Типы лесорастительных условий — Аналитика Лесной промышленности
Как известно, без леса мы не сможем существовать. Древесина применяется в различных областях промышленности. Типология лесорастительных условий появилась еще в начале двадцатого века. В этой статье мы разберем, какие же типы бывают.
Объединение участков леса
Классификация, о которой пойдет речь, была представлена ученым В.Н. Сукачевым. По его мнению, типы леса – это определенный тип биогеоценоза. Участки леса объединяются в определенные группы. А эти группы и называют биогеоценозом.
Участки леса могут объединяться по различным признакам:
- по однородности или неоднородности пород;
- по определенным растительным ярусам;
- по наличию или отсутствию микроорганизмов;
- по различным климатическим условиям;
- по состоянию почвы;
- по тому, как растения приспосабливаются к окружающей среде;
- по способам восстановления и обмену в лесах.

Типы леса по В.Н. Сукачеву
В.Н. Сукачев создал свою типологию леса, состоящего из сосен и елей, которые расположены в южной тайге.
Ельники и сосняки расположены на очень хороших, плодородных и поддающиеся дренажу землях. Ученый выделил один вид лесов – стержневой. Он самый характерный.
К зеленомошной группе относятся сосны и ели. Почвенный покров в таких местах отличается дренированностью, и рельеф хороший.
Если рассматривать долгомошную группу, то можно сделать вывод о том, что почва богатая, но присутствуют болота, а рельеф развит не очень хорошо.
Есть, также, сфагновые сосняки и ельники, которые расположены на котловинном дне или на равнине.
Если тип леса травяной, то вы должны знать, что он расположен на болотистой почве, хотя там должна находиться проточная вода.
В сосновых лесах можно встретить лишайниковые боры, в который почвенный покров сухой. При таких условиях ель расти не может. Также, в сосновом лесу очень сильно заболачивается местность.
Существует два вида ельников: травяно-сфанговые и оско-сфагновые. Первый тип бывает на почвах, достаточно, плодородных, а второй тип растет в котловинах, где протекает небольшой медленный водоем.
Лесной вид отличает определенный вид подлеска. По этим видам и называют типы лесов.
По Сукачеву бывают коренные и производные леса. К коренным относится лес с елями, а к производным – леса с соснами. Исключением могут быть лишайники и сфагновые. Сосняки появляются на месте ельников после рубки и пожаров.
Производный вид делится на первично-производный лес и вторично-производный лес.
К первому типу относится кисличный, брусничный и черничный сосняк.
Когда происходят какие-либо изменения в первично-производном типе, то появляются вторично-производные.
Какие бывают главные группы
Можно выделить лишайниковую группу типов леса, как главную. Леса на таких почвах сухие, бедные. Почва либо с песчаная, либо каменистая. Можно сказать, что лес, достаточно, редкий.
Подлеска можно не увидеть, либо он будет очень редкий. Из живого покрова, который преобладает, можно выделить лишайники.
Процесс восстановления леса очень медленный или может вообще не происходить, так как почва очень сухая и бедная, как было сказано выше.
Еще одна важная группа – зеленомошный тип леса. Здесь растут сосняки, ельники, березняки, разные виды осинников.
Осинники кисличные растут на свежих супесях или суглинках. Зачастую, это смешенные леса, но достаточно продуктивные. Живой покров включает в себя полевые и лесные трав. Если есть вырубки, то на них начинают расти злаки.
Осинники черничные растут на влажных почвах. Дренажа на таких почвах, практически, нет. Леса бывают, как смешанные, так и чистые. Древесина очень высокого качества. В подлесках наблюдается рябина, которая вырастает до средних размеров.
Существуют различные зоны микроповышения и микропонижения. Вырубки в таких местах очень быстро превращаются в болотистые местности, которые зарастают мхом.
Брусничники, в свою очередь, занимают зону сухих и песчаных почв. Зачастую там растут сосны, редко бывают смешанные леса. Если же лес не смешенный, то к соснам присоединяются березы или ели. Брусника и мхи представляют лесной покров. Подлесок состоит из рябины и очень редкий. Древесина сосны достаточно качественная. Вырубки зарастают злаковыми растениями.
Долгомошная группа растет на сырых почвах, которые, зачастую, торфяные и заболоченные. Растут там сосняки, березняки и ельники. Зачастую лес представлен чистым, без примесей. Древесина среднего качества. Вырубки становятся заболоченными. В таких местах, вряд ли, вырастут какие-либо хвойные деревья. На их месте вырастают березы и сосны.
К сфагновым лесам относятся сосняки, березняки и ельники. Растут они на торфяниках. Деревья расположены редко, и они не смешанные. Вырубки становятся болотистыми землями.
Травяной лесной тип составляют следующие деревья: ель, береза, ольха и сосна. Почва заболоченная. Подлесок большой.
Еще один тип – сложный. Почвы для этого типа глинистые или песчаные. Произрастать здесь будут липа, клен и другие виды. Сосны в таких лесах имеют не особо качественное дерево. Покров почвы незаметный, так как солнечный свет почти не проникает в такие леса.
Типология Г.Ф. Морозова
Основоположником всего учения о различных типах лесов был ученый Г.Ф. Морозов. Он различал типы лесов по тому, какая почва в тех или иных местах. Эти условия непосредственно связывают с климатом. Более менее одинаковые почвенные и климатические условия можно объединить в один лесной тип.
Под определенной группой леса Морозов понимал сходство лесных участков, имеющих в своем составе одинаковый состав деревьев, которые, в свою очередь, растут в одинаковых условиях среды. Однородность должна быть, как морфологическая, так и внутренняя.
До Морозова тоже разграничивали различные типы леса по их почвенному составу и древесным породам. Например, выделяли боры, дубравы и так далее. Эти названия использовали для лесоописаний. Но только Морозов сумел создать полную классификацию и найти взаимосвязь почвенного покрова с древесным составом.
Эти названия использовали для лесоописаний. Но только Морозов сумел создать полную классификацию и найти взаимосвязь почвенного покрова с древесным составом.
В дальнейшем, Морозов ввел биогеоценотическое направление, которое возглавил уже В.Н. Сукачев. В его типологии учитывался тип почв, растительный покров, какие виды деревьев произрастают, какая существует тенденция к смене лесных пород.
Морозов создал монографию, где подробно рассказывается его типология. Благодаря его труду, развитие типологии продолжилось.
Типология леса по П.С. Погребняку
П.С. Погребняк тоже занимался типологией лесов. Он строил эдафическую сетку, учитывая влажность земли и то, плодорна ли почва или нет.
Погребняк выделяет следующие виды почвенного состояния: очень сухие, сухие, влажные, сырые, болота.
По типу плодородия он выделяет: боровые, субори, сложные субори, дубравные леса.
Когда происходит сочетание гигротопов и трофотропов, то получаются эдатопы.
Эдафичская сетка нужна для характеристики условий, в которых растут леса.
Типология вырубок леса
И.С. Мелехов разработал свою типологию вырубок. Она представлена разными типами лесов, которые существуют и, которые еще вырастут.
Типология показывает, где лучше вырубать лес. Она учитывает однородность леса, покров, время, которое требуется, чтобы лес восстановился. Это все нужно для того, чтобы знать, как долго будет восстанавливаться лес, и возможна ли смена лесных пород.
В каждом типе условий может быть несколько видов вырубки. Соответственно, если лес богатый, то и типология здесь будет присутствовать в большем объеме. Если же, почва чересчур увлажнена или, наоборот, очень сухая, то здесь будет один вид какой-либо вырубки.
Если в лесу был низовой пожар, то типов вырубок здесь будет много. В таком случае можно выделить две группы вырубок: после воздействия огня и без огня.
Важно учитывать при различных вырубках состав леса, какова растительность в нижних ярусах и многие другие факторы. Именно из-за этого типов лесов значительно меньше, чем типов вырубки.
Без изучения типов леса невозможно описать лес и изучить его. Лес, ведь, это не просто растущие деревья и кустарники, а определенное сообщество деревьев и растений, которое должно быть хорошо изучено. Ведь без этого невозможно определить, где можно вырубать леса, где нельзя. А также, нельзя точно сказать, какое количество времени леса будут восстанавливаться. Поэтому вы должны понимать, что создание типологий было важным этапом в развитии науки о лесе.
Лесная типология
Учение о типах леса, рассматривающее вопросы диагностирования, выделения, классификации типов леса и типов лесорастительных условий как естественно-исторической основы лесного хозяйства. Основателем этого учения является Г. Ф. Морозов, создавший учение о типах насаждений: «Тип насаждений есть совокупность насаждений, объединенных в одну обширную группу общностью условий местопроизрастания или почвенно-грунтовых условий». Это первое определение типа насаждений (леса) — основной классификационной единицы Л.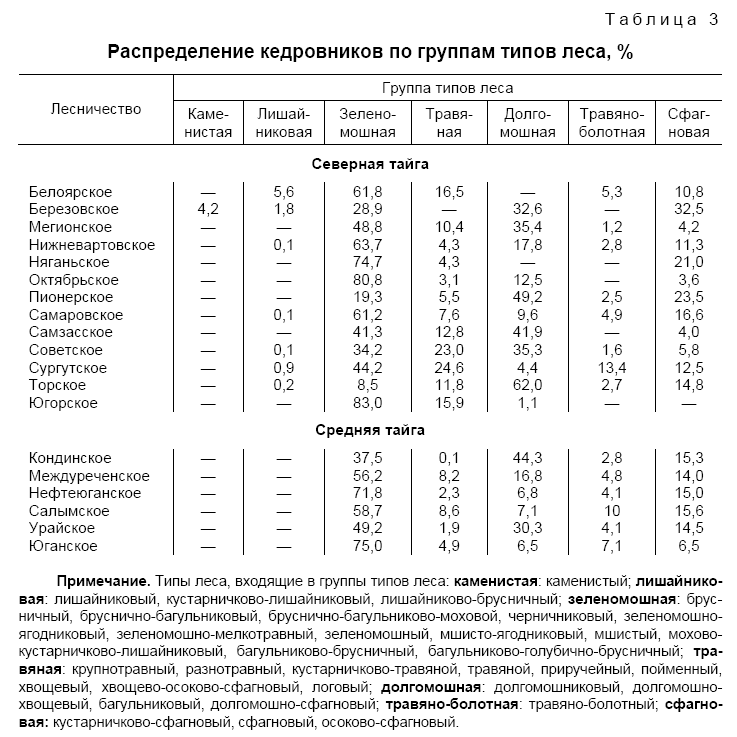 т. В дальнейшем Г. Ф. Морозов предложил классифицировать лесные сообщества на основе совокупности всех факторов лесообразования (см. Лес). Дальнейшее развитие Л. т. связано с именем В. Н. Сукачева. Он дополнил характеристику типа леса показателем производительности условий местопроизрастания по бонитировочной таблице, используя «среднюю высоту господствующих стволов нормального роста». При этом он допускал в лесах одного типа и возраста различие средних высот в пределах 3-4 м, что соответствует амплитуде высот в одном классе бонитета. По В. Н. Сукачеву, «тип леса — это объединение участков леса (т. е. отдельных лесных биогеоценозов), однородных по составу древесных пород, по другим ярусам растительности и фауне, по микробному населению, климатическим, почвенным и гидрологическим условиям, взаимоотношениям между растениями и средой, внутрибиогеоценотическому обмену веществом и энергией, восстановительным процессам и направлению смен в них. Эта однородность свойств компонентов биогеоценозов и свойств биогеоценозов в целом, объединяемых в один тип, требует при одинаковых экономических условиях применения и одинаковых лесохозяйственных мероприятий».
т. В дальнейшем Г. Ф. Морозов предложил классифицировать лесные сообщества на основе совокупности всех факторов лесообразования (см. Лес). Дальнейшее развитие Л. т. связано с именем В. Н. Сукачева. Он дополнил характеристику типа леса показателем производительности условий местопроизрастания по бонитировочной таблице, используя «среднюю высоту господствующих стволов нормального роста». При этом он допускал в лесах одного типа и возраста различие средних высот в пределах 3-4 м, что соответствует амплитуде высот в одном классе бонитета. По В. Н. Сукачеву, «тип леса — это объединение участков леса (т. е. отдельных лесных биогеоценозов), однородных по составу древесных пород, по другим ярусам растительности и фауне, по микробному населению, климатическим, почвенным и гидрологическим условиям, взаимоотношениям между растениями и средой, внутрибиогеоценотическому обмену веществом и энергией, восстановительным процессам и направлению смен в них. Эта однородность свойств компонентов биогеоценозов и свойств биогеоценозов в целом, объединяемых в один тип, требует при одинаковых экономических условиях применения и одинаковых лесохозяйственных мероприятий».
 Являясь продолжателем нек-рых концепций А. А. Крюденера, Е. В. Алексеев объединил все разнообразие типов леса по почвеннофунтовым условиям в 6 групп: 4 из них на суходолах (боры — песчаные почвы; субори — в основном супеси; груды — суглинки; дубравы — черноземные почвы) и 2 группы в мокрых местообитаниях (ольшаники с низинным торфом и багны с моховым торфом). Суходольные группы он подразделил, так же по влажности почвы, на основные типы леса: сухой бор, свежий бор, влажный бор, сырой бор и т. д. П. С. Погребняк объединил типы леса на суходолах и в мокрых местообитаниях в один почвенный ряд. Была построена эдафическая сетка по двум ведущим факторам почвенного плодородия — богатству (трофотопы) и влажности (гигротопы) почвы. Но и в одном трофотопе почвы могут быть неоднородными по механическому составу, но однородными по богатству: боры (А) — крайне бедные почвенно-грунтовые условия; простые субори (В) — относительно бедные; сложные субори (С) — относительно богатые; дубравы (D) — наиболее плодородные почвы.
Являясь продолжателем нек-рых концепций А. А. Крюденера, Е. В. Алексеев объединил все разнообразие типов леса по почвеннофунтовым условиям в 6 групп: 4 из них на суходолах (боры — песчаные почвы; субори — в основном супеси; груды — суглинки; дубравы — черноземные почвы) и 2 группы в мокрых местообитаниях (ольшаники с низинным торфом и багны с моховым торфом). Суходольные группы он подразделил, так же по влажности почвы, на основные типы леса: сухой бор, свежий бор, влажный бор, сырой бор и т. д. П. С. Погребняк объединил типы леса на суходолах и в мокрых местообитаниях в один почвенный ряд. Была построена эдафическая сетка по двум ведущим факторам почвенного плодородия — богатству (трофотопы) и влажности (гигротопы) почвы. Но и в одном трофотопе почвы могут быть неоднородными по механическому составу, но однородными по богатству: боры (А) — крайне бедные почвенно-грунтовые условия; простые субори (В) — относительно бедные; сложные субори (С) — относительно богатые; дубравы (D) — наиболее плодородные почвы. П. С. Погребняк выделил также 6 гигротопов: 0 — крайне сухие местообитания, 1 — сухие, 2 — свежие, 3 — влажные, 4 — сырые, 5 — лесные болота. Каждый участок леса характеризуется двумя классификационными единицами — трофотопом и гигротопом. В единстве они составляют эдатоп — участок леса с одинаковыми почвенно-грунтовыми условиями. Б. П. Колесников разработал генетическую классификацию типов леса. В основу ее положены закономерности процессов возникновения и развития леса. Тип леса при генетическом подходе рассматривается как этап лесообразовательного процесса, к-рый объединяет в границах какого-либо одного типа лесорастительных условий лесные участки. Эти участки представлены насаждениями, к-рые находятся на последовательно сменяющихся стадиях возрастного или восстановительного развития древостоев лесообразующей породы (или пород в смешанных лесах). Каждому типу лесорастительных условий соответствуют: высота над уровнем моря, форма рельефа, крутизна склона, экспозиция склона, генетический тип почвы, гранулометрический состав почвы, степень и режим влажности, мощность почвенного профиля, — определяющие потенциальную биологическую продуктивность леса.
