Смешанные леса плодородие. Почвы лесного пояса умеренных широт
Зона широколиственных лесов занимает широкую полосу в Евразии. Зональным типом почв в ней являются лесные почвы, которые распространены под широколиственными лесами в умеренно теплых и влажных приокеанических областях суббореального пояса в Западной, Средней Европе, на Дальнем Востоке, в приатлантической и береговой части Северной Америки. Особенно распространены эти почвы в Западной Европе.
Климат . Умеренно теплый с мягкой зимой и значительным количеством осадков (600–1000 мм). Коэффициент увлажнения больше единицы (1,1–1,3), водный режим промывной.
Рельеф . Равнинный.
Растительность . Широколиственные леса из бука, дуба, граба, ясеня, липы, клена, пихты, кедра и саянской ели. Леса светлые, разреженные, поэтому в них формируется густой травяной покров.
Почвообразующие породы – это преимущественно эллювиально-деллювиальные и аллювиальные отложения, лессы, лессовидные и покровные суглинки, породы, обогащенные карбонатами или силикатными основаниями.
Бурые лесные почвы (буроземы) . Формируются при сочетании процессов гумусонакопления, оглеения, лессиважа. Лиственные леса образуют богатый зольными элементами опад, который во влажных и теплых условиях поддается процессам гумификации и минерализации при активном участии большого количества микрофлоры и беспозвоночных. В результате глубокой переработки органического вещества образуется тонкий илистый (мулевый) гумус, так называемый «мягкий», в котором преобладают гуминовые кислоты. Гуминовые кислоты с оксидами железа образуют нерастворимые в воде соединения, которые оструктуривают почву (органожелезистые комплексы).
Кроме того, в буроземах проявляется процесс внутрипочвенного оглеения, т.е. обогащения иллювиального горизонта илистыми частицами (вторичными минералами), которые образуются из первичных в результате биохимических и химических процессов, а также синтезу из продуктов минерализации . Возможен также перенос илистых частиц в горизонт В сверху в условиях промывного режима путем лессиважа.
Физико-химические бурых лесных почв значительно варьируют в зависимости от направления почвообразования и состава пород. У большинства почв реакция среды слабокислая (рН 5,0– 6,5) и уменьшается с глубиной. Количество гумуса в горизонте А1 может быть 4–10%, емкость поглощения довольно высока (Е=30–35 мг/экв/100 г почвы), насыщенность основаниями высокая (V может быть до 80–90%). Худшие показатели имеют оподзоленные буроземы.
Бурые лесные более плодородные, чем . Используются они как пахотные земли, сенокосы, пастбища и лесные угодья. Основное мероприятие по повышению плодородия – создание окультуренного пахотного горизонта, внесение органических и минеральных удобрений, по мере необходимости проводят известкование. На бурых лесных почвах размещаются наиболее качественные лесопитомники.
Какие почвы преобладают в смешанных лесах, Вы узнаете из этой статьи.
Какие почвы в зоне смешанных лесов?* Дерново-подзолистые грунты
В смешанных лесах широко образовывается почва дерново-подзолистого типа. Благодаря погодным условиям здесь сформировался мощный гумусо-аккумулятивный горизонт, в котором небольшая доля принадлежит дерновому слою. В процессах образования дерново-подзолистых грунтов участвуют следующие элементы:
1. Зольные частицы
3. Кальций
7. Водород
8. Алюминий
Так как среда данного типа грунта окисленная, то его плодородность не слишком большая. В нем содержится 3-7% гумуса. Дерново-подзолистый грунт обогащен кремнеземом, но в то же время практически не содержит азота и фосфора. Содержит большое количество влаги.
В нем содержится 3-7% гумуса. Дерново-подзолистый грунт обогащен кремнеземом, но в то же время практически не содержит азота и фосфора. Содержит большое количество влаги.
* Лесные серые почвы
Серый грунт считается переходным грунтом от подзолистой почвы до черноземов. Данный тип сформировался благодаря теплому климату и растительной разнообразности. Основа образования серых грунтов – частички растений, экскременты животных, остатки микроорганизмов. Перемешавшись, они создают большой слой гумуса.
* Бурые почвы
Бурые грунты также формируются под влиянием теплого климата, а точнее умеренно-жаркого и постоянного увлажнения почвы. Имеют насыщенный коричневый оттенок. Благодаря тому, что на таких грунтах прорастает большое количество травы, они достаточно обогащены гумусом. Но его уровень плодородности несколько ниже, чем у чернозема за счет того, что повышенная влажность приводит к вымыванию некоторых элементов.
Что такое смешанный лес?Смешанные леса являют собой природную зону, где прорастают различные виды лиственных и хвойных деревьев.
В смешанных лесах встречаются разные виды деревьев. Основными породами являются — клен, дуб, липа, береза, граб, сосна, лиственница, пихта, ель. Благодаря высокому уровню влажности и частой смене сезонов погоды в этих зонах сформировались разные виды грунтов, а именно бурые, дерново-подзолистые и лесные серые грунты. Они характеризуются большим уровнем вместительности гумуса.
Записи на букву П
Пóчвы смéшанных и широколи́ственных лесóв .
В смешанных лесах умеренного пояса подзолистые почвы (см. Почвы тайги ) приобретают гумусовые почвенные горизонты. Прежде всего это объясняется тем, что здесь произрастает много травянистых растений, остатки которых перемешиваются почвообитающими животными (червями, кротами и др.) с минеральными веществами почвы. Такие почвы с гумусовым горизонтом, горизонтом вымывания железа и глинистых частиц, а также с бурым горизонтом вмывания называются дерново‑подзолистыми .
