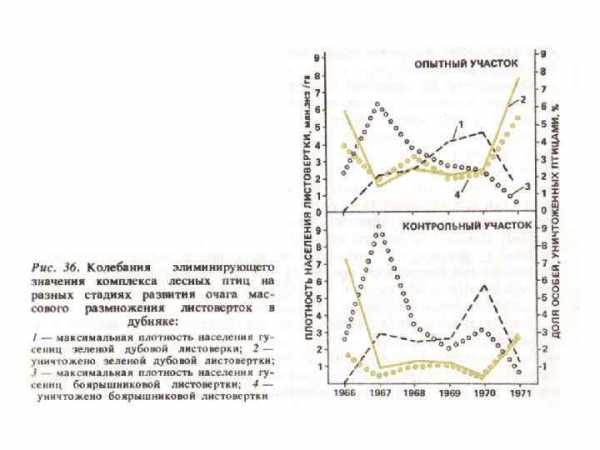
 Зоогенная дефолиация и последующее изменение режима и интенсивности биоценотических процессов приводят к изменению состава, ритма развития, продуктивности травяного покрова. Увеличивается интенсивность солнечной радиации на поверхности почвы, повышается температура воздуха и почвы на несколько градусов (Злотин, 1970 а, б; Домников, 1979 а, б). Поступление в подстилку богатых азотом экскрементов листогрызущих насекомых влияет на газовый режим припочвенного слоя воздуха. Экскременты — центры массового развития микроорганизмов, поэтому интенсивность выделения углекислого газа с поверхности подстилки увеличивается в 2 -3 раза. У деревьев с объеденной кроной резко уменьшается транспирация, расход воды из корнеобитаемого слоя замедляется, что ослабляет конкуренцию трав и деревьев за влагу. В годы массового размножения насекомых усиленно развиваются лесные и опушечные травы. Так, по данным Р. И. Злотина и К. С. Ходашевой (1974), в Курском заповеднике на участках, где листовертка создает прорывы в пологе леса, Aegopodium podagraria и Clematis recta цветут обильнее, чем на неповрежденных участках, у них больше высота, средняя масса, суммарная листовая поверхность. Видовое разнообразие трав при повреждениях деревьев насекомыми возрастает в основном за счет светолюбивых и нитрофильных видов.
Зоогенная дефолиация и последующее изменение режима и интенсивности биоценотических процессов приводят к изменению состава, ритма развития, продуктивности травяного покрова. Увеличивается интенсивность солнечной радиации на поверхности почвы, повышается температура воздуха и почвы на несколько градусов (Злотин, 1970 а, б; Домников, 1979 а, б). Поступление в подстилку богатых азотом экскрементов листогрызущих насекомых влияет на газовый режим припочвенного слоя воздуха. Экскременты — центры массового развития микроорганизмов, поэтому интенсивность выделения углекислого газа с поверхности подстилки увеличивается в 2 -3 раза. У деревьев с объеденной кроной резко уменьшается транспирация, расход воды из корнеобитаемого слоя замедляется, что ослабляет конкуренцию трав и деревьев за влагу. В годы массового размножения насекомых усиленно развиваются лесные и опушечные травы. Так, по данным Р. И. Злотина и К. С. Ходашевой (1974), в Курском заповеднике на участках, где листовертка создает прорывы в пологе леса, Aegopodium podagraria и Clematis recta цветут обильнее, чем на неповрежденных участках, у них больше высота, средняя масса, суммарная листовая поверхность. Видовое разнообразие трав при повреждениях деревьев насекомыми возрастает в основном за счет светолюбивых и нитрофильных видов.
 Шуттер (Schutter, 1956) изучал влияние рыжих лесных муравьев и птиц на численность дубовой листовертки в Нижней Франконии (ФРГ). Он взял 3 опытных участка: 1) с муравейниками и с гнездами птиц (0, 3 га), 2) с гнездами птиц, но без муравейников (1 га), 3) без муравейников и без птичьих гнезд (1 га). После обследования сохранности листвы оказалось, что на первом участке остались неповрежденными 53% листьев, на втором — 32% и на третьем —только 14%. Было также подсчитано количество ниц, отложенных самками дубовой листовертки, и оказалось, что на первом участке было на 38% меньше, а на втором на 12% больше яиц, чем на третьем. Рыжие лесные муравьи уничтожают дубовую листовертку на всех стадиях развития (Gosswald, 1953 d; Gosswald, Kloft, 1956 a), однако наибольший эффект действия муравьев наблюдается в ГДР с мая по июнь (Otto, 1959 а; Rust, 1958), так как молодые личинки от них обычно ускользают. Гессвальд (Gosswald, 1962 b) подсчитал, что в очаге листовертки рабочие одного гнезда уничтожают за день в среднем 65 — 75 тыс. гусениц, около 28 тыс. куколок и около 26 тыс. бабочек. Во время вспышки массового размножения листовертки муравьи чаще посещают заселенные ею деревья и перестраивают систему дорожек. Форма охотпичьей территории при этом меняется (Gosswald, 1962 b; Gosswald, Kloft, 1956 a). Б. С. Щербаков (1953) пишет, что во время массового размножения дубовой листовертки в 1938 г. в Московской области муравьи одного среднего гнезда уничтожали в сутки около 7200 гусениц, а позднее, после окукливания — 6500 куколок вредителя. Истребление последних длилось около 20 дней, и за это время было уничтожопо более 10 тыс. куколок. Когда начался вылет бабочек, муравьи переключились на питание ими, причем ловили их, главным образом, в момент выхода из куколок. За 30 дней лёта было уничтожено более 150 тыс. взрослых бабочек.