П. С. Погребняк выделил также 6 гигротопов: 0 — крайне сухие местообитания, 1 — сухие, 2 — свежие, 3 — влажные, 4 — сырые, 5 — лесные болота. Каждый участок леса характеризуется двумя классификационными единицами — трофотопом и гигротопом. В единстве они составляют эдатоп — участок леса с одинаковыми почвенно-грунтовыми условиями. Б. П. Колесников разработал генетическую классификацию типов леса. В основу ее положены закономерности процессов возникновения и развития леса. Тип леса при генетическом подходе рассматривается как этап лесообразовательного процесса, к-рый объединяет в границах какого-либо одного типа лесорастительных условий лесные участки. Эти участки представлены насаждениями, к-рые находятся на последовательно сменяющихся стадиях возрастного или восстановительного развития древостоев лесообразующей породы (или пород в смешанных лесах). Каждому типу лесорастительных условий соответствуют: высота над уровнем моря, форма рельефа, крутизна склона, экспозиция склона, генетический тип почвы, гранулометрический состав почвы, степень и режим влажности, мощность почвенного профиля, — определяющие потенциальную биологическую продуктивность леса. И. С. Мелехов сформулировал задачи динамической типологии, показав, что в развитии типа леса можно выделить этапы, предшествующие его образованию (тип вырубки, гари), — этапы формирующегося типа леса (включая промежуточные или переходные типы), этапы сложившегося типа леса (в спелом возрасте древостоя), возможные последующие этапы с переходом или без перехода в новый тип леса — и подчеркнул, что изучение последовательности различных этапов лесообразования и типологических связей не только в пространстве, но и во времени — важнейшая задача лесоведения и лесоводства. В лесохозяйственной практике России используется в основном, типология В. Н. Сукачева и классификация П. С. Погребняка. При лесоустроительных работах в каждом таксационном выделе определяется наиболее распространенный тип лесного биогеоценоза и тип лесорастительных условий. За рубежом существует несколько направлений Л. т. Наиболее известна типология финского ученого А. К. Каяндера. Типы леса, по его определению, соответствуют типам лесорастительных условий, экологические режимы к-рых устанавливаются по напочвенному покрову без учета лесообразующей породы.
И. С. Мелехов сформулировал задачи динамической типологии, показав, что в развитии типа леса можно выделить этапы, предшествующие его образованию (тип вырубки, гари), — этапы формирующегося типа леса (включая промежуточные или переходные типы), этапы сложившегося типа леса (в спелом возрасте древостоя), возможные последующие этапы с переходом или без перехода в новый тип леса — и подчеркнул, что изучение последовательности различных этапов лесообразования и типологических связей не только в пространстве, но и во времени — важнейшая задача лесоведения и лесоводства. В лесохозяйственной практике России используется в основном, типология В. Н. Сукачева и классификация П. С. Погребняка. При лесоустроительных работах в каждом таксационном выделе определяется наиболее распространенный тип лесного биогеоценоза и тип лесорастительных условий. За рубежом существует несколько направлений Л. т. Наиболее известна типология финского ученого А. К. Каяндера. Типы леса, по его определению, соответствуют типам лесорастительных условий, экологические режимы к-рых устанавливаются по напочвенному покрову без учета лесообразующей породы. В Западной Европе возникло учение о типах местообитания. Каждая единица местообитания, как экологическая система, характеризуется особенностями рельефа, почвы (общая мощность, гранулометрический состав, структура, кислотность, водоудерживающая способность), микроклимата, состава растительных ярусов. В др. странах лесоводы используют геоботаническую классификацию с выделением растительных ассоциаций.
В Западной Европе возникло учение о типах местообитания. Каждая единица местообитания, как экологическая система, характеризуется особенностями рельефа, почвы (общая мощность, гранулометрический состав, структура, кислотность, водоудерживающая способность), микроклимата, состава растительных ярусов. В др. странах лесоводы используют геоботаническую классификацию с выделением растительных ассоциаций.Типы леса | Экология, лес и почва
В основу рассматриваемой классификации типов лесного участка положены экологические факторы: минеральное богатство почв, или трофность (от греческого слова «трофос» — пища), влажность почв и климат. В пределах одного климатического района местное разнообразие лесов будет зависеть от изменения плодородия почвы и влажности почвогрунта. Эти две координаты составляют основу эдафической, т. е. почвенной сетки классификации типов лесного участка.
Эдафическая сетка для классификации типов лесного участка по Е. В. Алексееву и П. С. Погребняку.
Алексееву и П. С. Погребняку.
| Группы влажности | Группы богатства почвы | |||
| А боры | В субори | С судубравы | Д дубравы | |
| 0 — очень сухие | А0 | В0 | С0 | Д0 |
| 1 — сухие | А1 | В1 | С1 | Д1 |
| 2 — свежие | А2 | В2 | С2 | Д2 |
| 3 — влажные | А3 | В3 | С3 | Д3 |
| 4 — сырые | А4 | В4 | С4 | Д4 |
| 5 — мокрые | А5 | В5 | С5 | Д5 |
В приведенной сетке по горизонтали слева направо возрастает степень плодородия почвы от А до Д, образуя трофогенный ряд. В этом ряду условно выделено четыре градации по плодородию почвы для лесных участков: А — боры; В — субори; С — судубравы, сугрудки; Д — дубравы, груды, которые называются трофотопами. По вертикали сверху вниз возрастает степень увлажнения, образуя гигрогенный ряд. В этом ряду выделяют шесть градаций влажности почвы: О — очень сухие, 1 — сухие, 2 — свежие, 3 — влажные, 4 — сырые, 5 — мокрые. Эти группы называют гигротопами. Пересечение рядов трофотопов и гигротопов образует эдатоп (буквально: эдафическое, или почвенное, место), т. е. тип лесного участка. Названия эдатопов образуются из соответствующих названий трофотопов и гигротопов. Например: Ах— сухой бор, В2— свежая суборь, Д3— влажная дубрава и т. д.
В этом ряду условно выделено четыре градации по плодородию почвы для лесных участков: А — боры; В — субори; С — судубравы, сугрудки; Д — дубравы, груды, которые называются трофотопами. По вертикали сверху вниз возрастает степень увлажнения, образуя гигрогенный ряд. В этом ряду выделяют шесть градаций влажности почвы: О — очень сухие, 1 — сухие, 2 — свежие, 3 — влажные, 4 — сырые, 5 — мокрые. Эти группы называют гигротопами. Пересечение рядов трофотопов и гигротопов образует эдатоп (буквально: эдафическое, или почвенное, место), т. е. тип лесного участка. Названия эдатопов образуются из соответствующих названий трофотопов и гигротопов. Например: Ах— сухой бор, В2— свежая суборь, Д3— влажная дубрава и т. д.
Рассмотрим, как меняется состав и форма леса в ряду трофности. Здесь от бора к дубравам нарастает плодородие почвы и соответственно этому изменяется облик леса. Состав, форма и производительность коренных насаждений меняются примерно так:
А — боры. Одноярусные сосновые с примесью березы насаждения средних или низких бонитетов на песках; в подлеске можжевельник, вереск, ягодники; в живом покрове багульник, лишайники, зеленые мхи.
Одноярусные сосновые с примесью березы насаждения средних или низких бонитетов на песках; в подлеске можжевельник, вереск, ягодники; в живом покрове багульник, лишайники, зеленые мхи.
В — субори. Двухъярусные сосново-дубово-березовые насаждения (сосна высокого бонитета) на глинистых песках; в подлеске, кроме боровых видов, рябина; в покрове грушанки, папоротник орляк и др.
С — судубравы. Трех-четырехъярусные насаждения из сосны высших бонитетов и березы в I ярусе; в подчиненных ярусах дуб, липа, клен, граб. В подлеске, кроме суборевых видов, лещина, жимолость, бересклет; в покрове кислица, звездчатка, ясменник, сныть и др.
Д — дубравы. Многоярусные насаждения на суглинистых и глинистых почвах. В подлеске преобладает лещина с примесью бересклетов европейского и бородавчатого, клена татарского, свидины, боярышника, бузины, бирючины, крушины и др.; в покрове сныть, копытень, чина весенняя, колокольчик крапиволистный, осока волосистая, ежа сборная, гравилат городской, медуница, звездчатка, фиалка и др. В тайге место дубрав занимают рамени, т. е. ельники.
В тайге место дубрав занимают рамени, т. е. ельники.
Каждая группа трофотопов делится на типы участков по степени увлажнения. Поскольку большинство древесных видов растений является мезофитами, то изменение увлажнения почвогрунтов прежде всего сказывается на продуктивности насаждений. Она будет возрастать от сухих условий к свежим и влажным, а затем убывать к сырым и мокрым. В ряде случаев при этом будет меняться и видовой состав (ольшаники на болотах), а также соотношение между видами растений. Условия увлажнения в значительной степени связаны с рельефом местности и глубиной залегания грунтовых вод.
Классификация типов леса строится для каждого типа лесного участка (эдатопа), что позволяет сравнивать типы леса разных климатических районов и дает возможность обобщать передовой опыт ведения лесного хозяйства и внедрения его в сходных условиях среды.
Биогеоценоз, тип леса, тип лесорастительных условий » Строительный онлайн-ресурс
16.01.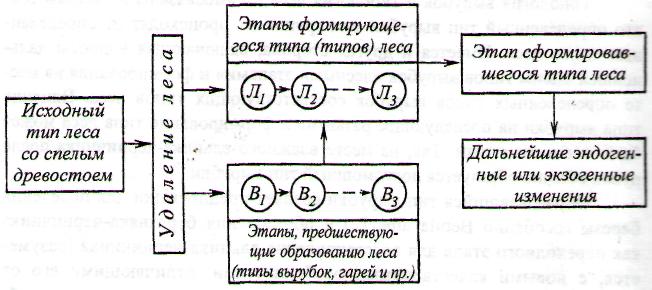 2014
2014 По определению академика В.Н. Сукачева, биогеоценоз — «совокупность на определенном протяжении земной поверхности однородных природных явлений атмосферы, горной породы, растительности, животного мира и мира микроорганизмов, почвы и гидрологических условий, имеющая свою особую специфику взаимодействия этих слагающих ее компонентов и определенный тип обмена веществом и энергией их между собой и с другими явлениями природы и представляющая собой внутреннее противоречивое единство, находящееся в постоянном движении, развитии».
По В.Н. Сукачеву, биогеоценоз (БГЦ) включает пять основных компонентов: 1) фитоценоз; 2) зооценоз; 3) микробоценоз; 4) эдафотоп; 5) климатоп.
Первые три компонента составляют органическую часть БГЦ — его биоценоз, а эдафотоп и климатоп — неорганическую среду. Наиболее важная часть БГЦ — фитоценоз, который создает органику среды, тогда как зооценоз и микробоценоз — потребители этого вещества. Поэтому классификация БГЦ построена на классификации фитоценозов.
Биогеоценотический подход к лесу как лесному биогеоценозу четко отражен в его современном определении. Лес (по ГОСТ 18486 — 87) — элемент географического ландшафта, состоящий из совокупности древесных, кустарниковых, травянистых растений, животных и микроорганизмов, в своем развитии биологически связанных, влияющих друг на друга и на внешнюю среду. Согласно положению «Основы лесного законодательства РФ (1993)», лес — это совокупность земли, древесной, кустарниковой и травянистой растительности, животных, микроорганизмов и других компонентов окружающей среды, биологически взаимосвязанных и влияющих друг на друга в своем развитии. Сходные по своим показателям биогеоценозы объединяют в тип лесного биогеоценоза, который лесоводы понимают как тип леса. В соответствии с ГОСТ 18486—87 тип леса — участок леса или их совокупность, характеризующийся общим типом лесорастительных условий, одинаковым составом древесных пород, количеством ярусов, аналогичной фауной, требующих одних и тех же лесохозяйственных мероприятий при равных экономических условиях.
Отдельным типам леса дают соответствующее бинарное название. Оно может соответствовать названию доминирующей в данном типе леса лесной ассоциации (сосняк лишайниковый, ельник черничный, сосняк с дубом и т.д.), но может быть определено и по-другому (ельник-лог, сосняк сложный, осинник приручейниковый, черноольховник приречный и т.д.).
He всегда территории, характеризующиеся определенными условиями местопроизрастания, заняты именно лесной растительностью. На этой территории могут находиться ассоциации других типов растительности или временно растительность может быть сильно нарушена (вырубка леса, лесные пожары и т.д.). В подобных случаях условия местопроизрастания принято рассматривать и классифицировать путем выделения типов лесорастительных условий, которые по ГОСТ 18486—87 представляют собой совокупность однородных лесорастительных условий на покрытых и не покрытых лесом участках.
Тип лесорастительных условий, или тип условий местопроизрастания (ТУМ), устанавливают по растениям-индикаторам эдафических условий. На примере южной тайги все типы леса с преобладающей древесной породой — елью объединены в пять групп (эдафофитоценотический ряд): 1) ельники-зеленомошники; 2) ельники-долгомошники; 3) ельники сфагновые; 4) ельники травяные; 5) ельники сложные. Каждая из этих групп — один тип условий местопроизрастания. Для первой группы типов леса характерны хорошо дренированные более или менее богатые различного типа почвы, приуроченные к рекам; второй тип лесов располагается на тех же почвах, но хуже дренированных, с элементами заболачивания, вдали от рек. Для третьей группы явно выражено заболачивание на равнине, вне предела влияния рек. По поймам рек, ручьев с избыточным, но проточным увлажнением расположены типы леса четвертой группы. Леса пятой группы привязаны к самым богатым в зоне почвам с максимально высоким pH среды.
Украинские ученые (Е.В. Алексеев, П.С. Погребняк) построили классификацию типов условий местопроизрастаний леса (эдафическую сетку лесов) на основе увлажнения и трофности почв. В сетке трофотопы — это члены ряда, отличающиеся по содержанию в почве количества питательных веществ (по плодородию почв). П. С. Погребняк выделил четырехступенчатый ряд, начиная от крайне бедных А (боры) до наиболее плодородных почв Д (дубравы). По степени увлажнения ряд состоит из 6 ступеней — гигротопов: от крайне сухих — 0; через сухие — 1; свежие — 2; влажные — 3; сырые — 4 до лесных болот — 5. Считается, что для разных лесорастительных подзон наиболее благоприятными гигротопами являются свежие условия местопроизрастания. Комбинация трофотопа с гигротопом понимается как эдатоп. Здесь же надо отметить, что кроме вышеназванных классификаций типов условий местопроизрастания в лесной типологии существуют и другие направления, учитывающие как своеобразие лесных насаждений, так и сами принципы построения классификационных схем (математическая типологизация, генетико-экологическая концепция), однако все классификации типов условий местопроизрастаний строятся на биогеоценотической основе или по показателям химического богатства и влажности почвы.
В пределах одного и того же типа условий местопроизрастания может быть несколько типов леса в зависимости от древесных пород, их образующих. Например, в кисличниковом типе лесорастительных условий могут формироваться сосняки, ельники, пихтарники, березняки, осинники.