В подзоне смешанных лесов встречаются также переувлажнённые почвы заболоченных лугов с гумусовым и глеевым почвенными горизонтами — они называются дерново‑глеевыми почвами .
В широколиственных лесах умеренного пояса формируются серые лесные почвы и бурые лесные почвы, или бурозёмы.
Серые лесные почвы представляют собой переход между дерново‑подзолистыми почвами смешанных лесов и чернозёмами лесостепей и степей. Они формируются в более тёплом и сухом климате и под более обильной растительностью, чем дерново‑подзолистые почвы. Растительных остатков и перемешивающих их почвенных животных здесь больше, поэтому гумусовый горизонт в них глубже и темнее. Однако в связи с устойчивым снежным покровом каждую весну при таянии снега почва испытывает своеобразный удар — активно промывается, поэтому в ней формируются горизонты вымывания и вмывания.
Бурые лесные почвы — это почвы более тёплого, но не менее влажного климата, чем тот, в котором формируются дерново‑подзолистые почвы. Распространены в Зап. и Центр. Европе, на северо‑вост. побережье США, на самом Ю.
В таких условиях органические остатки разлагаются постепенно, формирующийся
В смешанных лесах растут различные деревья. Лесообразующими породами являются, как широколиственные (клены, дубы, липы, березы, грабы), так и хвойные (сосны, лиственницы, пихты, ели). В таких природных зонах формируются дерново-подзолистые, бурые и серые лесные грунты. В них отмечается достаточно большой уровень содержания гумуса, что обусловлено произрастанием в этих лесах большого количества трав. Из них вымывается железо и глинистые частицы.
Дерново-подзолистые грунты
В хвойно-широколиственных лесах широко образовывается земля дерново-подзолистого типа. В условиях леса формируется значительный гумусо-аккумулятивный горизонт, а дерновый слой проявляет небольшую мощность. В процессе почвообразования участвуют зольные частицы и азот, магний и кальций, железо и калий, алюминий и водород, а также другие элементы. Уровень плодородия такого грунта не высокий, поскольку среда является окисленной. Дерново-подзолистая земля содержит от 3 до 7% гумуса. Еще она обогащена кремнеземом, и бедна на фосфор и азот. Этот тип грунта обладает большой влагоемкостью.
Уровень плодородия такого грунта не высокий, поскольку среда является окисленной. Дерново-подзолистая земля содержит от 3 до 7% гумуса. Еще она обогащена кремнеземом, и бедна на фосфор и азот. Этот тип грунта обладает большой влагоемкостью.
Серые почвы и буроземы
Бурые и серые грунты образовываются в лесах, где произрастают одновременно хвойные и широколиственные деревья. Серый тип является переходным между подзолистыми грунтами и черноземами. Серые почвы формируются в условиях теплого климата и растительного разнообразия. Это способствует тому, что растительные частички, экскременты животных вследствие деятельности микроорганизмов перемешиваются, и появляется большой и обогащенный различными элементами гумусовый слой. Он глубже залегает и имеет темную окраску. Однако почва каждую весну, когда тает снег, испытывает значительное увлажнение и вымывание.
Лесные бурые грунты формируются еще в более теплом климате, чем лесные. Для их образование лето должно быть умеренно-жарким, а зимой не должно быть постоянного снежного слоя. В течение круглого года увлажнение земли происходит равномерно. При таких условиях гумус становится бурого коричневого оттенка.
В течение круглого года увлажнение земли происходит равномерно. При таких условиях гумус становится бурого коричневого оттенка.
В смешанных лесах можно встретить разнообразные типы грунтов: буроземы, серые лесные и дерново-подзолистые. Условия их образования приблизительно одинаковые. Наличие густой травы и лесной подстилки способствует тому, что почва обогащается гумусом, а вот повышенная влажность способствует вымыванию различных элементов, что несколько понижает плодородие грунта.
Макро- и микроэлементы.
Непрерывное использование земель носит негативный характер. Начиная с восьмидесятых годов прошлого столетия стали непригодными 10 млн. га пашен. Большая часть почв России была закислена, засолена, переувлажнена, а также подверглась химическому и радиоактивному загрязнению. Отрицательно на плодородии почв сказывается ветровая и водная эрозия.
Типы и карта почв России
Огромная протяженность, разнообразие климата, рельефа и водного режима сформировали пестрый почвенный покров. Каждому региону соответствует свой тип почв. Важнейшим показателем плодородия является толщина гумусного горизонта. Гумусом называют верхний плодородный слой грунта. Он образуется благодаря деятельности микроорганизмов, которые перерабатывают остатки растительного и животного происхождения.
Каждому региону соответствует свой тип почв. Важнейшим показателем плодородия является толщина гумусного горизонта. Гумусом называют верхний плодородный слой грунта. Он образуется благодаря деятельности микроорганизмов, которые перерабатывают остатки растительного и животного происхождения.
На территории России наиболее распространены следующие типы почв:
Арктические почвы
Арктические почвы находятся на Северного Ледовитого . Они практически не содержат гумуса, почвообразовательные процессы на низком уровне из-за . Арктические районы используются в качестве охотничьих угодий или сохранения популяций уникальных видов животных.