Шуттер (Schutter, 1956) изучал влияние рыжих лесных муравьев и птиц на численность дубовой листовертки в Нижней Франконии (ФРГ). Он взял 3 опытных участка: 1) с муравейниками и с гнездами птиц (0, 3 га), 2) с гнездами птиц, но без муравейников (1 га), 3) без муравейников и без птичьих гнезд (1 га). После обследования сохранности листвы оказалось, что на первом участке остались неповрежденными 53% листьев, на втором — 32% и на третьем —только 14%. Было также подсчитано количество ниц, отложенных самками дубовой листовертки, и оказалось, что на первом участке было на 38% меньше, а на втором на 12% больше яиц, чем на третьем. Рыжие лесные муравьи уничтожают дубовую листовертку на всех стадиях развития (Gosswald, 1953 d; Gosswald, Kloft, 1956 a), однако наибольший эффект действия муравьев наблюдается в ГДР с мая по июнь (Otto, 1959 а; Rust, 1958), так как молодые личинки от них обычно ускользают. Гессвальд (Gosswald, 1962 b) подсчитал, что в очаге листовертки рабочие одного гнезда уничтожают за день в среднем 65 — 75 тыс. гусениц, около 28 тыс. куколок и около 26 тыс. бабочек. Во время вспышки массового размножения листовертки муравьи чаще посещают заселенные ею деревья и перестраивают систему дорожек. Форма охотпичьей территории при этом меняется (Gosswald, 1962 b; Gosswald, Kloft, 1956 a). Б. С. Щербаков (1953) пишет, что во время массового размножения дубовой листовертки в 1938 г. в Московской области муравьи одного среднего гнезда уничтожали в сутки около 7200 гусениц, а позднее, после окукливания — 6500 куколок вредителя. Истребление последних длилось около 20 дней, и за это время было уничтожопо более 10 тыс. куколок. Когда начался вылет бабочек, муравьи переключились на питание ими, причем ловили их, главным образом, в момент выхода из куколок. За 30 дней лёта было уничтожено более 150 тыс. взрослых бабочек.



 Плотность поселения муравьев в развитых комплексах гнезд (15— 20 м 2 оснований куполов на 1 га) может достигать огромных величин — около 1, 5 • 107 особей или свыше 100 кг биомассы на 1 га. Согласно имеющимся Данным по энергетике муравьев при такой численности они должны ежедневно получать с гектара угодий не менее 1 кг белковой пищи. Основную часть белковой пищи муравьи добывают, охотясь на насекомых — вредителей леса, сохраняя тем самым кроны деревьев от повреждения. Рыжие лесные муравьи обеспечивают сохранность насаждений от повреждения таких хозяйственно важных хвое- и листогрызущих вредителей, как сосновая совка, сосновая пяденица, зимняя пяденица, малый еловый пилильщик, рыжий сосновый пилильщик, обыкновенный сосновый пилильщик, ткач-пилильщик, березовый пилильщик, сосновый шелкопряд и др. Активно уничтожают рыжие лесные муравьи гусениц дубовой зеленой листовертки, монашенки, непарного шелкопряда, совкилишайницы, лиственничной минирующей моли и многих других.
Плотность поселения муравьев в развитых комплексах гнезд (15— 20 м 2 оснований куполов на 1 га) может достигать огромных величин — около 1, 5 • 107 особей или свыше 100 кг биомассы на 1 га. Согласно имеющимся Данным по энергетике муравьев при такой численности они должны ежедневно получать с гектара угодий не менее 1 кг белковой пищи. Основную часть белковой пищи муравьи добывают, охотясь на насекомых — вредителей леса, сохраняя тем самым кроны деревьев от повреждения. Рыжие лесные муравьи обеспечивают сохранность насаждений от повреждения таких хозяйственно важных хвое- и листогрызущих вредителей, как сосновая совка, сосновая пяденица, зимняя пяденица, малый еловый пилильщик, рыжий сосновый пилильщик, обыкновенный сосновый пилильщик, ткач-пилильщик, березовый пилильщик, сосновый шелкопряд и др. Активно уничтожают рыжие лесные муравьи гусениц дубовой зеленой листовертки, монашенки, непарного шелкопряда, совкилишайницы, лиственничной минирующей моли и многих других.