Тип лесорастительных условий — классификационная единица территории, которой пользуются лесоводы наряду с типами леса. Совокупность типов лесорастительных условий, в которых естественно распространен тот или иной вид древесного растения, составляет его эдафофитоценотический ареал.
Кроме основной систематической единицы растительного покрова — ассоциации, в лесной геоботанике принято выделять единицы более высокого ранга. Их иерархия (в порядке возрастания) такова: лесная ассоциация — группа ассоциаций — лесная формация — группа формаций — класс растительности — тип растительности.
В группу ассоциаций объединяют ассоциации, отличающиеся по составу только одного из ярусов. Например, древостой (совокупность деревьев, являющихся основным компонентом насаждения) образован елью европейской. Кустарниковый ярус отсутствует, моховой ярус хорошо развит и повсюду представлен зелеными мхами одного и того же состава. Ho травянисто-кустарничковый ярус различен: в одних фитоценозах — из черники (ассоциация ельник черничный), в других — из брусники (ельник брусничный), в третьих — из кислицы (ельник кисличный), а в четвертых — из зеленых мхов (ельник зеленомошный). Все эти четыре ассоциации составляют группу ассоциаций — ельник зеленомошный. Аналогичным образом объединяют в группы и другие лесные ассоциации.
Группы ассоциации объединяют в формации на основе общего эдификатора (или общих эдификаторов). Так. различают формации пихтовые (образованы разными видами пихт), еловые, сосновые, кедровые, лиственничные, березовые, осиновые, дубовые и т.д.
К группе формаций относят формации с одной и той же жизненной формой эдификаторов. Так, пихтовые, еловые, кедровые (сосна сибирская кедровая) формации составляют группу темнохвойных формаций; сосновые и лиственничные — группу светло-хвойных формаций; березовые, осиновые, тополево-ивовые и ольховые — группу мелколиственных формаций; дубовые, буковые, ореховые, ясеневые, липовые, кленовые, ильмовые, платановые и др. — группу широколиственных формаций.
Еще более крупной систематической единицей является класс формации, объединяющий все группы формаций, у которых эдификаторы близки по своим жизненным формам. Так, группы темно- и светлохвойных формаций составляют класс формаций хвойных лесов, а группы мелко- и широколиственных формаций образуют класс формаций лиственных лесов.
Совокупность классов формаций составляет тип растительности — самая крупная таксономическая единица в геоботанике (рис. 1).
По А.П. Шенникову, к одному типу растительности относят все формации, ассоциации которых в господствующем ярусе сложены одной и той же биоморфой. Различают типы растительности с господством деревьев, кустарников, трав, мхов и т.д. Лес как гип растительности представляет собой совокупность ассоциаций, в господствующем ярусе которого эдификаторами являются деревья. Эдификаторами типа растительности кустарниковые заросли служат кустарники и кустарниковые деревья; эдификаторы луговой, степной, травяно-болотной растительности — цветковые травянистые растения; эдификаторы ассоциаций моховых болот — сфагновые мхи.
В зависимости от географического положения природной среды лесные массивы России образованы различными древесными породами: на севере страны доминантами выступают ель, лиственница и сосна; южнее появляется пихта; хвойно-таежная зона переходит в зону смешанных лесов, далее идут лесостепь и степь. Особый тип лесной растительности формируется по поймам рек.
Темнохвойные лесные формации на западе образованы елью обыкновенной (европейской), к востоку ее сменяет ель сибирская. Область распространения пихты сибирской — северо-восток европейской части России, Западная Сибирь. К этой же группе относят сосну сибирскую кедровую, ареал которой во многом совпадает с ареалом пихты сибирской.
Светлохвойные формации сформированы сосной обыкновенной и видами лиственницы — сибирской, Сукачева и Гмелина (даурской). В европейской части страны распространены преимущественно сосновые леса, в Западной Сибири — сосновые и лиственничные, в Восточной Сибири — лиственничные.
Мелколиственные формации лесов характерны для всей лесной зоны. Образователи мелколиственных лесов — виды березы, тополь, осина, ольха черная.
Широколиственные леса образованы дубом черешчатым, липой метколистной, кленом остролистным, вязами — гладким и голым, ясенем обыкновенным.
В растительном покрове хвойно-широколиственных (смешанных) лесов преобладают южнотаежные ельники и сосняки в сочетании с чистыми дубравами и смешанными лиственными породами из видов вяза, ясеня обыкновенного, липы сердцевидной. На западе зоны в образовании этой группы лесов участвуют более теплолюбивые лиственные породы: бук лесной, граб обыкновенный, дуб скальный, вяз граболистный, клен ложноплатановый.
Деревья второго яруса в лесных фитоценозах хвойно-широколиственных лесов представлены рябиной обыкновенной, видами черемухи, кленами — полевым и татарским, боярышниками, яблоней лесной, грушей обыкновенной, ивой козьей, ольхой серой. Деревья этого яруса встречаются только на опушках.
В лесных фитоценозах в третьем ярусе, как правило, возникает подлесок, сформированный из различных видов кустарников и подроста главных пород. В хвойных и хвойно-широколиственных лесах лесной зоны подлесок образован ивами серой, чернеющей, козьей, ушастой; рябиной, ольхой серой, березой кустарниковой, крушиной ломкой. В различных типах сосняков в подлеске произрастают можжевельник обыкновенный, ива ушастая, береза карликовая, рябина, ракитник, дрок красильный. По эдафическим рядам в хвойно-широколиственных и широколиственных лесах прослеживается ряд от кисличников к травяно-дубравным и травяно-болотным типам леса, в подлеске которых широко распространены розы коричная, собачья, колючейшая; виды спирей, рябинник, боярышник сибирский, черемуха; крушина ломкая, жостер слабительный, жимолости татарская и синяя; калина красная: бузина черная и красная; лещина обыкновенная, бересклет бородавчатый и европейский; смородины черная, обыкновенная, альпийская, дерен красный, липа сердцевидная, вишня степная, слива колючая.
Степная зона также имеет древесно-кустарниковую растительность, представленную степными кустарниковыми зарослями, прибрежными лесами, полевыми защитными полосами, садами и парками населенных мест, сыртовыми лесами и овражно-балочными типами древесной растительности.
По оценке П.А. Богданова, в кустарниковой степи насчитывается до 25 видов кустарников, представителей девяти семейств сосновых и цветковых растений.
Древесно-кустарниковая растительность речных пойм интразональна. Своеобразие речных пойм связано как с условиями увлажнения, так и с многообразием типов почв.
Древесные и кустарниковые породы пойменных лесов по экологическим свойствам, морфологии существенно отличаются от пород коренных лесов природных зон. Первый ярус создают тополя черный, белый, лавролистный, душистый, темнолистный, корейский; древовидные ивы — ломкая, русская, белая; ольха черная. Во втором ярусе сосредоточены крупные и мелкие кустарники: боярышник, черемуха, рябина, шиповник коричный, виды смородины, бузина красная, жимолость татарская, чилига степная, вишня степная, ивы — русская, трехтычинковая, пурпурная, прутовидная, остролистная. Сомкнутость крон очень значительная (0,8—1,0). Травяной ярус выражен слабо или полностью отсутствует.
Травянистая растительность (растительность наземного яруса) формируется как зональный тип. В лесотундре на вечной мерзлоте и с высокой степенью заболачивания почвы широко распространены сфагновые болота, на которых основные компоненты — белые (сфагновые) мхи и лишайники (олений мох), из магнолиецветных — пушица, полярный мак, купальница, лютики, мытники.
В лесной зоне видовой состав травянистой растительности довольно богатый и кроме лишайников и мхов много видов осоки. Мятликовые представлены щучкой дернистой, белоусом, душистым колоском. Из бобовых обычны виды клевера, вики, чины.
От общей биологической массы лесного фитоценоза на долю травяного покрова приходится не более 1 % и за вегетационный сезон, например, в дубравах накапливается менее 10 % массы органического вещества, образуемого лесными фитоценозами. Тем не менее травяному покрову принадлежит важнейшая роль в общем круговороте веществ, так как травы, как правило, более высокозольные по сравнению с древесными видами. Поданным B.C. Ипатова, в 300-летней дубраве на долю травяного покрова приходится 1/3 азота и фосфора и около 2/3 калия, ежегодно поступающих в почву с растительными остатками. Этот факт подчеркивает важнейшую роль травяного покрова в общем биологическом круговороте лесных фитоценозов.
В сосновых лесах напочвенный покров более богат как по видовому составу, так и по образуемой за вегетационный сезон биомассе, что связано с достаточно большим количеством света, проникающего к напочвенному покрову. С этим явлением связана и малая амплитуда температур в кроне и у поверхности почвы. Считается установленным факт, что в сосновом лесу древесный полог задерживает до 325 мм годовых осадков. Запас доступной влаги в сосняках закономерно снижается от весны к осени.
Нижний ярус соснового леса представлен хорошо развитым травяным покровом из лесных, луговых, опушечных и сорных видов. Это купырь лесной, виды вероники, герани, полевицы полыней, тысячелистники, ястребинка зонтичная, tohkohoг стройный, пырей ползучий, чистотел майский, латук, подмаренники. Зеленые мхи представлены видами Pleurozium schreberi Mitt, Dicranium polysetum Mich, лишайники — родом Cladonia.
Фитоценологические понятия о типе леса и типе лесорастительных условий. Представления о задачах лесной типологии
Новые «типы леса» фитоценологов являются лишь весьма робким шагом в сторону лесоводственной типологии, ибо «в флористическом составе могут быть некоторые различия», т. е. различия небольшие. В. Н. Сукачев (1939) поясняет это на четырех примерах.
1-й пример. Можно объединить в один тип леса ельники-черничники с Picea excelsa и Р. obovata, так как это экологически сходные породы. Однако в «Дендрологии» В. Н. Сукачев подчеркивает неизученность экологии сибирской ели, а это противоречит утверждению об экологическом сходстве упомянутых видов ели и не дает права на их объединение.
2-й пример. Аналогичный случай с лиственницами сибирской и даурской, которые также разрешается объединять. С этим столь же трудно согласиться, ибо данные виды резко отличаются по экологии.
3-й пример. Во втором ярусе дубрав в одном случае господствует груша, в другом — яблоня при большом сходстве прочих ярусов. «Ассоциации здесь различные, но тип будет один». Такое объяснение с лесоводственной точки зрения неприемлемо, так как «при большом сходстве прочих ярусов» груша — великолепный индикатор более сухих, а иногда, вероятно, и слегка засоленных почв, в то время как яблоня — представитель более, выщелоченных и оподзоленных почв. Столь формальное отношение к древесным породам-индикаторам местообитания тем более нелогично, что, по мнению В. Н. Сукачева, дубравный покров является ненадежным показателем типов леса.
4-й пример. Дубравы на одинаковых почвах и при одном и том же видовом составе растительности могут быть семенными и порослевыми. Флористически — это одна и та же ассоциация. Но так как «борьба за существование» и отношения к среде у них разные, то это — различные типы леса. В данном случае лесоводы получают, наконец, «счастливую возможность» различать семенные и порослевые дубняки, хотя это достижение добыто ценой не объединения, как обещал В. Н. Сукачев, а, наоборот, еще большего дробления типов леса. Здесь перед нами полная потеря всякой ориентировки, ибо семенные и порослевые насаждения лесоводы не только хорошо различают, но и весьма обстоятельно изучили, а для дальнейшего изучения им ничем не поможет выделение семенных и порослевых «типов леса».
Регламентируя третье свое понятие — тип лесорастительных условий,. В. Н. Сукачев упрекает автора этих строк в том, что он «отождествляет тип; леса с типом лесорастительных условий». Оказывается, этого нельзя делать потому, что «могут существовать так называемые производные ценозы (березняки и осинники на месте хвойных пород и пр.), т. е. при одних и тех же лесорастительных условиях в данный момент могут расти разные древесные породы и реализоваться разные типы». Кроме того, «ряд древесных пород еще не достиг своих естественных ареалов» и возможны одинаковые местообитании при разном составе коренных типов леса.
Насколько нам известно, наличие коренных и производных насаждений впервые установил Гуторович, и это нисколько не помешало ему заимствовать народные представления о единстве насаждения и местообитания, равным образом как и логически вытекающую из наличия этого единства тенденций относить насаждения березы на песчаных почвах к борам. Факты незаконченного расселения растительности совершенно не противоречат единству насаждения с местообитанием, если только понимать единство организмов и среды не как мертвое статическое единство, а как текучее, меняющееся в процессе взаимодействия организмов и условий существования. Вообще в самом ходе мыслей фитоценологов, устанавливающих, что существование в однотипных условиях местообитания разных коренных и производных насаждений не дает возможности оценивать местообитание по составу растительности, есть существенный дефект, так как методом исследования признается лишь анализ (расчленение целого на все более и более дробные части) и исключается синтез.
Здесь же В. Н. Сукачев кратко излагает особенности лесоводственной классификации типов леса, как он ее себе представляет (она называется «взгляды П. С. Погребняка»). «П. С. Погребняк сводит почвенные свойства, влияющие на лес, к основным двум признакам — к влажности местообитания и к химическому богатству почв». Тут же указано, что в нашей классификации получается «24 сочетания…, с которыми лесоводу приходится считаться в своей деятельности». Цифра «24» в дальнейшем упоминается еще несколько раз. Только после ознакомления с данной статьей В. Н. Сукачева цифра «24» представилась нам впервые как свойство лесоводственной классификации, ибо до этих пор мы не удосужились поинтересоваться счетом клеточек на схеме, иллюстрирующей принципы классификации. Читатель уже знаком из предыдущего с лесоводственной классификацией и знает, что число «сочетаний» в ней может сильно колебаться, примерно до 120 в пределах эдафических ординат, а климатические формы, если отнести классификацию к географическим пределам европейской части СССР, вносят свой множитель, увеличивающий число типов леса в два-три раза по отношению к числу эдафических вариантов. Поэтому упрек в слишком малом количестве единиц нашей классификации настолько же справедлив, насколько было бы справедливым обвинение в чрезмерном их обилии.
Но не в этом, конечно, заключаются оригинальные черты лесоводственной классификации типов леса и не к этой «количественной» стороне свозится ее производственное значение. На практике получается так, что типологи лесоводы выделяют в пределах лесничества всего 4—5, редко 8—10 или более типов леса; фитоценологи выделяют обычно несколько десятков типов леса. Но, как это ни странно на первый взгляд, лесоводственная классификация гораздо детальнее фитоценологической. Дело в том, что лесоводственная классификация, твердо придерживаясь своего гибкого классификационного принципа (сходство и различие типов леса по 1) почвенному плодородию, 2) увлажнению и 3) климату), фиксирует внимание на более существенных признаках, шире и полнее охватывает амплитуду естественного разнообразия лесов по этим существенным признакам. Фитоценологи же, выделяя «типы леса» на основе внешних признаков состава растительности и ее обилия, конкурируют с таксационным описанном, и их выделы не только дробят на части одни и те же тины леса (например, выделяют особо сосняки и особо березняки одного и того же свежего бора), по и оста пли ют без законного и необходимого разделения участки разных типов леса, если они имеют одну и ту же господствующую породу при небольшой разнице в ее бонитетах (например, березняки свежего бора и свежей субоои, имеющие зачастую одинаковый состав доминантой травяного покрова). Если в конкретных случаях у них получается больше типов леса, то это происходит вовсе не в результате более точной работы.