Тундровые почвы
Тундровые почвы расположены в и вдоль побережье морей Северного Ледовитого океана. В этих районах господствует вечная мерзлота. Лишайники и мхи, образовавшиеся в летний период, не являются хорошим источником для формирования гумуса. Из-за многолетней мерзлоты почва за короткое лето оттаивает лишь на 40 см вглубь.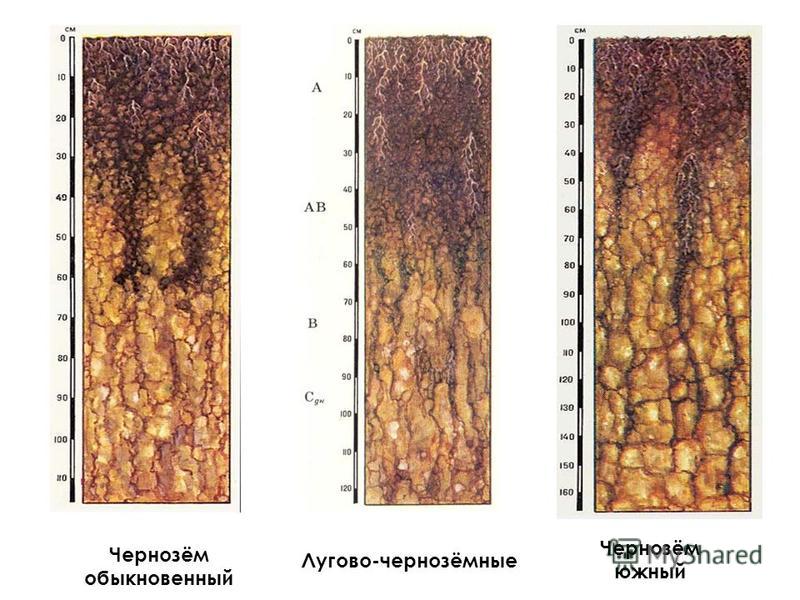 Земли зачастую засолены. Содержание гумуса в почве тундровой зоны незначительно из-за слабой микробиологической активности. Земли используются местными жителями в качестве пастбищ для оленей.
Земли зачастую засолены. Содержание гумуса в почве тундровой зоны незначительно из-за слабой микробиологической активности. Земли используются местными жителями в качестве пастбищ для оленей.
Подзолистые почвы
Подзолистые почвы распространены в и смешанных лесах. Территории занимают 75% от общей площади России. Обилие воды и прохладный климат создают кислую среду. Из-за нее органические вещества уходят на глубину. Гумусовый горизонт не превышает десяти сантиметров. В почве мало питательных веществ, но много влаги. При правильной обработке она пригодна для сельского хозяйства. На обогащённых удобрениями подзолистых почвах дают хороший урожай злаковые, картофель и зерновые.
Серые лесные почвы
Серые лесные почвы располагаются в Восточной Сибири, ее лесостепях и широколиственных лесах. На формирование флоры региона влияет умеренный климат и рельеф. Земли представляют собой сочетание подзолистых и черноземных почв. Обилие растительных остатков, летние дожди и полное их испарение способствует накоплению перегноя. Леса богаты землями с углекислым кальцием. Благодаря высокому плодородию 40% серых лесных почв активно используются для нужд сельского хозяйства. Десятая часть приходится на пастбища и сенокосы. На остальных землях выращивают кукурузу, свеклу, гречиху и озимые культуры.
Леса богаты землями с углекислым кальцием. Благодаря высокому плодородию 40% серых лесных почв активно используются для нужд сельского хозяйства. Десятая часть приходится на пастбища и сенокосы. На остальных землях выращивают кукурузу, свеклу, гречиху и озимые культуры.
Черноземные почвы
Черноземные почвы находятся на юге страны, у границ с Украиной и Казахстаном. На толстый гумусный слой повлиял равнинный рельеф, теплый климат и незначительные осадки. Такой тип почв считается самым плодородным во всем мире. России принадлежит около 50% от мировых запасов черноземов. Большое количество кальция препятствует вымыванию полезных веществ. В южных районах наблюдается недостаток влаги. Земли возделывают сотни лет, но до сих пор они остаются плодородными. Больше остальных культур черноземы засевают пшеницей. Высокий урожай дает сахарная свекла, кукуруза и подсолнечник.
Каштановые почвы
Каштановые почвы преобладают в Астраханской области, Минусинских и Приамурских степях. Здесь наблюдается нехватка гумуса из-за высоких температур и недостатка влаги. Земля плотная, набухает при увлажнении. Соли плохо вымываются водой, почва имеет слабокислую реакцию. Она пригодна для земледелия, если поддерживать регулярное орошение. Здесь выращивают люцерну, хлопчатник, пшеницу и подсолнечник.
Здесь наблюдается нехватка гумуса из-за высоких температур и недостатка влаги. Земля плотная, набухает при увлажнении. Соли плохо вымываются водой, почва имеет слабокислую реакцию. Она пригодна для земледелия, если поддерживать регулярное орошение. Здесь выращивают люцерну, хлопчатник, пшеницу и подсолнечник.
Бурые и серо-бурые почвы
Бурые и серо-бурые почвы встречаются на Прикаспийской низменности. Их характерным признаком является пористая корка на поверхности. Она образуется из-за высоких температур и слабой увлажненности. Гумуса здесь незначительное количество. В почве накапливаются карбонаты, соли и гипс. Плодородие земель низкое, большинство территорий используется под пастбища. На орошаемых участках выращивают рис, хлопчатник и бахчевые культуры.
Почвы природных зон России
Карта природных зон России
Природные комплексы сменяют друг друга с севера на юг страны, всего их насчитывается восемь. Каждая природная зона России, характеризуется своим уникальным почвенным покровом.