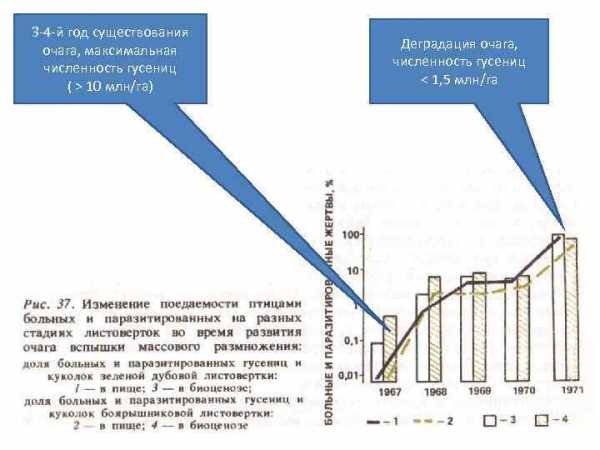 3 -4 -й год существования очага, максимальная численность гусениц ( > 10 млн/га) Деградация очага, численность гусениц
3 -4 -й год существования очага, максимальная численность гусениц ( > 10 млн/га) Деградация очага, численность гусениц

 Масса листового опада в широколиственных лесах от 3 до 5 тонн/га (в буковых лесах до 7 тонн/га) (сухой вес). Толщина подстилки за счет годового опада увеличивается в среднем на 2 – 3 см. Скорость разложения подстилки уменьшается в большей степени, чем количество опада, поэтому в широколиственных лесах происходит накопление лесной подстилки (100 -150 до 500 ц/га),
Масса листового опада в широколиственных лесах от 3 до 5 тонн/га (в буковых лесах до 7 тонн/га) (сухой вес). Толщина подстилки за счет годового опада увеличивается в среднем на 2 – 3 см. Скорость разложения подстилки уменьшается в большей степени, чем количество опада, поэтому в широколиственных лесах происходит накопление лесной подстилки (100 -150 до 500 ц/га),
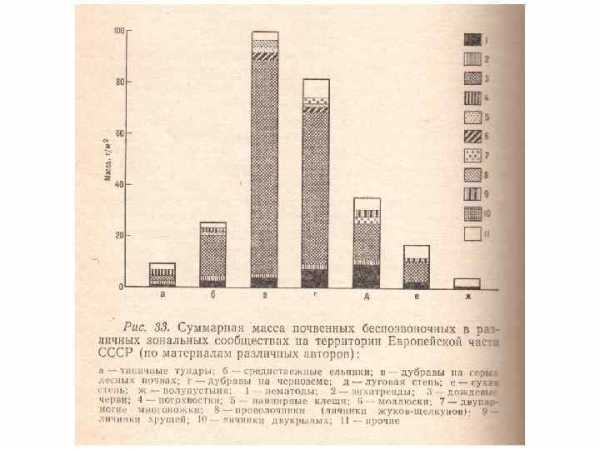
 Планетарные масштабы деятельности дождевых червей привлекли внимание Чарльза Дарвина еще в годы молодости. Его сочинение о дождевых червях появилось первым изданием в конце 1881 года, уже незадолго до его смерти. На одном гектаре общее количество дождевых червей составляло от 1 до 200 млн особей (в среднем около 20 млн), вес же биомассы — от 2 до 5 т/га, что почти в 100 раз превышает биомассу наземных животных на данной площади. За лето популяция из 100 червей на одном квадратном метре прокладывает в почве километр ходов, делая ее рыхлой, водо- и воздухопроницаемой. Установлено, что червь за сутки пропускает через пищеварительный канал количество земли с органикой, равное весу своего тела. Если принять средний вес червя в 0, 5 г, а количество их на 1 м 2 — 100 шт. (1 000 особей/га), то за сутки они пропустят 50 г на 1 м 2, или 0, 5 т/га. Активная деятельность червей продолжается в средней полосе 200 дней в году, значит количество почвы, прошедшей через их пищеварительный канал, выразится массой в 10 кг/м 2 (100 т/га).
Планетарные масштабы деятельности дождевых червей привлекли внимание Чарльза Дарвина еще в годы молодости. Его сочинение о дождевых червях появилось первым изданием в конце 1881 года, уже незадолго до его смерти. На одном гектаре общее количество дождевых червей составляло от 1 до 200 млн особей (в среднем около 20 млн), вес же биомассы — от 2 до 5 т/га, что почти в 100 раз превышает биомассу наземных животных на данной площади. За лето популяция из 100 червей на одном квадратном метре прокладывает в почве километр ходов, делая ее рыхлой, водо- и воздухопроницаемой. Установлено, что червь за сутки пропускает через пищеварительный канал количество земли с органикой, равное весу своего тела. Если принять средний вес червя в 0, 5 г, а количество их на 1 м 2 — 100 шт. (1 000 особей/га), то за сутки они пропустят 50 г на 1 м 2, или 0, 5 т/га. Активная деятельность червей продолжается в средней полосе 200 дней в году, значит количество почвы, прошедшей через их пищеварительный канал, выразится массой в 10 кг/м 2 (100 т/га).