Практическую цель типологии В. Н. Сукачев формулирует следующим образом: «Типология леса, с одной стороны, должна помочь нам расчленить естественные лесные массивы на единицы, однородные по производительности и требующие одинаковых хозяйственных мероприятий, отвечающих природе этих объединений; с другой стороны, она должна дать указания, как создать новые, более полезные, более производительные типы леса, по своей продукции превосходящие природные». В первой части этого высказывания перед типологией поставлена весьма простая и ограниченная задача формального деления целого на части по признакам однородности выделяемых пространственных единиц без раскрытия их содержания. Во второй части, наоборот, перед типологией поставлена непосильная задача, относящаяся к компетенции лесокультурников, специалистов по рубкам ухода, селекционеров и других специалистов. В ее решении типологи могут принять участие лишь в случае достаточной их компетентности в области лесоводственных наук, чего, как видно из приведенных выше примеров, у многих типологов недостает.
Подчеркивая свою мысль о том, что интерес лесоводов к типологии связан прежде всего с интересом к древесным породам, фитоценологи, видимо, и не подозревают, что лесоводы нуждаются как раз в противоположном, что интересы лесоводов куда глубже и сложнее, чем фитоценологическое представление о задачах типологии. Лесоводы стремятся увидеть за деревьями лес, т. е. отношение деревьев к среде, к местообитанию, уяснить причины разнообразия лесов. Фитоценологи же настойчиво хотят оставить лесоводов наблюдателями внешней стороны леса, ограничиваемой господствующими породами и названиями фитоценозов. При этом они рисуют далекую перспективу внедрения типологии в производство после того, когда типы леса будут изучены путем длительной и полной индукции.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
5 Тематика лабораторных занятий по
%PDF-1.5 % 1 0 obj > /Metadata 2 0 R /Outlines 3 0 R /PageLayout /OneColumn /Pages 4 0 R /StructTreeRoot 5 0 R /Type /Catalog >> endobj 6 0 obj /CreationDate (D:20160628170003+03’00’) /Creator /Keywords () /ModDate (D:20160628170018+03’00’) /Producer (Adobe PDF Library 10.0) /SourceModified (D:20160628135953) /Subject () /Title >> endobj 2 0 obj > stream 2016-06-28T17:00:18+03:002016-06-28T17:00:03+03:002016-06-28T17:00:18+03:00Acrobat PDFMaker 10.1 для Worduuid:18c76967-497b-48e8-9706-26ebc8c0474euuid:17904841-e7a3-40a5-90ba-b4d768087580
Леса, их типы
За основу объединения типов леса в группы, следуя Колесникову, берется качественное сходство их типов условий местопроизрастания по ведущим факторам (трофность, влажность), что определяет однородность экологического облика растительности. В зависимости от экономических условий ведения лесного хозяйства группа типов леса может быть основной единицей, для которой необходимо планировать систему хозяйственных мероприятий (способ главной рубки, вид и метод рубок ухода, меры по лесовосстановлению и т. д.).[ …]
Тип леса (коренной тип леса, тип лесорастительных условий, за исключением нелесных земель, тип местопроизрастания, серия типов леса и т.д.) — совокупность сходных коренных лесных биогеоценозов (экосистем), включающих все стадии их восстановления после разных нарушений.[ …]
Влияние лесов на почву в СССР сказалось прежде всего в том, что при их участии образовались определенные типы почв — подзолистые, лесные суглинки и деградированные черноземы.[ …]
Учение о типах леса как раздел науки получило наибольшее развитие в нашей стране. Под типом леса следует понимать совокупность участков с древостоями или насаждениями, объединенных однородными лесорастительными условиями, более или менее общим составом и прочими общими компонентами лесного сообщества, единством происхождения, сходной историей развития, общим лесоводственным характером. При сходных экономических условиях эти участки требуют одинакового хозяйственного на них воздействия и позволяют осуществить однородное их использование.[ …]
Описание типов леса в статике на первых порах было вполне правомерным; оно оправдывалось неизученностью и новизной вопроса, преобладанием девственных или мало тронутых человекам лесов, ограниченными потребностями практики, всецело заинтересованной в характеристике спелых лесов, их инвентаризации для целей эксплуатации. Конечно, лесная типология давно стремится служить и целям лесовозобновления, преимущественно через выделение типов условий местопроизрастания.[ …]
Во влажных типах леса, например, условия среды для возобновления леса более благоприятны на бровках (валиках) волоков и неблагоприятны на донной части их, которая нередко заливается водой; наоборот, в типах леса с легкими дренированными почвами дно волока мажет представлять наиболее благоприятную среду, за исключением случаев, когда под влиянием многократного прохождения трактора почва может уплотниться. На условиях наземной и почвенной среды сказывается сезон заготовки, так как с ним связан характер воздействия трелевки. Большое влияние на лесорастительные условия вырубок оказывает огонь.[ …]
В развитии типа леса, как указывает И. С. Мелехов, можно выделить этапы, которые предшествуют образованию леса (типы вырубок, гарей), этапы формирующегося типа леса, этап сложившегося типа леса, возможные последующие этапы с переходом или без перехода в новый тип леса [10, 11, 14]. Системный анализ этапов и их границ в формировании типов леса во времени имеет большое практическое значение для раскрытия путей и возможностей рационального ведения лесного хозяйства [12, 14, 18].[ …]
После рубки леса происходят большие изменения в условиях среды: меняется световой и температурный режим, напочвенные н почвенные условия. Тенелюбивые растения, росшие в лесу, отмирают, а на их месте разрастаются светолюбивые. Они в значительной мере и определяют новую среду и тип вырубки.[ …]
В отличие от лесов умеренной зоны, где жизнь приурочена к надпочвенной зоне, в дождевом лесу значительно большая доля животных обитает в верхних ярусах растительности. Так, из 59 видов млекопитающих Гайаны 31 вид — древесные животные, 5 — амфибионты » только 23 живут в надпочвенном слое (Хевиленд, 1927). Кроме древесных млекопитающих, здесь встречаются в изобилии хамелеоны, игуаны, гекконы, древесные змеи, лягушки и птицы. Важную экологическую роль играют муравьи и прямокрылые, а также дневные бабочки и моли. Флора и фауна дождевого леса безгранично богаты видами. Так, на 15 км2 района Барро-Колорадо, хорошо изученного участка дождевого леса в зоне Панамского канала, насчитывается 20 ООО видов насекомых, тогда как на территории всей Франции их всего несколько сот. Разнообразные архаичные типы животных и растений выжили в многочисленных нишах неизменяющейся среды. Многие ученые полагают, что для тропического леса характерна высокая скорость эволюционного изменения и видообразования; вполне возможно поэтому, что эти районы послужили местом возникновения многих видов, вошедших в состав более северных сообществ.[ …]
Отношения по типу 2 могут быть причиной относительной стабильности популяции некоторых видов, в частности, они распространяются иа грибы под пологом леса. Их можно распространить и на некоторых животных из числа тех, кто питается падалью, а не нападает непосредственно на свою жертву. Для животных это отношение может быть не столь простым, если питающаяся популяция некоторым образом влияет на популяцию-пищу, но при этом не уменьшает ее способность к размножению и самосохранению. Конкуренция в этом случае возникает между особями питающейся популяции, которые используют только «процент», или излишки, не влияющие на поддержание устойчивости кормящего вида.[ …]
Для изучения типов леса, возрастных и восстановительных этапов их развития закладываются экологические профили, к которым привязывают постоянные пробные площади. На этих стационарных научных объектах проводят лесоводственные, таксационные, геоботаниче-ские и зоологические исследования.[ …]
Периодичность их связана и с чередованием сухих и влажных лет ), и с сезонными и суточными изменениями погоды, и с характером леса (в частности, с типом леса) и его изменениями во времени (фенологическими и фито-ценотическими), и с характером предшествовавших пожаров, и с территориальным местонахождением лесных массивов и отдельных частей их, и4 с деятельностью человека. Таким образом, периодичность в горимости лесов может быть правильно понята, а отсюда и сделаны практические выводы лишь при учете ряда моментов и их возможных сочетаний.[ …]
Распределение лесов земного шара по составу пород тесно связано с типами климата. Так, во влажных тропических районах произрастают преимущественно лиственные леса, отличающиеся чрезвычайным разнообразием составляющих их древесных пород; в холодных и умеренно холодных районах преобладают хвойные леса, состоящие из сравнительно небольшого числа древесных пород. Исторически развиваясь в условиях более или менее определенного климата, древесные породы оказываются приспособленными к соответствующим климатическим условиям. Известно, что один и тот же ботанический вид древесной породы, произрастающий в различных климатических условиях, может дать различные климатические экотипы (расы), приспособленные к существованию в условиях соответствующей разности климатического типа.[ …]
Возобновление леса и напочвенные изменения, а также изменения в почве взаимообусловлены. Смены растительного покрова и формирование леса через последующее возобновление его можно представить на примере некоторых типов леса в виде следующей схемы (рис. 40). Число таких схем, естественно, может быть больше, но в этом нет необходимости, так как схема отражает принципиальную суть мет,одического подхода к раскрытию закономерностей в изменении растительных этапов на концентрированных вырубках. Различия в растительных этапах, их взаимосмены сказываются и на продолжительности возобновления леса, которое может быть весьма различным. Сокращение периода возобновления хозяйственно-ценных пород — одно из средств повышения продуктивности лесов. Последующее, тем более нерастянутое, возобновление леса дает возможность получить в будущем древесину высокого качества — с рав-номерньпм строением годичных слоев, очищением стволов от сучьев и т. д.[ …]
Все компоненты леса синтезируются в понятии „тип леса“. Важную задачу поэтому представляет выяснение опасности возникновения пожаров и их поведения в связи с различиями в типах леса.[ …]
К определенным типам леса приурочен не один только какой-нибудь вид животных, а целая группа их, связанная между собой процессом борьбы за существование в однородных экологических условиях. Дарвин вслед за Ньюменом установил, что изобилие некоторых цветковых растений, например красного клевера, зависит от большего или меньшего присутствия животных кошачьей породы, истребляющих мышей: мыши разоряют гнезда и соты шмелей, а шмели способствуют перекрестному опылению красного клевера.[ …]
Многие коренные типы леса бывают более производительными, чем их производные. Поэтому хозяйственная деятельность должна быть направлена на восстановление коренных типов леса, за небольшим исключением в курортных лесах, где на первое место выдвигаются другие требования. С хозяйственной точки зрения объединять типы леса в серию, принимая ее за хозсекцию (как предлагают некоторые био-геоценологи), ни в коем случае недопустимо, так как типы леса из одной серии в горных условиях произрастают в различных лесорастительных условиях.[ …]
Выборочные рубки в лесах разных типов необходимо анализировать по результатам их влияния на оставшийся древостой, на возобновление леса и формирование его.[ …]
Формирование нового типа леса на месте старого может идти разными путями. Вот почему при изучении типов леса, их использовании в практике лесного хозяйства все настоятельнее проявляется необходимость учета происхождения типа леса, значения этапов его формирования и происшедших смен, определения тенденций дальнейшего развития, понимания того, что будет в дальнейшем при естественном ходе событий и что может быть при различных формах вмешательства человека, в том числе — в смысле возможностей превращения существующего малопродуктивного типа леса в высокопродуктивный. В решении задачи создания высокопродуктивных и наиболее ценных лесов будущего, удовлетворяющих многосторонние запросы и потребности людей, динамическая типология леса может оказать большую пользу.[ …]
Образ жизни. Населяют леса различных типов, преимущественно смешанные и лиственные, осветленные, с прогалинами и полянами, любят пойменные леса. Насиживание начинается после завершения кладки и длится 14—15 (до 17) дней. Самец обычно сидит ночью, самка — днем. У гнезда осторожны, с начала насиживания и часто до вылета птенцов взрослые почти не подают голоса. Птенцы вылетают из гнезда в возрасте 24—28 дней. За 2—3 дня до вылета практически весь день кто-то из птенцов высовывается из дупла и подает голос. Питаются в бесснежное и малоснежное время больше всего муравьями, нередко собирают насекомых на земле. Зимой больше копаются в гнилушках, обшаривают древесные стволы. Долбят относительно немного. Взрослые оседлы, молодые в конце лета и осенью активно расселяются. Зимой могут кочевать и взрослые. Чаще других дятлов их можно видеть в городах и деревнях лазающими по деревянным домам, осматривающими щели в кирпичных кладках. Максимальный известный возраст — более 5 лет.[ …]
Вопросы возобновления леса в связи с типами вырубок уже освещались в печати. Часть их рассмотрена и в некоторых статьях настоящего сборника.[ …]
Зона обитания корней в лесу заселена многочисленными микроорганизмами — бактериями, актиномицетами, грибами, протозойными и другими формами мельчайших существ, Деятельность их оказывает серьезнейшее влияние на плодородие .почвы, а значит, и на рост и жизнедеятельность леса. Выяснение изменений, происходящих в составе и жизнедеятельности микроорганизмов на лесосеках, после рубки, представляет большой интерес для типологии вырубок. С одной стороны, состав и жизнедеятельность их должны различаться в разных типах вырубок, с другой, своей деятельностью они должны оказывать какое-то влияние на формирование типа вырубки, и самое существенное при этом то, что они во многом определяют среду для начальных этапов возобновления леса.[ …]
Как уже отмечалось выше, тип вырубки мы рассматриваем не оторванно от типа леса, от географической среды, от фактора времени. Тип вырубки с учетом этих моментов, прежде всего типа леса, даёт ключ крещению важнейших вопросов естественного и искусственного лесовозобновления на сплошных вырубках; он отражает лесорастительные условия и через них возможности и пути возобновления леса не только в данный момент, что, разумеется, очень важно и необходимо, но и позволяет судить и о прошлых и будущих изменениях лесорастительных условий и, следовательно, дает возможность прогноза возобновления леса. Степень достоверности прогнозов зависит от изученности типов вырубок, особенно их динамики, от раскрытия закономерности в возобновлении леса в связи с типами вырубок, в процессах заболачивания и разболачивания, задернения и раздернения и т. д.[ …]
Пьявченко Н. И. Некоторые типы болот и заболачивание лесов Тюменской области. Труды ии-та леса АН СССР, т. XXV!, 1955. Сукачев В. Н. Болота, их образование, развитие и свойства. Цинзерлинг Ю. Д. География растительного покрова Северо-Запада Европейской части СССР. Труды геоморфологического ии-та, вып. 4, Л., 1932.[ …]
В разнотравно-ситниковом типе вырубки по сравнению с другими происходят наименьшие изменения в живом напочвенном покрове. Динамика растительного покрова здесь зависит от характера повреждения почвы и сомкнутости сохраненного во время рубки подлеска и подроста. Участки вырубок с минерализованной поверхностью почвы интенсивно зарастают ситником, проективное покрытие которого на 3-й год после рубки достигает 50 %. На 2—3-й год в покрове появляется вейник тростниковидный (проективное покрытие 2—5 %). На волоках мощность опада трав, в основном ситника, на 3-й год достигает 10 см. На минерализованной поверхности происходит сильное задернение. В отдельных местах появляется ольха серая. На той части вырубок с неповрежденной поверхностью почвы, которая занята подлеском и подростом предварительного возобновления, в живом напочвенном покрове наблюдаются меньшие изменения. Кустарники и молодняки древесных пород, сохраненные при лесоразработке, сдерживают разрастание живого напочвенного покрова, о чем свидетельствует наличие значительной корреляции (г =0,56 ± 0,061; !г =9,18) между их проективными покрытиями. На остальной неповрежденной поверхности почвы (не занятой подростом и подлеском) в покрове преобладает вейник. В последующие 3 года проективное покрытие живого напочвенного покрова в целом на вырубках этого типа стабилизируется. Однако в его составе происходят изменения. Начинает постепенно деградировать ситник и заметно возрастает доля участия вейника. Сомкнутость подлеска возрастает до 0,3—0,4. На 7—8-й год после рубки под влиянием естественного возобновления леса проективное покрытие живого напочвенного покрова снижается. На минерализованной части почвы доля участия ситника уменьшается, а вейника возрастает. На остальной части вырубки вейник постепенно деградирует.[ …]