Почвы арктической пустыни
Почвенный покров в практически не выражен. На небольших участках произрастают мхи и лишайники. В теплое время над землей появляется трава. Все это выглядит, как небольшие оазисы. Растительные остатки не могут сформировать гумус. Талый слой земли летом не превышает 40 см. Переувлажнение, а также летнее высушивание приводят к растрескиванию поверхности земли. В почве много железа, из-за чего она имеет бурый окрас. В арктической пустыне практически нет болот, озер, в сухую погоду на поверхности образуются солевые пятна.
Почвы тундры
В почвы переувлажненные. Это объясняется близким залеганием вечной мерзлоты и недостаточным испарением влаги. Темпы гумификации очень замедлены. Растительные остатки не могут перегнить и остаются на поверхности в виде торфа. Количество питательных веществ минимально. Земля имеет сизоватый или ржавый окрас.
Почвы лесотундры
Лесотундра характеризуется переходом от тундровых почв к таежным. Редколесья уже напоминают лес, у них поверхностная корневая система. Вечная мерзлота начинается на уровне 20 см. Верхний слой хорошо прогревается летом, что способствует образованию пышной растительности. Влага плохо испаряется из-за низких температур, поэтому поверхность заболочена. Участки лесотундры представляют собой сочетание подзолистых и торфяно-глеевых почв. Здесь мало гумуса, земли закислены.
Вечная мерзлота начинается на уровне 20 см. Верхний слой хорошо прогревается летом, что способствует образованию пышной растительности. Влага плохо испаряется из-за низких температур, поэтому поверхность заболочена. Участки лесотундры представляют собой сочетание подзолистых и торфяно-глеевых почв. Здесь мало гумуса, земли закислены.
Почвы тайги
В практически отсутствует зона мерзлоты, поэтому почвы подзолистые. Железо разрушается под действием кислот и вымывается в глубокие слои грунта. В верхних слоях образуется кремнезем. В тайге слабо развит подлесок. Опавшая хвоя и мох долго разлагаются. Содержание гумуса минимальное.
Почвы широколиственных и смешанных лесов
В широколиственных и смешанных лесах преобладают дерново-подзолистые и бурые почвы. Эта природная зона является домом для дубов, лиственниц, кленов, берез и лип. Опад деревьев формирует много гумуса. Слой дерна сокращает мощность земли, поэтому дерново-подзолистая почва бедна на фосфор и азот. Бурые почвы обогащены питательными элементами.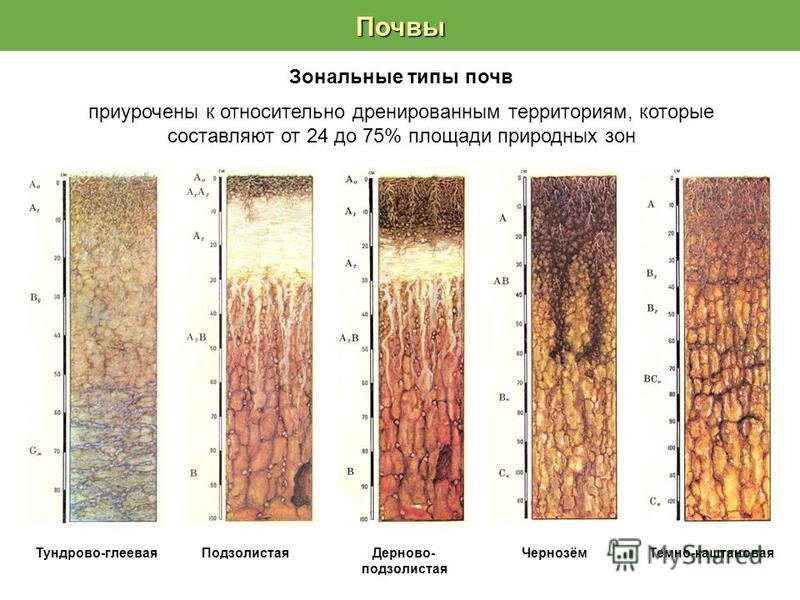 Гумус придает им темную окраску.
Гумус придает им темную окраску.
Почвы лесостепи
Лесостепи характеризуются высоким испарением влаги, в летний период наблюдается засуха и суховеи. В этой природной зоне формируются черноземные и серые лесные почвы. Слой гумуса большой, при этом минерализация замедленна. Благодаря особому плодородию земли лесостепи активно культивируются много лет подряд. Распаханные территории подвергаются выветриванию и высыханию.
Почвы степи
Представлена темно-каштановыми, обыкновенными и малогумусными черноземами. В почве достаточное количество питательных веществ. Гумуса в каштановых почвах меньше, поэтому они светлее остальных.
Почвы пустынь и полупустынь
В преобладают каштановые почвы. Из-за недостаточного увлажнения происходит накопление солей. Растительность не образует сплошного покрова. У растений глубокие корни, способные добывать влагу далеко от поверхности. Местами встречаются солончаки. Гумуса мало, в нижних слоях можно обнаружить гипс.
В каком регионе России почвы наиболее плодородны?
Чернозем — это самый плодородный тип почвы. Его невозможно сформировать искусственно. Чернозем занимает лишь 10% от общей территории страны, но его урожайность значительно превосходит другие почвы. Этот тип богат гумусом и кальцием. По структуре почва тяжелая, рыхлая, пористая, поэтому вода и воздух легко проникают к корням растений. Чернозем встречается в Центрально-Черноземном экономическом районе, включающем в себя Воронежскую, Курскую, Белгородскую, Липецкую и Тамбовскую области. Подзолистые почвы при правильной агротехнике также дают высокий урожай. Они распространены в европейской части России, на Дальнем Востоке и в Восточной Сибири.