 Кивсяки – многоножки (Diplopoda), многочисленные и неутомимые истребители листового опада в широколиственных и, в особенности, тропических лесах. Подобно дождевым червям, наиболее многочисленны и разнообразны в широколиственных и смешанных лесах (100 — 200 на 1 м 2) и единичны в тайге и горных хвойных лесах. Численность может достигать 400 особей на 1 м 2. В лесах где К. составляют значит, часть почвенной фауны, они обитают в лесной подстилке, верх, горизонтах почвы до глубины 30 см. Чувствительны к недостатку и избытку влаги, к содержанию кальция в почве, кислорода в окружающем воздухе, не переносят прямых солнечных лучей. Нижний слой лесной подстилки с выровненным гидротермическим режимом — наиболее благоприятная для них среда. Величина суточного рациона особи может в 1, 5 раза превышать массу её тела; при высокой численности К. способны перерабатывать до 50% разлагающегося за год опада. Высокая активность К. ускоряет темпы биол. круговорота и ведёт к повышению первичной продуктивности лесов.
Кивсяки – многоножки (Diplopoda), многочисленные и неутомимые истребители листового опада в широколиственных и, в особенности, тропических лесах. Подобно дождевым червям, наиболее многочисленны и разнообразны в широколиственных и смешанных лесах (100 — 200 на 1 м 2) и единичны в тайге и горных хвойных лесах. Численность может достигать 400 особей на 1 м 2. В лесах где К. составляют значит, часть почвенной фауны, они обитают в лесной подстилке, верх, горизонтах почвы до глубины 30 см. Чувствительны к недостатку и избытку влаги, к содержанию кальция в почве, кислорода в окружающем воздухе, не переносят прямых солнечных лучей. Нижний слой лесной подстилки с выровненным гидротермическим режимом — наиболее благоприятная для них среда. Величина суточного рациона особи может в 1, 5 раза превышать массу её тела; при высокой численности К. способны перерабатывать до 50% разлагающегося за год опада. Высокая активность К. ускоряет темпы биол. круговорота и ведёт к повышению первичной продуктивности лесов.



 Соня-полчок Сони – зимоспящие грызуны европейских широколиственных лесов. Практически всеядны, в рационе сочные плоды, ягоды, семена (желуди, орехи), насекомые, некоторые виды разоряют птичьи гнезда ( садовая соня)
Соня-полчок Сони – зимоспящие грызуны европейских широколиственных лесов. Практически всеядны, в рационе сочные плоды, ягоды, семена (желуди, орехи), насекомые, некоторые виды разоряют птичьи гнезда ( садовая соня)
 Садовая соня
Садовая соня
 Орешниковая соня
Орешниковая соня
 Японская соня – самый маленький представитель семейства. Семейство соневых – яркий пример европейско-восточноазиатского разрыва ареала, лежащего в области широколиственных лесов.
Японская соня – самый маленький представитель семейства. Семейство соневых – яркий пример европейско-восточноазиатского разрыва ареала, лежащего в области широколиственных лесов.
 Численность мышей достигает максимума (до 300 — 400 экз/га) на следующий год после урожая семян дуба, случающегося раз в 4 – 5 лет. В такие годы мыши съедают все семена, в том числе и поврежденные насекомыми, и в массе гибнут от бескормицы и эпизоотий, вызванных высокой плотностью. Желтогорлая мышь – характерный и многочисленный обитатель широколиственных лесов, занимающий важные позиции в биоценозе. Специализированный семеноед, в массе запасает и поедает семена дуба, лещины, липы, клена, ясеня, многих весенних эфемеров. Максимальная активность наблюдается в сентябре, в период опадания семян. В зависимости от численности популяции и урожая семян в разные годы изымают от 30 -40 до 100 % урожая. В одной кладовой находили до 10 кг желудей.
Численность мышей достигает максимума (до 300 — 400 экз/га) на следующий год после урожая семян дуба, случающегося раз в 4 – 5 лет. В такие годы мыши съедают все семена, в том числе и поврежденные насекомыми, и в массе гибнут от бескормицы и эпизоотий, вызванных высокой плотностью. Желтогорлая мышь – характерный и многочисленный обитатель широколиственных лесов, занимающий важные позиции в биоценозе. Специализированный семеноед, в массе запасает и поедает семена дуба, лещины, липы, клена, ясеня, многих весенних эфемеров. Максимальная активность наблюдается в сентябре, в период опадания семян. В зависимости от численности популяции и урожая семян в разные годы изымают от 30 -40 до 100 % урожая. В одной кладовой находили до 10 кг желудей.
 Лесная мышь широко распространена в зоне широколиственных лесов, но не столь тесно связана с высокоствольными сплошными насаждениями, обладающими максимальной продуктивностью. Чаще держится на опушках, полянах и на вырубках. Вглубь леса проникает в периоды депрессии численности желтогорлой мыши – более крупной и агрессивной.
Лесная мышь широко распространена в зоне широколиственных лесов, но не столь тесно связана с высокоствольными сплошными насаждениями, обладающими максимальной продуктивностью. Чаще держится на опушках, полянах и на вырубках. Вглубь леса проникает в периоды депрессии численности желтогорлой мыши – более крупной и агрессивной.
 Рыжая полевка – самый массовый грызун южной тайги, хвойно-широколиственных и широколиственных лесов. Питание смешанное – наряду с семенами деревьев в большом объеме потребляет семена травянистых растений и вегетативные части растений. Кормовая база более стабильна, чем у желтогорлой мыши.