Сосняки кисличной группы типов леса благодаря сочетанию благоприятных условий влажности, более высокой обеспеченности элементами питания и аэрации почвы являются наиболее высокопроизводительными фитоценозами района исследований. Их можно встретить на выравненных участках и верхних половинах склонов моренных холмов, террас и в поймах рек на песчаных или супесчаных дерновослабоподзолистых почвах. Древостой представлен значительным разнообразием древесных пород. Кроме сосны, в составе возможно участие лиственницы, березы, ели, ольхи и осины.[ …]
Выделенный нами лещиновый тип вырубки характеризуется сильным разрастанием лещины, которая нередко образует даже сплошные заросли [16]. В западной части зоны смешанных лесов к лещине подмешиваются клен и липа. Лещиновые вырубки формируются из-под группы сложных типов леса там, где под пологом произрастает лещина. Наличие последней под пологом древостоев и создает потенциальные возможности для формирования после их рубки лещинового типа вырубок.[ …]
Субформации лиственничных лесов характеризуются не только величиной ареала каждого вида лиственницы, но и комплексом экологических и топографических условий, обусловливающих их внутреннюю структуру. Способность лиственницы произрастать на самых различных элементах рельефа и формировать типы леса со своим преобладанием требует объединения этих условий в более крупные однородные единицы — геоморфологические комплексы лиственничных лесов. Геоморфологические комплексы являются таксономической единицей как лесорастительных условий, так и одновременно классификационной единицей типов лиственничных лесов, соответственно равных по рангу и объему. В составе геоморфологического комплекса объединяются типы лиственничных лесов, произрастающие на крупных категориях форм рельефа и характеризующиеся однородностью процессов их формирования. В целом для формации лиственничных лесов Сихотэ-Алиня выделяется 5 геоморфологических комплексов типов леса — высокогорные, горных склонов, горных плато, долин и лиственничных лесов на приморских песках. Последние хотя и занимают ничтожные площади в северной и средней части Сихотэ-Алиня, располагаясь узкой полосой по песчаным дюнам вдоль морского побережья, однако их своеобразные местообитания, связанные с постоянной морской засоленностью цочвы и воздействием морских ветров, и особенности строения позволяют выделить такие насаждения в особый геоморфологический комплекс лиственничных лесов. Весьма характерным является и тот факт, что лиственница в подобных условиях формирует коренные типы леса.[ …]
Типология вырубок и типология леса связаны между собою не только тем, что определенный тип вырубки или группа их происходит от определенных типов леса. Имеется и другая сторона, включающая вопросы дальнейшей смены типов зырубки «лесными этапами» и формирования на месте определенных типов вырубок соответствующих типов леса. Говоря в этом смысле о возникновении, развитии и формировании леса на сплошных концентрированных вырубках, можно выделить несколько путей, из них наиболее характерных и резко отличных один от другого три: 1) облесение вырубки сразу после рубки за счет предшествующего, а также и за счет быстрого последующего возобновления леса; 2) смену одного определенного типа вырубки непосредственно лесом; 3) образование леса после смены нескольких типов вырубок.[ …]
Динамический подход к изучению типов леса также облегчает диагностику определения коренного или производного типа леса. Дело в том, что при внешних сходствах и одинаковых названиях тип леса в одних лесорастительных регионах является коренным, в других — кратковременно-производным, поэтому их нужно относить в разные хозяйственные секции. Например, коренной березняк-черничник из восточной части Месхет-Джавахетии или Внутреннего Дагестана в регионах Западного Закавказья обычно бывает производным, вторичным от одноименного ельника-черничника. Таких примеров можно привести множество.[ …]
Преобладающая часть древостоев II типа формирования во всех рассматриваемых типах леса относится к категории умеренно густых. Только в лишайниковых борах около 20 % их оцениваются как густые и примерно 15 % — как перегущенные.[ …]
Общие сведения о насекомых — вредителях леса. Строение, питание, образ жизни. Основные сведения о насекомых. Термины и определения. Экологические факторы и их роль в жизни насекомых. Полезные и вредные насекомые. Основные группы. Межвидовые и внутривидовые отношения, колебания численности насекомых, вспышки их массового размножения. Симптомы болезней растений. Группы и типы болезней, вызываемых насекомыми. Экономический и экологический ущерб.[ …]
Под лесной формацией понимается совокупность всех типов леса, объединенных общностью одной или нескольких лесообразующих пород с близкими биоэкологическими свойствами на всех стадиях возрастных, восстановительных и аллювиальных смен их насаждений. Формация обладает определенным ареалом, т. е. занимает физико-географическую область, обусловливающую территориальную однородность растительности.[ …]
Однако нельзя эту новую среду рассматривать оторванно от леса; она связана с характерам леса до рубки и временем, истекшим после нее. Важное значение при оценке условий среды концентрированных вырубок имеет учет бывшего типа леса; с ним во многом преемственно бывает связан и растительный покров вырубок и процессы возобновления леса. Но этого недостаточно. На вырубках создаются новые напочвенные и даже почвенные условия (особенно если иметь в виду верхние горизонты), причем нередко разные даже на месте одного и того же типа леса. Возникает необходимость учета не только типов леса, но и выделения связанных с ними естественно-исторических типов вырубок, отражающих их природные свойства, основные особенности их среды.[ …]
На границе подзон хвойно-широколиственных и широколиственных лесов доминируют не девственные коренные насаждения с выраженными признаками каждого типа леса, а молодняки и средневозрастные древостой, нередко производных типов леса, характеристики которых в этих возрастных группах требуется установить. Еще Г. Ф. Морозов, выделивший брянские леса по песчаной равнине от Жиздры до Полесья, считал первой задачей опытного дела — расчленение массива на типы насаждений с генетической характеристикой их на основании почвенно-геологических и ботанико-географических исследований [7].[ …]
Одной из причин не всегда успешного искусственного возобновления леса в таежных районах является то, что лесные культуры осуществляются нередко без учета и правильной оценки процессов естественного возобновления леса, без достаточного понимания особенностей природы концентрированных вырубок, их лесорастительных условий. Только при учете этих моментов, то есть на научной основе, могут быть разработаны рациональные типы лесных культур для различных районов таежной зоны.[ …]
По способам производства лесных культур, а также по агротехнике проведения их все типы вырубок, как и типы леса, можно разбить на вырубки без избытка влаги и с избыточным увлажнением.[ …]
Ряд отделов лесоводственной науки создан или целиком русскими учеными, или при их ведущем участии.[ …]
Основное внимание в этом пособии уделено краткому описанию простейших приемов посева и посадки леса, применению их в условиях, наиболее характерных для таежных районов европейской части СССР типов вырубок, некоторым вопросам использования лесозаготовительной техники для лесовосстановительных целей.[ …]
Хотя фитоценоз, по В. Н. Сукачеву, является главной составной частью биогеоценоза, тем не менее границы типа биогеоценоза определяются границами типа фитоценоза (ассоциации) лишь на определенном этапе развития. Если в пределах типа леса имеются участки в стадии жердняка, приспевающего, спелого или перестойного леса, они часто сильно отличаются по составу и сложению травяно-кустарничкового и мохово-лишайникового ярусов, а иногда и по составу подлеска. Возможны некоторые различия и в древостое. На таких участках разновозрастного леса будут также значительно отличаться и почвенно-грунтовые, и гидрологические, и микроклиматические (точнее, фитоклиматические) условия. В этом случае нужно устанавливать различные ассоциации, но относить к одному типу лесного биогеоценоза, рассматривая их лишь как его разновозрастные стадии [19, с. 46].[ …]
В южнотаежных районах наиболее острая необходимость в искусственном возобновлении хвойных ощущается на вырубках в сложных типах леса, например в ельнике липняковом, ввиду крайне затрудненного естественного возобновления хвойных. Здесь образуются разнотравные типы вырубок с мощным травяным покровом или вырубки, покрывающиеся обильной порослью лиственных пород. Применительно к этим вырубкам особенно необходимо не опоздать с проведением культур — проводить их в первый год после рубки. На задернелых вырубках надо переходить на посадки. В разнотравных типах южнотаежных районов возникает необходимость в более ранних и более частых уходах за культурами. В условиях более тяжелых влажных почв сеют и сажают в микроповышения (создаваемые оборачиванием пласта и т. п.).[ …]
Количество семенников также связано не только с породой и с характером деревьев в пределах породы, но и с условиями произрастания леса. Так, в типе леса, где опасность задернения почвы невелика, можно оставлять семенников меньше, чем в типе, где опасность задернения значительна. Если в вересковом сосняке достаточно оставить 10 семенников -на га, то в кисличнике может потребоваться 25—30 семенных деревьев. Количество семенников связано с наличием или отсутствием других видов обсемен ителей. Так, при сплошно-лесосечных рубках определенную роль играют стены леса. Около них семенникоЬ можно не оставлять, сосредоточивая их в середине лесосеки. Само собой разумеется, что при этом расчет количества оставляемых семенников необходимо увязывать с шириной лесосеки. Надо также учитывать наличие подроста и его распределение по площади. Возьмем, например, два участка: в одном равномерно размещен хвойный подрост, в другом— подроста нет. В первом случае можно оставить несколько страховых семенников на гектар, буквально «на всякий пожарный случай», а в другом — семенников оставляется больше—из расчета полного обсеменения ими щеего участка. Семенники могут оставляться одиночно и группами, равномерно и неравномерно распределенными по площади.[ …]
Необходимо отметить, что, несмотря на сильные изменения состава и структуры лесных сообществ в разных стадиях возрастного развития, диагнозирование их принадлежности к определенному типу леса, отличающемуся своеобразным лесообразовательным процессом, вполне возможно, но для этого необходимы специальные исследования не только фитоценотического состава, но и других компонентов биогеоценоза, охватывающие все стадии развития насаждений. В условиях Литвы такие исследования проведены в елово-лиственных лесах, но пока еще недостаточно изучены сосновые леса.[ …]
Исследования изменений таксационных элементов во времени, сопряженность между ними интенсивно продолжаются как в нашей стране, так и за рубежом. Однако лес как целое в совокупном виде, т. е. как тип леса, до начала XX в. трактовался лишь как категория пространственная. Типы насаждений описывались применительно к участкам леса с естественными спелыми сомкнутыми древостоями, соответствующими определенным лесорастительным условиям. Описание типов леса в статике на первых порах было правомерным. Оно оправдывалось неизученностью и новизной вопроса, преобладанием девственных лесов, ограниченными потребностями практики, заинтересованной в характеристике спелых лесов и их инвентаризации для эксплуатации. Эксплуатация леса осуществлялась преимущественно выборочными или другими способами рубки, сохраняющими основные черты прежнего леса. По мере дальнейших лесотипологических исследований постепенно стало проявляться внимание к природным изменениям леса во времени.[ …]
По нашим данным, в почве кипрейно-паловых вырубок сравнительно немного маслянокислых бактерий. Закономерности в развитии маслянокислых бактерий в исследуемых почвах нам установить не удалось, можно только отметить, что их количество как в почве леса, так и в почвах вырубок обоих типов выражалось числами одного порядка.[ …]
Однако нельзя сказать, чтобы она в достаточной мере отражала выделы, „однородные по горимости и требующие однородных противопожарных мероприятий“. В ней отсутствуют или недостаточно оттенены такие важные определяющие моменты, как тип леса, полнота древостоя, порода.[ …]
Искусственное лесовосстановление на лесокультурных площадях проводится посевом семян и посадкой сеянцев или саженцев, реже черенков. Посев и посадка не являются универсальными методами и имеют определённые достоинства и недостатки, а применение их обусловлено лесорастительной зоной, бывшим типом леса, типами условий местопроизрастания и типами вырубок, лесоводственными свойствами древесных и кустарниковых пород, целевым назначением культур.[ …]
С теоретической точки зрения каждая из вышеописанных стадий лесообразовательного процесса представляет своеобразный биогеоценоз, так как изменение фитоценогического состава сообществ сопровождается изменениями других компонентов биогеоценоза, а также функциональных связей между ними, что изменяет особенности процессов обмена веществ. Таким образом, следуя концепции В. Н. Сукачева [8], что тип леса — это тип лесного биогеоценоза, отдельные стадии развития которого можно было бы считать отдельными типами леса. Однако разделять единый лесообразовательный процесс, охватывающий период развития одного древостоя на ряд типов леса, нецелесообразно. С позиций генетического направления лесной типологии Колесникова, а также динамической типологии Мелехова, концепции которых мы считаем справедливыми как с теоретической, так и с практической точки зрения, все стадии развития насаждения относятся к одному типу леса. В таком случае необходимо определить диагностические признаки, способствующие достоверно точно установить принадлежность отдельных биогеоценозов, отдельных участков леса, находящихся в разных стадиях возрастного развития к определенным типам леса. Такие признаки есть, хотя они и не всегда наглядны. Как уже упоминалось выше, даже на вырубках, под покровом господствующих растений вырубок, сохраняются и лесные виды. Это наглядно показано в табл. 2, из которой видно, что многие лесные виды сохраняются во всех стадиях лесообразовательного процесса, хотя и не на всех участках. Однако их проективное покрытие уменьшается и они становятся менее заметными.[ …]
У практиков обычно вызывает сомнение оставление еловых кулис. Действительно, еловые деревья, группы и кулисы нередко вываливаются ветром. Но это бывает преимущественно на влажных и мокрых почвах, а также и на почвах менее влажных, но подстилаемых на небольшой глубине известковой плитой, карбонатным плотным суглинком или карбонатной глиной. Основным обсеменителем здесь может быть подрост ели. Но при дренированных, легких, глубоких почвах.[ …]
Какие существуют типы лесов и какое значение они имеют?
Проще говоря, лес — это большая группа деревьев. По данным Продовольственной и сельскохозяйственной организации Объединенных Наций (ФАО), лес — это земля с кроной деревьев более 10 процентов и площадью не менее 0,5 гектара.
Леса покрывают около 30 процентов поверхности земли и содержат около 3 триллионов деревьев. Они существуют в сухом, влажном, ледяном и жарком климате. Лесная экосистема — это сообщество организмов, живущих в лесу.
Существуют различные типы лесов, классифицируемые в основном в зависимости от их удаленности от экватора. И если мы поймем важность лесов, тогда мы узнаем, что безлесный мир может обернуться гибелью для всех форм жизни, включая людей.
В этом свете мы рассмотрим некоторые из основных преимуществ лесов. Однако перед этим давайте исследуем различные типы лесов.
Различные типы лесов
Существует три основных типа лесов — тропических, умеренных и северных .Они классифицируются по широте. Кроме того, эти основные типы делятся на более конкретные категории.
Давайте подробно рассмотрим основные типы лесов?