Его невозможно сформировать искусственно. Чернозем занимает лишь 10% от общей территории страны, но его урожайность значительно превосходит другие почвы. Этот тип богат гумусом и кальцием. По структуре почва тяжелая, рыхлая, пористая, поэтому вода и воздух легко проникают к корням растений. Чернозем встречается в Центрально-Черноземном экономическом районе, включающем в себя Воронежскую, Курскую, Белгородскую, Липецкую и Тамбовскую области. Подзолистые почвы при правильной агротехнике также дают высокий урожай. Они распространены в европейской части России, на Дальнем Востоке и в Восточной Сибири.
Смешанные леса России. Растения и животные смешанного леса. Почвы смешанных лесов
Широколиственные и смешанные леса составляют значительно меньший процент лесной зоны России, чем хвойная тайга. В Сибири они отсутствуют совсем. Широколиственные и смешанные лесные массивы характерны для европейской части и дальневосточного региона РФ. Их образуют лиственные и хвойные породы деревьев. Они имеют не только смешанный состав древостоев, но и отличаются многообразием животного мира, устойчивостью к негативным воздействиям окружающей среды, мозаичностью строения.
Они имеют не только смешанный состав древостоев, но и отличаются многообразием животного мира, устойчивостью к негативным воздействиям окружающей среды, мозаичностью строения.
Типы и ярусность смешанных лесов
Существуют хвойно-мелколиственные и смешанно-широколиственные леса. Первые растут в основном в континентальных районах. Смешанные леса имеют хорошо заметную ярусность (изменение состава флоры, в зависимости от высоты). Самый верхний ярус — это высокие ели, сосны, дубы. Несколько ниже растут березы, клёны, вязы, липы, дикие груши и яблони, более молодой дубняк и другие. Далее идут более низкие деревья: рябина, калина и т. п. Следующий ярус образуют кустарники: калина, лещина, боярышник, заросли шиповника, малины и многие другие. Далее идут полукустарники. В самом низу растут травы, лишайники и мхи.
Промежуточные и коренные формы хвойно-мелколиственного леса
Интересной особенностью является то, что смешанно-мелколиственные массивы считаются лишь промежуточной стадией формирования хвойного леса. Однако они бывают и коренными: массивы из каменной березы (Камчатка), березовые колки в лесостепях, осиновые кустарники и заболоченные ольшаники (юг европейской части РФ). Леса мелколиственного типа очень светлые. Это способствует буйному росту травяного покрова и его разнообразию. Хвойно-смешанный лес широколиственного типа, напротив, относится к устойчивым природным формированиям. Он распространен в переходной полосе между таежными и широколиственными типами. Хвойно-широколиственные леса растут на равнинах и на самом нижнем горном поясе с умеренными и влажными климатическими условиями.
Однако они бывают и коренными: массивы из каменной березы (Камчатка), березовые колки в лесостепях, осиновые кустарники и заболоченные ольшаники (юг европейской части РФ). Леса мелколиственного типа очень светлые. Это способствует буйному росту травяного покрова и его разнообразию. Хвойно-смешанный лес широколиственного типа, напротив, относится к устойчивым природным формированиям. Он распространен в переходной полосе между таежными и широколиственными типами. Хвойно-широколиственные леса растут на равнинах и на самом нижнем горном поясе с умеренными и влажными климатическими условиями.
Зона смешанных и широколиственных лесов
Хвойно-широколиственные леса растут в более теплых регионах умеренного пояса. Они отличаются многообразием и богатством травяного покрова. Растут прерывистыми полосами от европейской части РФ до Дальнего Востока. Их ландшафты благоприятны для людей. К югу от тайги расположена зона смешанных лесов. Они распространены на всей площади Восточно-Европейской равнины, а также за Уралом (вплоть до Приамурья). Сплошную зону не образуют.
Сплошную зону не образуют.
Примерная граница европейского участка широколиственных и смешанных лесов на севере пролегает по 57° с. ш. Выше неё практически полностью исчезает дуб (одно из ключевых деревьев). Южная почти соприкасается с северной границей лесостепей, где совершенно исчезает ель. Данная зона представляет собой участок в форме треугольника, две вершины которого находятся в России (Екатеринбург, Санкт-Петербург), а третья — в Украине (Киев). То есть по мере удаления от основной зоны на север широколиственные, а также смешанные леса постепенно уходят с водораздельных пространств. Они предпочитают более теплые и защищенные от ледяных ветров речные долины с выходом на поверхность карбонатных пород. По ним леса широколиственного и смешанного типа небольшими массивами постепенно доходят до тайги.
Восточно-Европейская равнина в основном имеет низменный и плоский рельеф, лишь иногда встречаются возвышенности. Здесь расположены истоки, бассейны и водоразделы самых крупных российских рек: Днепр, Волга, Западная Двина. На их поймах луга перемежаются с лесами и пашнями. В некоторых регионах низины, из-за близкого расположения грунтовых вод, а также ограниченности стока, местами крайне заболочены. Встречаются также участки с песчаными грунтами, на которых растут сосняки. На болотах и вырубках растут ягодные кустарники и травы. Данная местность является наиболее подходящей для хвойно-лиственных лесов.
На их поймах луга перемежаются с лесами и пашнями. В некоторых регионах низины, из-за близкого расположения грунтовых вод, а также ограниченности стока, местами крайне заболочены. Встречаются также участки с песчаными грунтами, на которых растут сосняки. На болотах и вырубках растут ягодные кустарники и травы. Данная местность является наиболее подходящей для хвойно-лиственных лесов.