Рыжая полевка – самый массовый грызун южной тайги, хвойно-широколиственных и широколиственных лесов. Питание смешанное – наряду с семенами деревьев в большом объеме потребляет семена травянистых растений и вегетативные части растений. Кормовая база более стабильна, чем у желтогорлой мыши.
 Подземная полевка – характерный грызун зоны широколиственных лесов в Европе. Норы в толстом слое лесной подстилки и в самом верхнем слое почвы. Питается в основном травянистыми растениями, весной — луковицами и корневищами и прикорневыми частями, летом — листьями, к осени в пище появляются семена, в частности желуди (даже незрелые). Отмечено поедание насекомых. Инстинкт запасания корма выражен сравнительно слабо.
Подземная полевка – характерный грызун зоны широколиственных лесов в Европе. Норы в толстом слое лесной подстилки и в самом верхнем слое почвы. Питается в основном травянистыми растениями, весной — луковицами и корневищами и прикорневыми частями, летом — листьями, к осени в пище появляются семена, в частности желуди (даже незрелые). Отмечено поедание насекомых. Инстинкт запасания корма выражен сравнительно слабо.

 Семена ясеня грызуны поедают неохотно, зато снегири – с удовольствием и в большом количестве.
Семена ясеня грызуны поедают неохотно, зато снегири – с удовольствием и в большом количестве.

 Белоногий хомячок – один из главных потребителей желудей в широколиственных лесах Северной Америки (запасает на зиму). Одновременно он является важным истребителем куколок дубового шелкопряда и служит резервуаром болезни Лайма.
Белоногий хомячок – один из главных потребителей желудей в широколиственных лесах Северной Америки (запасает на зиму). Одновременно он является важным истребителем куколок дубового шелкопряда и служит резервуаром болезни Лайма.


 Blue Jays carry food in their throat and upper esophagus—an area often called a “gular pouch. ” They may store 2 -3 acorns in the pouch, another one in their mouth, and one more in the tip of the bill. In this way they can carry off 5 acorns at a time to store for later feeding. Six birds with radio transmitters each cached 3, 000 -5, 000 acorns one autumn.
Blue Jays carry food in their throat and upper esophagus—an area often called a “gular pouch. ” They may store 2 -3 acorns in the pouch, another one in their mouth, and one more in the tip of the bill. In this way they can carry off 5 acorns at a time to store for later feeding. Six birds with radio transmitters each cached 3, 000 -5, 000 acorns one autumn.











 Белохвостый олень
Белохвостый олень



 При плотности 3 -5 лосей на 1000 га они изымают из насаждений до 10% доступного корма и не оказывают существенного влияния на процесс возобновления леса. При плотности 9 -10 лосей степень изъятия корма увеличивается до 25 – 60%, что уже губительно для отдельных видов деревьев. При плотности 12 – 18 лосей на 1000 га угодий излюбленные кома – ива и осина повреждаются на 80 -90%.
При плотности 3 -5 лосей на 1000 га они изымают из насаждений до 10% доступного корма и не оказывают существенного влияния на процесс возобновления леса. При плотности 9 -10 лосей степень изъятия корма увеличивается до 25 – 60%, что уже губительно для отдельных видов деревьев. При плотности 12 – 18 лосей на 1000 га угодий излюбленные кома – ива и осина повреждаются на 80 -90%.
present5.com
Широколиственные леса.
Широколиственные леса распространены в умеренном поясе Европы, на Дальнем Востоке, в Маньчжурии и на востоке Китая, в Северной Америке и небольшие площади занимают они в Средней Азии и в южной части Южной Америки.
Районы распространения ландшафтов широколиственных лесов характеризуются умеренно теплым климатом и оптимальным соотношение тепла и влаги. Хотя в этих ландшафтах период резкого зимнего спада в функционировании геосистем выражен достаточно отчетливо, период активной вегетации и биогенного метаболизма составляет 6-7 месяцев. На Евразийском континенте ареал этого ландшафта расширяется в сторону Атлантического и Тихого океанов и выклинивается к центру материка, прерываясь в Западной и Восточной Сибири. В Северной Америке эти ландшафты приурочены преимущественно к восточному побережью, выклиниваясь к центральной части материка и занимая узкую береговую зону на западном побережье. В Южной Америке они распространены в Патагонии.
Климатические условия развития этих ландшафтов различны, что позволяет подразделять их на ряд подзон, в зависимости от степени континентальности климата. Летние температуры составляют в среднем 18-200С (от 13 до 230С), сумма активных температур равна 2200-30000. Различия в зимних температурах по подзонам несколько больше — на западе они составляют в среднем -40С, а внутри континента до -140С. Величина годовых осадков также колеблется от 1000мм на западе до 700-600 мм в континентальных условиях. Максимум осадков приходится на летнее время. Коэффициент увлажнения составляет в среднем 1,2.
Благоприятные условия во время вегетационного периода дают возможность пышного развития растительности мезофильного типа. Листья на деревьях имеют широкие пластинки (бук, дуб, клен, липа), поэтому эти леса называют широколиственными. Размеры и форма листьев варьирует очень незначительно, что отличает их от деревьев тропического леса. Листья не приспособлены к зимнему неблагоприятному времени и опадают; в этом видят защиту деревьев от излишнего зимнего испарения (земля зимой холодна и является физиологически сухой). Стволы и ветви деревьев, в отличие от влажной тропической зоны, защищены толстой корой, а почки плотными чешуями, нередко смолистыми. Все это — также защита от зимнего испарения.