1. Тропические леса
Тропические леса находятся между 23,5 o северной широты и 23,5 o южной широты. Температура в этих лесах колеблется от 68 o до 77 o по Фаренгейту в течение всего года.
Тропические тропические леса могут похвастаться большим разнообразием видов всех экосистем на Земле.Они не переживают зимы и обычно получают 100 дюймов дождя в год.
Из-за высоких температур и влажного воздуха в этих лесах разложение происходит с невероятной скоростью. Высокий уровень осадков обычно приводит к вымыванию питательных веществ из почвы. Это объясняет, почему почвы в тропических лесах бедны питательными веществами.
В тропических лесах преобладают широколиственные деревья, высота которых составляет от 82 до 115 футов. Другая растительность включает виноградные лозы, папоротники, мхи, орхидеи и пальмы.
Солнце почти не достигает нижних уровней леса из-за густо растущих деревьев, образующих густой полог. Итак, большинство животных, населяющих тропические леса, приспособлены к жизни на деревьях. В этих лесах можно встретить самых разных птиц, змей, летучих мышей и обезьян.
Различные категории тропических лесов включают:
- Вечнозеленые: Вечнозеленые леса получают дожди круглый год и не имеют засушливого сезона
- Сезонность: У них вечнозеленая растительность и короткий сухой сезон
- Сухой: В этих лесах длинный сухой сезон, когда деревья теряют листву
- Монтане: Известны как облачные леса, поскольку они получают большую часть осадков из тумана или тумана, поднимающегося с низменностей.
- Тропические и субтропические хвойные: Эти леса имеют сухой и теплый климат с хвойными растениями, адаптированными к изменчивой погоде
- Субтропические: Субтропические леса расположены к северу и югу от тропических лесов.Деревья здесь приспособлены к летней засухе
2. Леса умеренного пояса
Леса умеренного пояса делятся на две подкатегории:
- Лиственные леса умеренного пояса
- Хвойные леса умеренного пояса
Лиственные леса умеренного пояса
Лиственные леса умеренного пояса можно найти в Китае, Японии, Западной Европе и некоторых частях России, а также на востоке США и в Канаде. Они переживают четыре разных сезона, и осадки выпадают в течение всего года.
Осадки бывают в виде дождя весной, летом и осенью. Зимой идет снег. В лиственных лесах умеренного пояса ежегодно выпадает от 30 до 60 дюймов дождя. И почвы там очень плодородные.
Вы должны ожидать найти папоротники, мхи и полевые цветы на дне этих лесов. В лесах преобладают дубы, клены и березы. Вы также можете найти вечнозеленые деревья, такие как пихта и сосна, в лиственных лесах умеренного пояса.
Большинство животных, населяющих эти леса, приспособлены к холодным зимам.Некоторые из видов, которые вы найдете в лиственных лесах умеренного пояса, включают ястребов, рыжих лис, кардиналов и дятлов.
Хвойные леса умеренного пояса
Хвойные леса умеренного пояса обычно встречаются в прибрежных районах с мягкой зимой и обильными дождями. Вы также можете найти их в горных районах с мягким климатом. В эти регионы входят:
- Новая Зеландия
- Тихоокеанский Северо-Запад США и Канада
- Южная Япония
- Юго-западная Южная Америка
- Некоторые части Северо-Западной Европы
Хвойные леса умеренного пояса характеризуются влажным климатом и продолжительным вегетационным периодом из-за большого количества осадков.В лесах выпадает от 50 до 200 дюймов дождя в год. Почвы в основном богаты толстым слоем разлагающегося вещества.
В этих лесах преобладают вечнозеленые хвойные породы. Деревья вырастают очень высокими. Это объясняется высоким уровнем осадков и умеренными температурами.
Вы должны ожидать увидеть такие деревья, как кипарис, кедр, сосна, пихта Дугласа, красное дерево и ель в хвойных лесах умеренного пояса. Также распространены некоторые лиственные деревья, такие как клен, а также папоротники и мох.
Животные, населяющие эти леса, включают лосей, оленей, сурков, черных медведей, мраморных козлов и пятнистых сов.
3. Северные леса
Также известные как таежные леса, бореальные леса расположены между 50 o и 60 o северной широты. Вы можете найти их в Сибири, Канаде, Северной Азии и Скандинавии. Стоит отметить, что около 65 процентов бореальных лесов находятся в Скандинавии.
Для таежных лесов характерно короткое лето и продолжительная зима.Ежегодно они получают от 15 до 40 дюймов осадков, большая часть которых выпадает в виде снега. В этих лесах обычно тонкая почва из-за низких температур, которые снижают скорость разложения.
Деревья в бореальных лесах в основном вечнозеленые. Примеры деревьев включают сосну, ель и пихту. Лесная подстилка имеет ограниченную растительность из-за плотного полога.
Животные, обитающие в этих лесах, обычно имеют густой мех и приспособлены к долгим холодным зимам.Вы должны ожидать найти таких животных, как олень, лось, заяц на снегоступах, карибу, лось, рыси, волки и росомахи.
Теперь, когда вы знаете основные типы лесов, давайте посмотрим на важность лесов.
Значение лесов
Мировые леса играют важную роль не только в благополучии их жителей, но и в общем состоянии здоровья других форм жизни на планете, включая человека. Некоторые из основных преимуществ лесов включают следующее:
1.Климатическая стабильность
Леса вносят большой вклад в способность Земли поддерживать свой климат благодаря эффектам фотосинтеза. Они помогают предотвратить изменение климата, поскольку отводят углекислый газ, вызывающий парниковый эффект, и выделяют кислород в окружающую среду.
Таким образом, они помогают очищать атмосферу и контролировать повышение температуры.
2. Экологические преимущества
Леса помогают предотвратить эрозию почвы, сводя к минимуму количество осадков на поверхности почвы.Они также впитывают воду, предотвращая сток, который иначе смыл бы верхний слой почвы.
Кроме того, леса действуют как фильтры для воды. Они собирают и хранят воду в дополнение к пополнению подземных водоносных горизонтов. Леса также повышают влажность атмосферы за счет потоотделения, которое влияет на количество осадков и температуру.
3. Биоразнообразие
Ни один другой биом на Земле не содержит большего разнообразия биоразнообразия, чем леса. Это объясняет, почему изучена лишь небольшая часть видов, встречающихся в лесах.Огромный спектр деревьев и растений, произрастающих в тропических лесах, отличается особенно высоким биоразнообразием.
4. Экономическое значение
Мы получаем многочисленные выгоды от лесных экосистем. Многие сообщества по всему миру выживают за счет растительности и животных, обитающих в лесах.
Большинство лекарств и фармацевтических препаратов производятся из растений, выращенных в лесу. Многие обрабатывающие отрасли получают сырье из леса, например дерево и бумагу. К другим полезным лесным продуктам относятся камеди, специи и красители.
Леса также добавляют эстетики окружающей среде и способствуют развитию экотуризма.
Итог
Леса — это большие площади, покрытые деревьями, которые занимают около трети поверхности земли. Польза лесов для их обитателей и разнообразия жизни безгранична. Они поддерживают стабильность климата, способствуют сохранению биоразнообразия и способствуют экономическому развитию. И они являются крупнейшими в мире хранилищами углерода после океанов. Следовательно, крайне важно, чтобы леса были защищены от вырубки лесов и других пагубных последствий человеческой цивилизации.
Автор фотографии: pexels
лес | Определение, типы и факты
лес , сложная экологическая система, в которой деревья являются доминирующей формой жизни.
Леса с преобладанием деревьев могут возникать там, где температура поднимается выше 10 ° C (50 ° F) в самые теплые месяцы, а годовое количество осадков превышает 200 мм (8 дюймов). Они могут развиваться в различных условиях в этих климатических пределах, и вид почвы, растений и животных различается в зависимости от экстремальных воздействий окружающей среды.В прохладных высокоширотных приполярных регионах в лесах преобладают выносливые хвойные породы, такие как сосны, ели и лиственницы. Эти таежные (бореальные) леса имеют продолжительные зимы и от 250 до 500 мм (от 10 до 20 дюймов) осадков в год. В более умеренном климате высоких широт преобладают смешанные леса как из хвойных, так и из широколиственных лиственных пород. Широколиственные лиственные леса развиваются в климате средних широт, где средняя температура выше 10 ° C (50 ° F) в течение не менее шести месяцев каждый год, а годовое количество осадков превышает 400 мм (16 дюймов).Вегетационный период от 100 до 200 дней позволяет преобладать в лиственных лесах дубами, вязами, березами, кленами, буками и осинами. Во влажном климате экваториального пояса развиваются тропические леса. Там сильные дожди поддерживают вечнозеленые растения, у которых вместо хвои широкие листья, как в более прохладных лесах. В более низких широтах Южного полушария вновь появляется умеренный лиственный лес.
Типы лесов отличаются друг от друга по видовому составу (который частично определяется возрастом леса), густоте древесного покрова, типу почв, обнаруженных здесь, и геологической истории лесного региона.
Почвенные условия различаются по глубине, плодородию и наличию многолетних корней. Глубина почвы важна, потому что она определяет степень проникновения корней в землю и, следовательно, количество воды и питательных веществ, доступных деревьям. Почва таежных лесов песчаная, быстро дренируемая. В лиственных лесах почва коричневая, более богатая питательными веществами, чем песок, и менее пористая. Тропические леса и леса саванны имеют слой почвы, богатый железом или алюминием, что придает почве красноватый или желтоватый оттенок.Количество воды, доступной для почвы и, следовательно, доступной для роста деревьев, зависит от количества годовых осадков. Вода может теряться в результате испарения с поверхности или транспирации листьев. Испарение и транспирация также контролируют температуру воздуха в лесах, которая всегда немного теплее в холодные месяцы и прохладнее в теплые месяцы, чем воздух в окружающих регионах.
саванна в КенииСолнце садится в саванне в африканской стране Кения.
© Digital Vision / Getty Images Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту.Подпишитесь сейчасПлотность древесного покрова влияет на количество солнечного света и дождя, достигающего каждого яруса леса. Лес под пологом поглощает от 60 до 90 процентов доступного света, большая часть которого поглощается листьями для фотосинтеза. На движение осадков в лес в значительной степени влияет листовой покров, который имеет тенденцию замедлять скорость падающей воды, которая проникает до уровня земли, стекая по стволам деревьев или капая с листьев. Вода, не поглощаемая корнями деревьев для питания, проходит по корневым каналам, поэтому водная эрозия не является основным фактором формирования рельефа леса.
Знать о роли грибов в лесной экосистеме, где одни переваривают растительное вещество, а другие взаимодействуют с растением (микоризный) взаимно
Узнают о грибах в лесной экосистеме, в том числе о том, как некоторые грибы переваривают растительное вещество (сапротрофизм) ), а другие образуют мутуалистические (микоризные) отношения с растениями.
© Открытый университет (издательский партнер Britannica) См. Все видео к этой статьеУзнайте, как бактерии перерабатывают лесную подстилку в воду, углекислый газ, азот и минералы почвы
Разложение лесной подстилки и ее минерализация.
Encyclopædia Britannica, Inc. См. Все видео для этой статьи
Леса относятся к числу самых сложных экосистем в мире, и они демонстрируют обширную вертикальную стратификацию. Хвойные леса имеют самую простую структуру: слой деревьев, поднимающийся до высоты около 30 метров, слой кустарников, который является пятнистым или даже отсутствует, и слой земли, покрытый лишайниками, мхами и печеночниками. Лиственные леса более сложны; полог дерева разделен на верхний и нижний ярусы, в то время как пологи тропического леса разделены как минимум на три слоя.Лесная подстилка в обоих этих лесах состоит из слоя органического вещества, покрывающего минеральную почву. Слой гумуса тропических почв подвержен воздействию высоких уровней тепла и влажности, которые быстро разлагают все существующие органические вещества. Грибы на поверхности почвы играют важную роль в доступности и распределении питательных веществ, особенно в северных хвойных лесах. Некоторые виды грибов живут вместе с корнями деревьев, в то время как другие паразитически разрушительны.
Животные, обитающие в лесах, обладают высокоразвитым слухом, и многие из них приспособлены к вертикальному перемещению в окружающей среде. Поскольку пищи, кроме наземных растений, мало, многие наземные животные используют леса только в качестве укрытия. В лесах умеренного пояса птицы разносят семена растений, а насекомые помогают в опылении вместе с ветром. В тропических лесах опыление производят летучие мыши и птицы. Лес — самая эффективная экосистема природы, с высокой скоростью фотосинтеза, влияющей как на растительные, так и на животные системы в серии сложных органических взаимоотношений.
Типы лесов: определения, примеры и значение
В глобальном масштабе леса формируются под воздействием солнечной радиации и осадков, на которые влияет широта. Эти климатические условия определяют, какие организмы могут выжить в той или иной области, и помогали формировать эволюцию лесов на протяжении миллионов лет. В зависимости от широты существует три типа лесов: северный, умеренный и тропический.
Бореальные леса, расположенные на крайнем севере, отличаются долгой холодной зимой с коротким вегетационным периодом.Умеренные леса, расположенные в средних широтах, имеют четыре различных сезона. Тропические леса, расположенные вдоль экватора, отличаются высокими температурами, продолжительным вегетационным периодом и обладают невероятным биоразнообразием.
Леса поддерживают людей в местном и региональном масштабах, обеспечивая экосистемные услуги, такие как опыление, регулирование климата и сохранение почвы. По данным Продовольственной и сельскохозяйственной организации Объединенных Наций (ФАО), несмотря на ценность нетронутых лесов для благополучия людей, леса во всем мире находятся под угрозой из-за деятельности человека.
Что такое лес?
Лес — это экосистема, в которой преобладают деревья. Согласно параметрам, установленным ФАО, площадь должна составлять не менее половины гектара или около одной с четвертью акра, чтобы считаться лесом. Деревья в этом районе также должны иметь возможность вырасти до высоты более 16 футов и иметь навес, покрывающий не менее 10% неба.
Несмотря на точное определение, данное ФАО, все еще существуют разногласия по поводу того, что представляет собой лес.Одна из проблем интерпретации организации заключается в том, что она не делает различий между естественными и лесонасаждениями. Согласно исследованию ведущих лесных экологов, опубликованному в журнале Ambio, поскольку нынешнее определение леса не делает различий между типами леса, может быть сложно отслеживать изменения в количестве леса.
Северные леса
Озеро Гарибальди, Уистлер, Британская Колумбия. Маркус Тиммс / Getty ImagesБореальные леса, или тайга, находятся между 50 и 60 градусами широты в Северной Америке, Азии и Европе.Под бореальными лесами находится земля, образованная ледниками, оставившими наследие геологии, гидрологии и почвам этого региона. Суровый холодный климат бореальных лесов затрудняет жизнь, что приводит к низкому видовому разнообразию по сравнению с лесами умеренного и тропического пояса. Растения и животные, обитающие в бореальных лесах, специально приспособлены к короткому вегетационному сезону и низким температурам. Из-за своей протяженности и удаленности северные леса являются важными хранилищами углерода.
Из трех типов леса бореальные леса имеют самый короткий вегетационный период, около 130 дней.Бореальные леса обычно имеют мелкие, кислые, бедные питательными веществами почвы. Хвойные деревья являются наиболее распространенным типом деревьев, хотя есть и некоторые хорошо адаптированные лиственные деревья, такие как ивы, тополя и ольха. Видные виды включают пихту черную и белую, пихту бальзамическую и тамарак. В подлеске кусты черники и клюквы являются высокоэнергетической пищей для диких животных.