Влияние человека
Широколиственные, а также смешанные леса длительно подвергаются различным воздействиям со стороны людей. Поэтому многие массивы сильно изменились: коренная растительность либо совсем уничтожена, либо частично или полностью заменена вторичными породами. Сейчас остатки лесов широколиственного типа, которые уцелели под жестким антропогенным прессом, имеют различную структуру изменений флоры. Некоторые виды, утратив свое место в коренных сообществах, произрастают в антропогенно нарушенных местах обитания или заняли интразональные позиции.
Климат
Климат смешанных лесов довольно мягкий. Он характеризуется относительно теплой зимой (в среднем от 0 до –16°) и продолжительным летом (16-24 °С) по сравнению с таежной зоной. Среднегодовое количество осадков — 500-1000 мм. Оно повсеместно превышает испарение, что является особенностью явно выраженного промывного водного режима. Смешанные леса имеют такую характерную черту, как высокий уровень развития травяного покрова. Их биомасса в среднем составляет 2-3 тысячи ц/га. Уровень опада также превышает биомассу тайги, однако, благодаря более высокой активности микроорганизмов, разрушение органических веществ идёт значительно быстрее. Поэтому смешанные леса имеют меньшую толщину и больший уровень разложения подстилки, чем таежные хвойные.
Он характеризуется относительно теплой зимой (в среднем от 0 до –16°) и продолжительным летом (16-24 °С) по сравнению с таежной зоной. Среднегодовое количество осадков — 500-1000 мм. Оно повсеместно превышает испарение, что является особенностью явно выраженного промывного водного режима. Смешанные леса имеют такую характерную черту, как высокий уровень развития травяного покрова. Их биомасса в среднем составляет 2-3 тысячи ц/га. Уровень опада также превышает биомассу тайги, однако, благодаря более высокой активности микроорганизмов, разрушение органических веществ идёт значительно быстрее. Поэтому смешанные леса имеют меньшую толщину и больший уровень разложения подстилки, чем таежные хвойные.
Грунты смешанных лесов
Почвы смешанных лесов отличаются разнообразием. Покров имеет довольно пеструю структуру. На территории Восточно-Европейской равнины наиболее распространенным типом является дерново-подзолистый грунт. Он представляет собой южную разновидность классических подзолистых почв и формируется лишь при наличии почвообразующих пород суглинистого типа. Дерново-подзолистый грунт имеет такое же строение профиля и схожую структуру. Отличается он от подзолистого меньшей массивностью подстилки (до 5 см), а также более значительной мощностью всех горизонтов. И это не единственные различия. Дерново-подзолистые грунты имеют более выраженный гумусовый горизонт А1, который находится под подстилкой. Внешний вид его отличается от аналогичного слоя подзолистых почв. Верхняя часть содержит корневища травяного покрова и образует дернину. Горизонт может быть окрашен в различные оттенки серого цвета и имеет рыхлую структуру. Мощность слоя — 5-20 см, доля гумуса — до 4%. Верхняя часть профиля данных грунтов имеет кислую реакцию. По мере углубления она становится еще меньшей.
Дерново-подзолистый грунт имеет такое же строение профиля и схожую структуру. Отличается он от подзолистого меньшей массивностью подстилки (до 5 см), а также более значительной мощностью всех горизонтов. И это не единственные различия. Дерново-подзолистые грунты имеют более выраженный гумусовый горизонт А1, который находится под подстилкой. Внешний вид его отличается от аналогичного слоя подзолистых почв. Верхняя часть содержит корневища травяного покрова и образует дернину. Горизонт может быть окрашен в различные оттенки серого цвета и имеет рыхлую структуру. Мощность слоя — 5-20 см, доля гумуса — до 4%. Верхняя часть профиля данных грунтов имеет кислую реакцию. По мере углубления она становится еще меньшей.
Почвы смешано-широколиственных лесов
Серые лесные грунты смешанно-широколиственных лесов сформированы во внутриконтинентальных регионах. В России они распространены от европейской части до Забайкалья. В такие почвы осадки проникают на большую глубину. Однако горизонты грунтовых вод нередко находятся очень глубоко. Поэтому промачивание грунта до их уровня характерно лишь в сильно увлажняемых районах.
Поэтому промачивание грунта до их уровня характерно лишь в сильно увлажняемых районах.
Грунты смешанных лесов лучше подходят для земледелия, чем субстраты таежных. В южных регионах европейской части РФ пашни составляют до 45% площади. Ближе к северу и тайге доля пахотных земель постепенно уменьшается. Земледелие в этих регионах затруднено из-за сильной выщелоченности, заболоченности и завалуненности грунтов. Для получения хороших урожаев требуется много удобрений.
Общая характеристика фауны и флоры
Растения и животные смешанного леса очень разнообразны. По видовому богатству флоры и фауны они сравнимы лишь с тропическими джунглями и являются домом для многих хищников и травоядных. Здесь на высоких деревьях обживаются белки и другая живность, на кронах делают гнезда птицы, у корней обустраивают норы зайцы и лисы, а неподалеку от рек живут бобры. Видовое многообразие смешанной зоны очень велико. Тут комфортно себя чувствуют как жители тайги и широколиственных лесов, так и обитатели лесостепей. Одни бодрствуют круглый год, а другие на зиму впадают в спячку. Растения и животные смешанного леса имеют симбиотическую связь. Многие травоядные питаются разными ягодами, которых в смешанных лесах очень много.
Одни бодрствуют круглый год, а другие на зиму впадают в спячку. Растения и животные смешанного леса имеют симбиотическую связь. Многие травоядные питаются разными ягодами, которых в смешанных лесах очень много.