Широколиственные леса в большинстве случаев ярусные, можно различить два древесных яруса (один более высокий, другой -более низкий), кустарниковый, несколько травянистых, состоящих от трав различной высоты (до 3 ярусов), напочвенный ярус из мхов и лишайников. Но в некоторых фитоценозах (например, буковые леса) ярусность может отсутствовать и под плотными кронами деревьев нет ни подлеска ни травянистого покрова, а почва покрыта плотным слоем старых листьев. В отличие от тропических лесов, деревья верхнего яруса имеют одну и ту же высоту. Это зависит от очень небольшого числа видов деревьев, слагающих этот ярус. Нередко имеется вообще один вид и формируются чистые буковые, дубовые, грабовые леса, что сближает широколиственные леса с хвойными.
Очень характерен в широколиственных лесах световой режим. Здесь наблюдается два световых максимума; один весной, когда деревья еще не оделись листвой, другой — осенью, когда листва леса начинает изреживаться. В летние месяцы интенсивность света очень низкая. Подобный световой режим определяет особенности травянистого покрова. Ранней весной для этих лесов характерен резкий взрыв весенней растительности, состоящей из эфемеров-многолетников. Они очень быстро зацветают и затем также быстро заканчивают свою вегетацию. К июню они совершенно выходят из травянистого покрова, скрываясь под землей. Взрыв весенних эфемеров обычно очень красочен, так как цветки ярко окрашены, и их число так велико, что они покрывают землю сплошным покровом.
Лесообразующими породами в этих ландшафтах являются бук, дуб, клен, липа и каштан. В Западной Европе и Северной Америке доминируют различные виды бука и дуба, формирующие основные площади лесов. Дубовые леса в настоящее время уступают в площади буковым. Они приурочены к более сухим местообитаниям, эти леса менее тенистые и в них хорошо развивается подлесок и травянистый покров. Более влажные западноевропейские леса богаче в видов отношении. По мере продвижения на восток леса становятся беднее, исчезают граб, ясень. В древесном ярусе остаются дуб и липа (более морозоустойчивые), в травяном покрове значительное участие начинают принимать бореальные виды. В Северной Америке, доминирование в буковых лесах разделяет клен. Каштан появляется на бескарбонатных породах
Дальневосточные леса более разнообразны по видовому составу. Здесь наряду с местными видами дуба и клена, встречаются гледичия, магнолия, айлант, павловния, которые культивируются в Европе в парках. Есть в этих леса лианы, не свойственные в целом для зоны широколиственных лесов.
В настоящее время до 60-80% площади зоны широколиственных лесов распахано, что привело к сближению этих ландшафтов по характеру природных процессов со степными ландшафтами
Биомасса широколиственных лесов не намного ниже, чем влажных тропических лесов (3 000-5 000 ц/га), но продуктивность (П) и зеленая ассимилирующая масса (Б) меньше в несколько раз : П колеблется от 80 до 150 ц/га, Б в дубравах составляет 1% биомассы и достигает 40 ц/га (8% и 400 ц/га соответственно во влажных тропических лесах). К = 0,58-0,60 — больше, чем в тайге и меньше, чем во влажных тропических лесах.
Ежегодный растительный опад в несколько раз меньше, чем во влажных тропических лесах (50-70ц/га), из-за более низких температур и зимнего перерыва. меньше и темп его разложения Но скорость разложения уменьшается в большей степени, чем количество опада, поэтому в данных ландшафтах происходит накопление лесной подстилки (100-150 до 500 ц/га), которая практически отсутствует в тропиках. Подстилочный индекс равен 3-4 против 0,1 и менее во влажных тропических лесах, много накапливается гумуса (до 10% и более в горизонте А).
Зоомасса широколиственных лесов достигает 10-20 ц/га, около 90% общей зоомассы приходится на дождевых червей и 1% — на позвоночных.
Биологическая информативность этих ландшафтов ниже — число высших видов колеблется около 2000.
Широколиственные деревья богаты золой, особенно листья — до 5%, в ней много кальция — до 20%, (0,6-3,8% на сухое вещество), меньше калия — 0,15-2,0%, и кремния — 0,4-2,8%, еще меньше магния, алюминия, фосфора и меньше всего железа, марганца, натрия и хлора. При разложении растительного опада образуется значительное количество устойчивых органо-минеральных соединений.
По интенсивности биологического поглощения (АХ) последовательность иная:
10п п п-0,1п 0,п — 0,0п
S,P Ca, K, Mg, Mn Na Fe, Al, Si
Это предопределяет возможность биогенной аккумуляции в верхних горизонтах почв серы, фосфора, кальция, калия, магния и марганца, а также многих редких элементов, Ах которых больше 1 (бор, германий, никель, мышьяк, золото, берилл, кобальт, кадмий, свинец и др.). Но преобладание осадков над испаряемостью определяет большую роль выщелачивания и автономные ландшафты с вертикальным и боковым стоком теряют подвижные элементы. Высокое содержание в растениях кальция и его энергичное биологическое поглощение определяют кальциевый химизм БИКа. Кальций здесь типоморфный элемент.