Взрослая дикая рысь, lynx canadensis, в Канадских Скалистых горах. Коллин Гара / Getty ImagesЖивотные, обитающие в бореальных лесах, специально приспособлены к экстремально низким температурам — до -22 F (-30 C) — и низкой доступности ресурсов в течение значительных периодов года.Северный карибу — одно из немногих животных, которые живут в тайге круглый год, и они выживают, занимая площади почти в миллион акров в поисках пищи. Однако этот некогда многочисленный карибу сейчас находится под угрозой исчезновения из-за потери среды обитания и инфраструктуры, вырубающей оставшиеся леса. Многие виды птиц посещают водно-болотные угодья бореальных лесов во время своих ежегодных миграций, перемещаясь на юг, когда температура падает, а пищи становится мало.
Изменение климата представляет собой серьезную угрозу для бореальных лесов. Почти 80% бореальных лесов покрыты вечной мерзлотой — слоем почвы, которая остается мерзлой в течение всего года.Поскольку температура повышается неестественно быстрыми темпами, земля становится мягкой и болотистой, и многие деревья в конечном итоге теряют устойчивость и умирают. Ученые из Международной ассоциации исследования бореальных лесов считают, что сохранение бореальных лесов является ключом к замедлению изменения климата.
Типы бореальных лесов
- Открытый бореальный полог: также известный как лишайниковый лес, бореальный лес с открытым пологом встречается в более высоких широтах и отличается меньшим видовым разнообразием.
- Closed Canopy Boreal: Обитающие в более низких широтах бореальные леса с закрытым пологом имеют более богатую почву и более густые насаждения, которые пропускают мало света на лесную подстилку.Однако менее суровые условия приводят к большему видовому разнообразию.
Леса умеренного пояса
Лиственный лес умеренного пояса в Армении. 1255К / Getty ImagesЛеса умеренного пояса расположены в средних широтах, что дает им характерные четыре сезона. Осталось очень мало участков старовозрастного леса умеренного пояса; в зоне преобладают вторичные леса. По состоянию на 2020 год на леса умеренного пояса приходилось 16% от общего лесного покрова Земли.
В лесах умеренного пояса обитают виды, приспособленные к сезонности.Лиственные деревья, такие как клены, гикори, дубы и многие другие, сбрасывают листья и переходят в спячку осенью и зимой для экономии энергии. Медведи, рыси, белки и олени живут в лесах с умеренным климатом и могут хранить пищу, адаптировать свой рацион или впадать в спячку, чтобы справиться с недостатком питательной пищи зимой.
Хотя леса умеренного пояса имеют общую сезонность, они сильно различаются по годовым осадкам и температуре. Годовая температура колеблется от -22 F до 86 F в зависимости от местоположения и сезона.В лесах умеренного пояса выпадает в среднем от 30 до 59 дюймов дождя в год. Почвы, как правило, плодородные, с толстым слоем органических веществ, из которых растения могут извлекать питательные вещества для роста.
Самка красного волка (Canis rufus). Джефф Гоулден / Getty ImagesЛеса умеренного пояса являются домом для многих исчезающих видов. В США 12 видов млекопитающих, внесенных в список находящихся под угрозой исчезновения Службой охраны рыб и дикой природы, живут в лесах умеренного пояса. Красный волк, произрастающий в лесах умеренного пояса на востоке Северной Каролины, внесен в список МСОП как находящийся под угрозой исчезновения.Северная пятнистая сова была внесена в федеральный список как находящийся под угрозой исчезновения в 1990 году и в настоящее время считается находящимся под угрозой исчезновения. Эти хищные птицы предпочитают среду обитания в старовозрастных лесах Вашингтона, Орегона и Калифорнии, численность которых в последние десятилетия продолжает сокращаться.
Типы леса умеренного пояса
- Лиственный лес: В этом типе леса преобладают лиственные деревья, которые теряют листья в холодные месяцы и переходят в период покоя.
- Хвойный лес: в этом биоме больше вечнозеленых шишковидных деревьев.
- Тропический лес умеренного климата: с умеренными температурами в этих лесах выпадает чрезвычайно большое количество осадков — от 140 до 167 дюймов в год.
Тропические леса
Вечнозеленый облачный лес на склонах горы Рувензори в Центральной Африке. guenterguni / Getty ImagesРасположенные между тропиками Рака и Козерога на 23 градусе северной и южной широты, тропические леса являются одними из самых биоразнообразных экосистем на Земле. Эти леса покрывают лишь десятую часть поверхности планеты, но при этом служат убежищем для половины всех видов.Им также наиболее угрожает деятельность человека.
Тропические леса имеют относительно стабильные условия, которые позволяют жизни процветать. Это самые теплые и дождливые леса на Земле с температурой от 68 F до 77 F, ежегодно выпадает от 79 до 394 дюймов дождя.
Тропические леса известны своим необычайным биоразнообразием. Например, тропические леса Амазонки являются домом для 10% описанных видов в мире.
Разнообразие тропических лесов делает их очень эффективными в переработке питательных веществ.Мертвое и разлагающееся вещество быстро разрушается деструкторами и почти мгновенно поглощается другим организмом. Это делает почвы тропических лесов бедными питательными веществами. Чтобы справиться с бедными почвами, многие тропические деревья адаптировали мелкую корневую систему, которая распространяется по лесной подстилке и может легче усваивать питательные вещества.
Обезьяна тамарин с хлопковой вершиной. Рональд Виттек / Getty ImagesМногие харизматичные виды тропических лесов находятся под угрозой исчезновения. Например, африканский лесной слон, обитающий в Западной и Центральной Африке, внесен в список МСОП как находящийся под угрозой исчезновения из-за потери среды обитания и браконьерства.Приматы живут почти исключительно в тропиках, и большинство из них обитает в тропических лесах. В некоторых бразильских лесах на одной территории обитает до 13 видов приматов.
Человеческая деятельность, такая как вырубка леса, расчистка земель для ведения сельского хозяйства и браконьерство, представляет собой угрозу для будущего тропических лесов. По данным Института мировых ресурсов, только в 2020 году было потеряно более 12 миллионов гектаров тропических лесов.
Типы тропических лесов
- Вечнозеленые тропические леса: часто называемые «настоящими» тропическими лесами, это самые влажные (~ 80 дюймов дождя в год) и самые биоразнообразные тропические леса.
- Тропический влажный лес: дальше от экватора, чем вечнозеленые влажные леса, в тропических влажных лесах в целом меньше осадков и больше разница между сезонами.
- Сухой тропический лес: от четырех до шести месяцев в году выпадает очень мало дождя. У растений и животных есть особые приспособления, чтобы справиться с этим периодом нехватки воды.
- Мангровые заросли: прибрежные тропические леса с деревьями, приспособленными для жизни в солоноватой воде с меняющимся уровнем. Мангровые заросли защищают побережье от штормов и служат рассадниками водных видов
Леса — Типы лесов — Виды, Вечнозеленые, Деревья и Покрытосеменные
Многие страны разработали национальные схемы экологической классификации своих лесов.Обычно эти схемы основаны на биофизической информации и отражают естественные крупномасштабные закономерности видового состава, типа почвы, топографии и климата. Однако эти классификации могут сильно различаться в разных странах, даже для схожих типов лесов.
Международная система классификации экосистем была предложена научной рабочей группой под эгидой Организации Объединенных Наций по вопросам образования, науки и культуры (ЮНЕСКО). В этой схеме перечислены 24 типа лесов, разделенных на два широких класса: (i) леса с закрытым пологом с пологом не менее 16.5 футов (5 м) в высоту и с переплетающимися кронами деревьев и (ii) редколесья с относительно редким и более коротким пологом. Выбор типов леса в этих двух широких классах описан ниже:
- Тропические и субтропические леса.
- Тропический лес. Это богатый видами лес покрытосеменных деревьев (иногда называемых «тропическими лиственными деревьями»), произрастающих в условиях обильных дождей и постоянных высоких температур. Следовательно, виды в этой экосистеме не терпимы ни к засухе, , ни к морозу, а сам лес обычно находится в старовозрастном состоянии.Большая часть наземного биоразнообразия Земли встречается в лесных экосистемах этого типа.
- Тропический и субтропический вечнозеленый лес. Это также довольно богатый видами лес, но он встречается в регионах с редкими сезонными дождями. Отдельные деревья могут сбрасывать листья, обычно в результате относительно засушливых условий. Однако не все виды делают это одновременно, поэтому полог всегда в значительной степени покрыт листвой.
- Тропический и субтропический засухолиственный лес.Это относительно открытый покрытосеменный лес, в котором листва деревьев сбрасывается незадолго до засушливого сезона, который обычно наступает зимой.
- Мангровый лес. Это относительно бедный видами лес, расположенный в илистой приливной зоне местообитания в тропиках и субтропиках. В мангровых лесах преобладают вечнозеленые покрытосеменные деревья, устойчивые к затопленным почвам и воздействию соли . Широко распространены некоторые роды мангровых деревьев. Примерами из южной Флориды являются красные мангровые заросли ( Rhizophora mangle ) с их характерными корнями на ходулях и черные мангровые заросли ( Avicennia nitida ) с их пневматофорами, которые выходят из бедных кислородом отложений в атмосферу.
- Леса умеренного и субполярного пояса.
- Лиственный лес умеренного пояса. Это лиственный лес, в котором преобладают различные виды покрытосеменных деревьев, произрастающих в сезонных климатических условиях, включая умеренно холодные зимы. В лесах этого типа на востоке Северной Америки распространены такие виды деревьев, как клен, береза, гикори, ясень, грецкий орех, тюльпанное дерево, дуб и липа , среди прочих ( Acer, Betula, Carya, Fraxinus, Juglans, Liriodendron, Quercus, и Tilia соответственно).
- Умеренный и субарктический вечнозеленый хвойный лес. Это северный хвойный лес (иногда называемый бореальным лесом), произрастающий в регионах с очень сезонными условиями, включая суровые зимы. Доминирующими родами хвойных деревьев являются пихта, ель , сосна, кедр и болиголов, среди прочих ( Abies, Picea, Pinus, Thuja, и Tsuga соответственно).
- Вечнозеленые тропические леса умеренного и приполярного пояса. Этот лес произрастает во влажной, незамерзающей океанической среде Южного полушария, и в нем преобладают вечнозеленые виды покрытосеменных, такие как бук южный ( Nothofagus spp.) и южной сосны ( Podocarpus spp.).
- Умеренный, зимне-дождевой, вечнозеленый широколиственный лес. Это вечнозеленый лес покрытосеменных растений, произрастающий в регионах с ярко выраженным влажным сезоном, но с летней засухой. В Северная Америка , этот тип леса встречается в прибрежных частях южной Калифорнии, и преобладают вечнозеленые виды дубов ( Quercus spp.).
- Холодно-лиственный лес. Это северный лиственный лес, растущий в строго сезонном климате с очень холодной зимой.В этом типе леса обычно преобладают покрытосеменные деревья, такие как осина и береза ( Populus spp. И Betula spp.) Или лиственные хвойные деревья, лиственница ( Larix spp.).
Тип и высота леса важны в формировании высотного изменения радиальной реакции роста на изменение климата
IPCC. Изменение климата 2007: воздействия, адаптация и уязвимость . (Издательство Кембриджского университета, 2007).
Лара, А. и др. . Пространственные и временные изменения в росте Nothofagus pumilio на линии деревьев вдоль его широты (35 ° 40′ – 55 ° ю.ш.) в чилийских Андах. J. Biogeogr. 32 , 879–893 (2005).
Артикул Google Scholar
Хуанг Дж. и др. . Реакция радиального роста четырех доминирующих бореальных видов деревьев на климат вдоль широтного градиента в восточно-канадских бореальных лесах. Global Change Biol. 16 , 711–731 (2010).
ADS Статья Google Scholar
Бабст, Ф. и др. . Реакция роста лесов на климат на европейском континенте, зависящая от участков и видов. Глобальная экология и биогеография 22 , 706–717 (2013).
Артикул Google Scholar
Закон, Б.Е. и др. . Нарушения и климатические воздействия на запасы и потоки углерода в Западном Орегоне, США. Global Chang Biology 10 , 1429–1444 (2004).
ADS Статья Google Scholar
Д’Амато, А.В., Брэдфорд, Дж. Б., Фрейвер, С. и Палик, Б. Дж. Влияние рубок ухода за кожей на уязвимость к засухе и реакцию на климат в лесных экосистемах северного умеренного пояса. Ecol. Прил. 23 , 1735–1742 (2013).
Артикул Google Scholar
Аллен К. Д., Бреширс Д. Д. и МакДауэлл Н. Г. О недооценке глобальной уязвимости к гибели деревьев и вымиранию лесов в результате более жаркой засухи в антропоцене. Экосфера 6 , 1–55 (2015).
Артикул Google Scholar
Уильямс, А. П. и др. . Температура как мощный фактор стресса от засухи в лесах и гибели деревьев. Природа изменения климата 3 , 292–297 (2013).
ADS Статья Google Scholar
Адамс, Х. Д. и Колб, Т. Е. Реакция роста деревьев на засуху и температуру в горном ландшафте в северной Аризоне, США. J. Biogeogr. 32 , 1629–1640 (2005).
Артикул Google Scholar
Ван, Т., Чжан, К.Б. и Ма, К. П. Динамика лесов в зависимости от климатической изменчивости в центральных горах Тянь-Шань на северо-западе Китая. Global Ecol. Биогеогр. 15 , 406–415 (2006).
Артикул Google Scholar
Ферреро М. Э. и др. . Реакция роста деревьев на градиенты окружающей среды в субтропических лесах Аргентины. Plant Ecol. 214 , 1321–1334 (2013).
Артикул Google Scholar
Чен П.-Й., Уэлш К. и Хаманн А. Географические вариации реакции роста пихты Дугласовой на межгодовую изменчивость климата и прогнозируемое изменение климата. Global Change Biol. 16 , 3374–3385 (2010).
ADS Статья Google Scholar
Поултер Б. и др. . Последние тенденции динамики лесов Внутренней Азии в зависимости от температуры и осадков указывают на высокую чувствительность к изменению климата. Сельскохозяйственная и лесная метеорология 178–179 , 31–45, https://doi.org/10.1016/j.agrformet.2012.12.006 (2013).
ADS Статья Google Scholar
Чанг, Дж. Ф., Ван, X. П., Чжан, X. П. и Лин, X. Динамика альпийской границы леса в зависимости от климатической изменчивости в северных горах Дасинъань. Журнал горной науки 27 , 703–711 (2009).
Google Scholar
Цзэн, Л. Б. и др. . Рост экотона альпийских лесов в зависимости от климатической изменчивости для лесов Picea crassifolia в средней части гор Цилиан, северо-запад Китая. Журнал Пекинского университета лесного хозяйства 34 , 50–56 (2012).
Google Scholar
Сидор, К. Г., Попа, И., Влад, Р. и Керубини, П. Различные реакции древесных колец европейской ели на температуру воздуха через высотный градиент в Восточных Карпатах (Румыния). Деревья 29 , 985–997 (2015).
Артикул Google Scholar
Wu, Y. et al. . Тест BIOME-BGC с дендрохронологией для лесов вдоль высотного градиента горы Чанбайшань на северо-востоке Китая. Журнал экологии растений 10 , 415–425, https://doi.org/10.1093/jpe/rtw076 (2017).