Деревья смешанного леса
Смешанно-мелколиственные лесные массивы примерно на 90% состоят из пород деревьев хвойного и мелколиственного типа. Широколиственных разновидностей не так много. Вместе с хвойными деревьями в них растут осины, березы, ольхи, ивы, тополя. Березняков в составе массивов данного типа больше всего. Как правило, они являются вторичными — то есть вырастают на лесных пожарищах, на вырубках и просеках, старых неиспользуемых пашнях. На открытых местообитаниях такие леса хорошо возобновляются и в первые годы быстро растут. Расширению их площадей способствует хозяйственная деятельность человека.
Хвойно-широколиственные леса преимущественно состоят из елей, лип, сосен, дубов, вязов, ильмов, кленов, а в юго-западных районах РФ – бука, ясеня и граба. Такие же деревья, но местных разновидностей, растут в дальневосточном регионе вместе с виноградом, маньчжурским орехом и лианами. Во многом состав и структура древостоя лесов хвойно-широколиственного типа зависит от климатических условий, рельефа и почвенно-гидрологического режима конкретного региона. На Северном Кавказе преобладают дуб, ель, клен, пихта и другие породы. Но наиболее разнообразными по составу являются дальневосточные леса хвойно-широколиственного типа. Они образованы кедровой сосной, белокорой пихтой, айянской елью, несколькими разновидностями клёна, маньчжурским ясенем, монгольским дубом, амурской липой и вышеупомянутыми местными видами растительности.
Такие же деревья, но местных разновидностей, растут в дальневосточном регионе вместе с виноградом, маньчжурским орехом и лианами. Во многом состав и структура древостоя лесов хвойно-широколиственного типа зависит от климатических условий, рельефа и почвенно-гидрологического режима конкретного региона. На Северном Кавказе преобладают дуб, ель, клен, пихта и другие породы. Но наиболее разнообразными по составу являются дальневосточные леса хвойно-широколиственного типа. Они образованы кедровой сосной, белокорой пихтой, айянской елью, несколькими разновидностями клёна, маньчжурским ясенем, монгольским дубом, амурской липой и вышеупомянутыми местными видами растительности.
Видовое разнообразие животного мира
Из крупных травоядных животных в смешанных лесах обитают лоси, зубры, кабаны, косули и пятнистые олени (вид завезен и адаптирован). Из грызунов присутствуют лесные белки, куницы, горностаи, бобры, бурундуки, выдры, мыши, барсуки, норки, черные хорьки. Смешанные леса изобилуют большим количеством видов птиц. Далее перечислены многие из них, но далеко не все: иволга, поползень, чиж, дрозд-рябинник, ястреб-тетеревятник, рябчик, снегирь, соловей, кукушка, удод, серый журавль, щегол, дятел, тетерев, зяблик. Более или менее крупные хищники представлены волками, рысями и лисами. Смешанные леса также являются домом для зайцев (русаков и беляков), ящериц, ежей, змей, лягушек и бурых медведей.
Далее перечислены многие из них, но далеко не все: иволга, поползень, чиж, дрозд-рябинник, ястреб-тетеревятник, рябчик, снегирь, соловей, кукушка, удод, серый журавль, щегол, дятел, тетерев, зяблик. Более или менее крупные хищники представлены волками, рысями и лисами. Смешанные леса также являются домом для зайцев (русаков и беляков), ящериц, ежей, змей, лягушек и бурых медведей.
Грибы и ягоды
Ягоды представлены черникой, малиной, брусникой, клюквой, ежевикой, черемухой, земляникой, костяникой, бузиной, рябиной, калиной, шиповником, боярышником. В лесах данного типа очень много съедобных грибов: подосиновики, белые, валуи, лисички, сыроежки, опята, грузди, маслята, волнушки, различные рядовки, подберезовики, моховики, рыжики и другие. Одними из самых опасных ядовитых макромицетов являются мухоморы и бледные поганки.
Кустарники
Смешанные леса России изобилуют кустарниками. Ярус подлеска необычайно развит. Для дубовых массивов характерно наличие лещины, бересклетов, волчьего лыка, жимолости лесной, а в северной зоне – крушины ломкой. На опушках и в редколесье растет шиповник. В лесах хвойно-широколиственного типа встречаются и лианообразные растения: повой заборный, хмель вьющийся, паслен сладко-горький.
На опушках и в редколесье растет шиповник. В лесах хвойно-широколиственного типа встречаются и лианообразные растения: повой заборный, хмель вьющийся, паслен сладко-горький.
Травы
Большое видовое разнообразие, а также сложную вертикальную структуру имеют травы смешанных лесов (особенно хвойно-широколиственного типа). Наиболее типичная и широко представленная категория — это мезофильные неморальные растения. Среди них выделяются представители дубравного широкотравья. Это растения, у которых листовая пластина имеет значительную ширину. К ним относятся: пролесник многолетний, сныть обыкновенная, медуница неясная, ландыш майский, копытень европейский, осока волосистая, зеленчук желтый, звездчатка ланцетовидная, сочевник (черный и весенний), фиалка удивительная. Злаковые представлены мятликом дубравным, овсяницей гигантской, вейником лесным, коротконожкой перистой, бором развесистым и некоторыми другими. Плоские листья данных растений являются вариантом приспособления к специфической фитосреде хвойно-широколиственных лесов.