Кальций и другие катионы, образующиеся при разложении растительных остатков, нейтрализуют большую часть органических кислот, в связи с чем реакция гумусового горизонта почв слабокислая или даже нейтральная, в поглощающем комплексе часто преобладает кальций.
Под широколиственными лесами формируются плодородные бурые (более влажные условия) или серые (более сухие и холодные условия) лесные почвы. Серые лесные почвы имеют мощный гумусово-аккумулятивный горизонт, слабовыраженный подзолистый, переходящий в иллювиальный. Присутствие в гумусовом горизонте, наряду с гуминовыми кислотами, фульвокислот способствует интенсивному выносу кальция. Поэтому верхние горизонты выщелочены от карбонатов.
В условиях более устойчивого увлажнения и большей теплообеспеченности под широколиственными лесами формируются бурые лесные почвы. Энергичное химическое выветривание приводит к оглинению почвенного профиля. Продукты гумификации представлены главным образом подвижными фульвокислотами, кальций легко мигрирует, карбонаты выщелачиваются и происходит относительное накопление алюминия и железа, дающих буроватую окраску почвам. Но почвенные растворы насыщены основаниями и придают почвам слабокислую и нейтральную реакцию и обеспечивают их высокое плодородие.
Влажный климат благоприятствует энергичному стоку. С наземным стоком выносится около 2,5-3,0 ц/га солей, несколько меньше, чем потребляется растительностью за год (3,5-5,0 ц/га), поэтому КВ здесь равен 1,3-1,4. С атмосферными осадками ежегодно поступает около 0,9-1,05 ц/га солей, которые включаются в БИК. Коэффициент атмогеохимической активности КА составляет 0,3-0,4. С ионным стоком ежегодно выносится значительно больше солей, чем поступает с атмосферными осадками, КИ- 3,4 — 4,9. Следовательно, основным источником солей в водах служит БИК и выветривание (80%).
Грунтовые и поверхностные воды в этих ландшафтах немного отличаются в зависимости от типа ландшафта. В Н-Са классах химических состав в основном зависит от разложения органических веществ, воды слабоминерализованы, менее 0,5г/л, гидрокарбонатно-кальциевые. В ландшафтах Са класса, где коры выветривания и континентальные отложения содержат СаСО3 в формировании химического состава вод, помимо БИК, важная роль принадлежит процессам растворения карбонатов. Воды более минерализованы, часто жесткие.
Таким образом, главные геохимические особенности ландшафтов широколиственных лесов состоят:
* в ежегодном продуцировании 80-150 ц/га живого вещества и средней скорости его разложения. При этом кислотные продукты распада частично нейтрализуются катионами, поступающими с растительными остатками. Реакция почв слабокислая, или близкая к нейтральной, что определяет невысокую подвижность биофильных элементов и гумуса;
* в биогенном накапливании в почвах многих элементов. Несмотря на вынос элементов из ландшафтов, особенно автономных, в почвах происходит некоторое накопление водных мигрантов, что связано с особенностями химического состава опада. Биогенная аккумуляция является в этих ландшафтах эффективным механизмом, стабилизирующим состав почв и повышающим их плодородие (отрицательная обратная связь).
* в усилении, по сравнению с влажными тропическими лесами, роли обратных отрицательных биокосных связей, и ослаблении биотических. Формирование подстилки в результате невысоких скоростей разложения опада и накопление биогенных элементов в почвах усиливают роль почв как среды миграции элементов в биогенной миграции, т.е. укрепляют обратные биокосные связи в ландшафте. БИК в широколиственных лесах улучшает условия существования организмов.
* прямые водные связи сильнее обратных и выщелачивание в целом преобладает. Но роль прямых нисходящих водных связей заметно меньше, чем во влажных тропиках. Элементы могут накапливаться не только в живых организмах, но в с опаде и гумусовом горизонте.
Прекрасные климатические условия ( достаточное количество тепла и постоянное увлажнение), сравнительно плодородные почвы определили важную роль равнинных ландшафтов зоны широколиственных лесов в сельском хозяйстве. Эти территории являются традиционно земледельческими. Первичные леса почти повсеместно вырублены и территории распаханы. Однако естественный химизм ландшафты (большая роль процессов выщелачивания) не обеспечивает необходимого уровня развития сельского хозяйства и высокой продуктивности растениеводства без постоянного внесения удобрений. Особенно большое значение имеют азотные и фосфорные удобрения, местами также калийные. К числу дефицитных элементов относятся азот, фосфор, калий, местами кобальт, медь, цинк, марганец, йод, молибден и другие микроэлементы. Избыточных элементов в ландшафтах нет.