Артикул Google Scholar
Лян, П. Х. и др. . Реакция роста широколиственных и смешанных лесов кедра корейского на разных этапах сукцессии на изменение климата в заповеднике Шэншань провинции Хэйлунцзян, Китай. Китайский журнал экологии растений 40 , 425–435 (2016).
Артикул Google Scholar
Райан М. и Йодер Б. Дж. Гидравлические ограничения по высоте и росту деревьев. Bioscience 47 , 235–242 (1997).
Артикул Google Scholar
Райан М. Г., Филлипс Н. и Бонд Б. Дж. Гипотеза гидравлического ограничения пересмотрена. Plant, Cell Environ. 29 , 367–381 (2006).
Артикул Google Scholar
Мериан П. и Лебуржуа Ф. Взаимосвязь климата и роста, опосредованная размером в лесах умеренного пояса: многовидовой анализ. Экология и управление лесами 261 , 1382–1391 (2011).
Артикул Google Scholar
Мартинес-Вилалта, Дж., Лопес, Б. К., Лепфе, Л. и Льорет, Ф. Детерминанты реакции радиального роста сосны обыкновенной на засуху на уровне древостоя и деревьев. Oecologia 168 , 877–888 (2012).
ADS Статья Google Scholar
Айба С. И. и Китайма К. Структура, состав и видовое разнообразие в матрице высот-субстрат сообществ тропических лесов на горе Кинабалу, Борнео. Plant Ecol. 140 , 139–157 (1999).
Артикул Google Scholar
Симард, М., Пинто, Н., Фишер, Дж. Б. и Баччини, А. Глобальное картирование высоты лесного покрова с помощью космического лидара. Журнал геофизических исследований 116 , G04021 (2011).
ADS Статья Google Scholar
Ван, X. P., Fang, J. Y., Tang, Z. Y. & Zhu, B. Климатический контроль первичной структуры леса и аллометрия высоты DBH в Северо-Восточном Китае. Для. Ecol. Управлять. 234 , 264–274 (2006).
Артикул Google Scholar
Fang, J. Y. и др. . Обследование лесных сообществ и структурные характеристики лесов в Китае. Экография 35 , 1059–1071, https://doi.org/10.1111/j.1600-0587.2013.00161.x (2012).
Артикул Google Scholar
Ван Б. и др. . Реакция роста и климата ели Цинхай (Picea crassifolia) между нижним и верхним пределами градиента: тематическое исследование вдоль последовательного склона в районе середины гор Цилиан. Науки об окружающей среде и Земле 75 , 236 (2016).
Артикул Google Scholar
Fritts, H.C. Годовые кольца и климат . (Academic Press, 1976).
Каллен, Л. Э. и Грирсон, П. Ф. Изменчивость осенне-зимних осадков в масштабе нескольких десятилетий в юго-западной Австралии с 1655 года нашей эры, реконструированная по годичным кольцам Callitris columellaris. Климатическая динамика 33 , 433–444 (2009).
ADS Статья Google Scholar
Латте, Н., Лебуржуа, Ф. и Клаессенс, Х. Повышенная синхронизация роста деревьев у бука (Fagus sylvatica L.) в ответ на изменение климата в северо-западной Европе. Dendrochronologia 33 , 69–77, https://doi.org/10.1016/j.dendro.2015.01.002 (2015).
Артикул Google Scholar
Вебер, П., Бугманн, Х., Плюсс, А. Р., Вальтерт, Л. и Риглинг, А. Реакция на засуху и изменение средней чувствительности европейского бука близко к пределу сухого распределения. Деревья — структура и функции 27 , 171–181, https://doi.org/10.1007/s00468-012-0786-4 (2013).
Артикул Google Scholar
Чжан, Ф., Чи, З. и Ли, X. Анализ и первичная оценка климата горной местности Чанбайшань. Исследование лесной экосистемы 1 , 193–214 (1980).
Google Scholar
Wang, Z. и др. . Основные типы лесов и их особенности структуры сообществ северного склона горы Чанбайшань (I). Исследование лесной экосистемы 1 , 25–42 (1980).
Google Scholar
Schweingruber, F. H. Годовые кольца и окружающая среда: дендроэкология . (Пауль Хаупт А.Г. Берн, 1996).
Холмс, Р. Л. Компьютерный контроль качества при датировании и измерении годичных колец. Бюллетень Tree-Ring 43 , 69–78 (1983).
Google Scholar
Кук, Э. Р. и Холмс, Р. Л. Руководство пользователя программы ARSTAN. Лаборатория исследования древесных колец, Университет Аризоны, Тусон, США (1986).
Бриффа, К. Р. Взаимосвязь между деревьями и климатом и дендроклиматологическая реконструкция на Британских островах , Университет Восточной Англии (1984).
Ван, X. П., Фанг, Дж. Ю. и Чжу, Б. Биомасса лесов и распределение корневых побегов на северо-востоке Китая. Для. Ecol. Управлять. 255 , 4007–4020 (2008).
Артикул Google Scholar
Гастон, К. Дж. Глобальные закономерности в биоразнообразии. Nature 405 , 220–227 (2000).
CAS Статья Google Scholar
Wang, X. P., Ouyang, S., Sun, O. J. и Fang, J. Y. Структура лесной биомассы на северо-востоке Китая в значительной степени определяется высотой леса. Для. Ecol. Управлять. 293 , 149–160, https://doi.org/10.1016/j.foreco.2013.01.001 (2013).
Артикул Google Scholar
Дункансон, Л. И., Дубая, Р. О. и Энквист, Б. Дж. Оценка общих закономерностей структуры леса: количественная оценка взаимосвязи аллометрического масштабирования деревьев и лесов в Соединенных Штатах. Global Ecol. Биогеогр. 24 , 1465–1475 (2015).
Артикул Google Scholar
Сан, Х. и др. . Что вызывает большие отклонения от предсказаний теории метаболического масштабирования в более ранних сукцессионных лесах? Экология и управление лесами 405 , 101–111 (2017).
Артикул Google Scholar
Джонсон, Дж. Б. и Омланд, К. С. Выбор модели в экологии и эволюции. Trends Ecol. Evol. 19 , 101–108 (2004).
Артикул Google Scholar
Грёмпинг, У. Относительная важность линейной регрессии в R: пакет relaimpo. Журнал статистического программного обеспечения 17 , 1-27 (2006).
Артикул Google Scholar
Грёмпинг, У. Важность переменных в регрессионных моделях. Междисциплинарные обзоры Wiley: вычислительная статистика 7 , 137–152 (2015).
MathSciNet Статья Google Scholar
Джонсон, Дж. У. и ЛеБретон, Дж. М. История и использование индексов относительной важности в организационных исследованиях. Организационные методы исследования 7 , 238–257 (2004).
Артикул Google Scholar
Zhang, J., Nielsen, S.E., Mao, L., Chen, S. & Svenning, J.-C. Региональные и исторические факторы дополняют текущий климат в формировании глобальной высоты лесного полога. J. Ecol. 104 , 469–478, https://doi.org/10.1111/1365-2745.12510 (2016).
Артикул Google Scholar
Liang, E., Wang, Y., Xu, Y., Liu, B. & Shao, X. Изменчивость роста Abies georgei var. smithii вдоль высотных уклонов в горах Сыгера, на юго-востоке Тибетского плато. Деревья 24 , 363–373 (2010).
Артикул Google Scholar
Gou, X.H. et al. . Анализ хронологии годичных колец гор Цилиан на разной высоте. Acta ecologica sinica 24 , 172–176 (2004).
Google Scholar
Ратгебер, К. Б., Росси, С. и Бонтемпс, Дж. Д. Кембиальная активность, связанная с размером дерева на плантации зрелой серебристой пихты. Энн Бот 108 (3), 429–438 (2011).
Артикул Google Scholar
Де Луис, М., Новак, К., Чуфар, К. и Равентос, Дж. Связь климата и роста, опосредованная размером, у Pinus halepensis и Pinus pinea. Деревья 23 , 1065–1073 (2009).
Артикул Google Scholar
Мартин-Бенито, Д., Керубини, П., Del Río, M. & Cañellas, I. Реакция роста Pinus nigra Arn на климат и засуху. деревья разных классов кроны. Деревья 22 , 363–373 (2008).
Артикул Google Scholar
West, P. W. Измерение деревьев и лесов . (Springer, 2009).
Кимминс, Дж. П. Экология леса . (Macmillan Publishing Company, 1987).
Сюй, К. и др. . Ширина годичных колец является хорошим показателем годового изменения продуктивности лесов в лесах умеренного пояса. Scientific Reports 7 , 1945, https://doi.org/10.1038/s41598-017-02022-6 (2017).
ADS CAS Статья PubMed PubMed Central Google Scholar
Лю Л. и др. . Рост деревьев и его климатический сигнал вдоль широтных и высотных градиентов: сравнение годичных колец между Финляндией и Тибетским плато. Biogeosciences Discuss ., Https://doi.org/10.5194/bg-2016-5559 (2017).
Ю., Д. и др. . Пространственная изменчивость и временная нестабильность во взаимосвязи климата и роста корейской сосны в горном регионе Чанбайшань на северо-востоке Китая. Для. Ecol. Управлять. 300 , 96–105 (2013).
Артикул Google Scholar
Körner, C. & Paulsen, J. Всемирное исследование температур на высотных деревьях. J Biogeography 31 , 713–732 (2004).
Артикул Google Scholar
Кёрнер, К. Переоценка положения высотных линий деревьев и их объяснение. Oecologia 115 , 445–459 (1998).
ADS Статья Google Scholar
Ллойд А. Х., Банн А. Г. и Бернер Л. Широтный градиент реакции роста деревьев на потепление климата в сибирской тайге. Global Change Biol. 17 , 1935–1945 (2011).
ADS Статья Google Scholar
Клос, Р. Дж., Ван, Г. Г., Бауэрле, В. Л. и Рик, Дж. Р. Влияние засухи на рост и смертность лесов на юго-востоке США: анализ с использованием данных о состоянии и мониторинге лесов. Ecol. Прил. 19 , 699–708 (2009).
Артикул Google Scholar
Льорет Ф., Килинг Э. Г. и Сала А. Компоненты устойчивости деревьев: последствия последовательных эпизодов замедленного роста в старых сосновых лесах пондерозы. Oikos 120 , 1909–1920 (2011).
Артикул Google Scholar
Olivar, J., Bogino, S., Rathgeber, C., Bonnesoeur, V. & Bravo, F. Прореживание оказывает положительное влияние на динамику роста и взаимосвязь между ростом и климатом сосны Алеппо (Pinus halepensis) деревья разных классов кроны. Annals of Forest Science 71 , 395–404, https://doi.org/10.1007/s13595-013-0348-y (2014).
Артикул Google Scholar
лиственные
Лес: Растения Высокие лиственные деревья составляют верхний слой роста растений, и они создают умеренно густой полог леса. Хотя навес умеренно плотный, он позволяет солнечному свету достигать лесной подстилки. Этот солнечный свет позволяет растения в других слоях для роста. Второй слой роста растений включает саженцы и виды деревьев, которые от природы короче ростом.Третий слой (или подлесок) будет включать кусты. Лесные травы, такие как полевые цветы и ягоды, составляют четверть слой. Весной, до того, как лиственные деревья распускаются, эти травы цветут и быстро растут, чтобы использовать в своих интересах солнечный свет. Пятый слой будет включать мхи и лишайники, растущие на деревьях. стволы. Завод
приспособления Более низкие температуры и ограниченный солнечный свет — это два климатических условия, которые говорят дереву чтобы начать адаптироваться. Осенью, когда возникают эти условия, дерево перекрывает подачу воды к листьям и перекрывает область между стволом листа и стволом дерева. С ограниченным солнечным светом и вода, лист не может продолжать производить хлорофилл, «зелень» в листьях, а по мере уменьшения содержания хлорофилла листья меняют цвет.Красивое отображение ярко-красного цвета, желтые и золотые листья, связанные с лиственными лесами в падение, является результатом этого процесса. Большинство лиственных деревьев сбрасывают листья, как только листья станут коричневыми и сухими. Фотографии 2000-www.arttoday.com Назад | Следующий |
Типы лесных экосистем | Sciencing
По словам Элизабет Мигатт, автора статьи «Мировые леса продолжают сокращаться, примерно 30 процентов поверхности Земли покрыто различными типами лесов.Мигатт продолжает объяснять решающую роль, которую лесная экосистема играет в поддержании здоровья планеты, например, в управлении круговоротом воды и стабилизации почв, помощи в выравнивании климата путем насыщения и хранения углекислого газа, обеспечения среды обитания для диких животных и снабжения древесина, продукты питания и лекарства. По всему миру существует несколько различных видов лесов. Хрестоматийное определение экосистемы, как указано в «Элементных геосистемах», — это «саморегулирующееся объединение живых растений, животных и их неживых физических и химическая среда.»
Определение тропических лесов
В тропических лесах дни обычно длятся 12 часов со средней температурой около 77 градусов по Фаренгейту. Избыток дождя и высокая инсоляция (солнечный свет) являются другими круглогодичными атрибутами влажных тропических лесов. Тропические леса покрывают регион Амазонки, а также экваториальные регионы Африки, Юго-Восточной Азии, восточное побережье Центральной Америки и другие места вдоль экватора. Эти типы лесных экосистем характеризуются широколиственными вечнозелеными деревьями, виноградными лозами, древовидными папоротниками и пальмами.
Многие тропические леса известны своим невероятным биоразнообразием. Джунгли, такие как Амазонка, являются домом для тысяч различных видов животных. К ним относятся животные, которые ползают по земле, такие как насекомые, ящерицы и грызуны, а также те, которые качаются по деревьям, например обезьяны. В тропических лесах также обитают злобные хищники, такие как анаконды и ягуары.
Тропический сезонный лес
На окраине тропических лесов находятся тропические сезонные леса, которые получают нерегулярные осадки в уменьшающемся количестве.Некоторые из ключевых частей леса в этой зоне — это широколиственные вечнозеленые деревья, некоторые лиственные деревья и колючие деревья. Листопадные деревья теряют листья зимой.
Биом вечнозеленых и лиственных лесов умеренного пояса
Вечнозеленые и лиственные леса умеренного пояса, обитающие в Северной Америке, Европе и Азии, временами сливаются воедино. В лесах обитают игольчатые и широколиственные деревья. В южных и восточных районах, где растут вечнозеленые сосны, до сих пор происходят контролируемые лесные пожары как естественный цикл восстановления и обогащения леса.
Их называют лесами умеренного пояса из-за погодных условий. По сравнению с экстремальной жарой и влажностью тропических лесов, вечнозеленые и лиственные леса умеренного пояса имеют умеренный климат с приятно теплым летом и терпимо холодной зимой.
Северный лес
Северный лес является домом для многих растений и животных, которых обычно считают лесными обитателями, таких как:
- лисы
- лоси
- северный олень
- медведи
- белки
- волки
Такие растения, как грибы, мхи и лишайники, хорошо себя чувствуют в северном климате, потому что они достаточно выносливы, чтобы выжить в очень холодные ледяные зимы.
Саванна и лесной массив
Саванна и лесные экосистемы подвержены пожарам и обладают способностью омолаживаться и восстанавливаться. Саванны и лесные массивы, распространенные в Южной Америке, Африке и Австралии, характеризуются обширными лугами, зарослями кустарников и скоплениями редких деревьев с уплощенными кронами.