Среди ландшафтов широколиственных лесов выделяют два основных класса: переходные от кислого к кальциевому (Н+ — Са2+) и кальциевые ландшафты.
www.geograf-stud.ru
Зона широколиственных лесов Рязанской области
В отличие от подтаежной зоны, где антропогенная нагрузка на ландшафты не привела к сколько-нибудь существенному изменению их характерных особенностей, зона широколиственных лесов на территории области в определенной мере утратила свою специфику. Дубравы, которые еще в XVII в. почти сплошной полосой в 40 – 90 км протягивались по правобережью Оки до р. Пары и далее через Окско-Цнинское плато на правобережье р. Цны, ныне практически сведены. Занятые ими ранее междуречья на большей их части распаханы, на меньшей – поросли вторичными березово-осиповыми лесами с примесью дуба, липы, клена, ясеня. Местами сформировались рощи из дуба с примесью липы и других лиственных пород. За исключением правобережья р. Цны ландшафты приобрели лесостепной характер. Под лесами к западу от р. Цны сейчас занято в среднем 18 % площади этого участка зоны.
Значительную трансформацию в процессе земледелия претерпели и характерные для зоны широколиственных лесов серые лесные почвы. Многовековая их вспашка привела к изменению профиля почв, осреднению содержания гумуса в пахотном слое, смыву гумусового горизонта на пологонаклонных придолинных участках междуречий.
Границу между зоной широколиственных лесов и лесостепью мы проводим с учетом распространения оподзоленных и выщелоченных черноземов, относя даже небольшие их «пятна» к лесостепи. Таким образом, в реконструируемую широколиственных лесов вошел пояс серых лесных почв па покровных лессовидных суглинках с вытянутыми в меридиональном направлении массивами дерново-подзолистых почв в пределах долинных и придолинных зандров па правобережьях pp. Рановы, Пары и Цны.
Суммарная солнечная радиация в пределах этой зоны на территории области 90 – 93 ккал/см- • год, радиационный баланс 35 – 37 ккал/см2 • год. По количеству осадков зона широколиственных лесов несколько уступает подтайте. Среднегодовая их величина здесь составляет 450 – 500 мм, из них па теплый период приходится – 300 – 350 мм. С мая по сентябрь испаряемость здесь превышает количество выпадающих осадков. В апреле – мае и сентябре – октябре испаряемость меньше, чем выпадает осадков. Коэффициент увлажнения в период с начала апреля до конца октября в среднем несколько больше 1. Модуль стока меняется от 3,5 до 4 л/сек/км2, слой стока -от 1 Юдо 130 мм. При этом от 90 до 80 мм на разных участках приходится на период снеготаяния.
Продолжительность периода со среднесуточной температурой выше +10 °С (периода вегетации) составляет 140 – 143 дня, с температурой выше + 15 °С (лето) 85 -90 дней. Сумма активных температур колеблется от 2200 до 2300 °С.
Из -за распаханности большей части междуречий эрозионные процессы по своему характеру здесь ближе к лесостепным и степным, чем к подтаежным. Мутность речных вод достигает 100 г/м\ а весной и более (в Мещере всего 20 -30 г/м ). Модуль твердого стока составляет 10 -20 т/ км- год. Поверхностные воды по сравнению с подтайгой (0,2 – 0,3 г/л) более минерализованы – до 0,5 г/л.
Фитомасса современных лесов 300 – 400 т/га, местами более. Ежегодный прирост 8-10 т/га, опад 5 – 8 т/га [7], истинный прирост 2-4 т/га, ежегодно потребляется до 300 кг/га химических элементов, с опадом возвращается 200 – 250 кг/га. В опаде содержится большое количество кальция, калия и азота, а также фосфора и серы.
Зоомасса достигает 1000 – 2000 кг/га. В основном это почвенные сапрофаги (около 90 % дождевые черви). При разложении растительного опада образуется большое количество устойчивых органо-минеральных соединений, однако оснований в почве недостаточно для полной нейтрализации кислых продуктов разложения (рН = 5,7- 6,8). Светло-серые почвы весьма близки к дерново-подзолистым (иод маломощным гумусово-аккумулятивным горизонтом выражен оподзоленный горизонт). У серых и темно-серых лесных почв гумусово-аккумулятивный горизонт мощностью до 40 – 60 см постепенно переходит в иллювиальный. Темно-серые лесные почвы богаты гуминовыми кислотами, но содержат и фульвокислоты, что способствует выщелачиванию верхних горизонтов.
В условиях антропогенного прессинга наибольшую трансформацию претерпели самые молодые и динамичные компоненты ПК -растительность и почвы. Заметно изменился режим стока поверхностных вод. В меньшей мере антропогенное влияние сказалось на рельефе, который формировался на протяжении многих тысяч лет, причем в отдельные эпохи при совсем иных, нежели сейчас, физико-географических условиях. Сохранила свои основные особенности и литогенная основа, представленная средне-поздпеплейстоценовыми и голоцен о вы ми отложениями разного генезиса.
Пространственная дифференциация антропогенно измененных ландшафтов в пределах зоны широколиственных лесов, таким образом, связана с неоднородностью ведущих компонентов рельефа и субстрата.
В границах зоны широколиственных лесов с учетом существующих различий в рельефе и субстрате нами выделены две провинции: Среднерусская, расположенная в пределах Среднерусской возвышенности и Окско-Донская – в пределах одноименной равнины.
rznp.ru