Помимо вышеперечисленных многолетних видов в данных массивах встречаются также травы эфемероидной группы. Свой вегетационный период они переносят на весеннее время, когда освещение максимальное. После того как сходит снег, именно эфемероиды образуют красиво цветущий ковер из желтых ветрениц и гусиных луков, фиолетовых хохлаток и сиренево-голубоватых пролесок. Данные растения проходят жизненный цикл за пару-тройку недель, а когда распускаются листья деревьев, их надземная часть со временем отмирает. Неблагоприятный период они переживают под слоем грунта в виде клубней, луковиц и корневищ.
Природная зона смешанных и широколиственных лесов
ECOportal.info предлагает узнать растительный мир смешанных лесов России, Евразии и Америки. Кустарники, грибы и основные деревья смешанных лесов. Виды растений.
Содержание
Особенности
Хвойно-лиственные леса, расположенные в районе умеренного пояса, характеризуются подзолистыми почвами с выраженными гумусовыми горизонтами. Это получается по той причине, что здесь встречается большое количество трав. Опад перемешивают с минеральными элементами животные и живые организмы, обитающие в почве, к примеру, землеройки и черви. Так появляются дерново-подзолистые почвы, имеющие гумусовый горизонт, бурый горизонт вмывания, а также горизонт вымывания глинистых элементов и железа.
Это получается по той причине, что здесь встречается большое количество трав. Опад перемешивают с минеральными элементами животные и живые организмы, обитающие в почве, к примеру, землеройки и черви. Так появляются дерново-подзолистые почвы, имеющие гумусовый горизонт, бурый горизонт вмывания, а также горизонт вымывания глинистых элементов и железа.
В подзоне смешанных лесов зачастую можно встретить дерново-глеевые почвы. Они имеют глеевые и гумусовые почвенные горизонты, располагаются в районах заболоченных лугов, в основном являются переувлажненными.
Особенно часто такие почвы встречаются на европейской территории нашей страны, имея при этом широкую зону распространения.
Широколиственные леса расположены на территории умеренного пояса. Для них характерны как серые, так и бурые лесные почвы, известные под названием «буроземы». В первом случае почва является переходом от черноземов степных районов к дерново-подзолистым почвам, характерным для смешанных лесов. Территориально они размещаются там, где климат является относительно теплым и минимально влажным, а растительность – максимально обильной.
Территориально они размещаются там, где климат является относительно теплым и минимально влажным, а растительность – максимально обильной.
На такой местности образуется гораздо больше растительных остатков, а также увеличивается количество живых организмов, обитающих в почве. По этой причине гумусовый горизонт оказывается темнее и глубже. Но так как снежный покров в таких случаях образуется очень устойчивый, возникают и определенные проблемы. В весенний период таяние снега становится серьезным испытанием для почвы (она начинает промываться).
В значительно более теплом и влажном климате располагаются бурые лесные почвы. Они особенно часто встречаются в центральной и западной части Европы, на северо-востоке Соединенных Штатов, на юге дальневосточного побережья и в Японии. Эти регионы отличаются довольно устойчивым климатом, здесь могут встречаться муссонные ветра, но при этом нет жаркого и засушливого лета, чередующегося с появлением обильного и стойкого снежного покрова в зимний период.
Поэтому такие почвы отличаются равномерным увлажнением на протяжении года, что позволяет органике разлагаться более медленно и равномерно. При этом гумус становится более темного бурого цвета, а железо вымывается менее активно.
Источник: http://stroy-podskazka.ru/pochva/shirokolistvennyh-i-smeshannyh-lesov/
География таежно-лесных почв
Таежно-лесные почвы занимают огромную часть России, около 1 155 млн га. Они расположились на территории от западной границы страны до Охотского и Японского морей.
Рельеф таежно-лесной зоны представлен чередованием равнин и гор. Встречаются холмы, понижения, равнины, бассейны рек, овраги. 65% всех таежно-лесных почв находятся на равнинах, а оставшиеся 35% – в гористой местности.
Обратите внимание на географическую карту, представленную ниже:
Территория, которую занимают таежно-лесные почвы, отмечена зеленым и салатовым цветами. Эта площадь огромна! Ни одна другая зона даже близко не охватывает столько районов и не представлена так же широко, как область тайги, смешанных и широколиственных лесов. Все это не может не сказаться на процессах почвообразования местных покровов. Об этом – в следующей части статьи.
Все это не может не сказаться на процессах почвообразования местных покровов. Об этом – в следующей части статьи.
Источник: http://gruntovozov.ru/chasto-zadavayemiye-voprosy/vidyi-pochv-klassifikatsiya/tipyi-pochv-zonalnaya-klassifikatsiya-po-prirodnyim-zonam/pochvyi-taygi-smeshannyih-i-shirokolistvennyih-lesov/
Географическое положение
Смешанные и широколиственные леса находятся к югу от тайги. В европейской части России представлены оба типа, а все то, что за Уралом, лишь смешанный лес. На Дальнем Востоке его выделяют особо, часто добавляя к названию слово «муссонный», что связано с климатом региона и, как следствие, иным видовым разнообразием зоны.
Источник: http://sitekid.ru/planeta_zemlya/rossiya/prirodnaya_zona_smeshannyh_i_shirokolistvennyh_lesov.html
Что такое зона смешанных лесовХарактерный признак лесов подобного типа – наличие в них и лиственных, и хвойных деревьев, причем доля последних не должна быть менее чем 5 %.
Зоны смешанных лесов можно найти практически на любом континенте, они характеризуются большим разнообразием флоры и фауны.
Источник: http://karatu.ru/zona-smeshannyx-lesov/
Особенности почвенных ресурсов России
Далеко не все земли страны пригодны для сельскохозяйственной деятельности, часть из них находится в арктическом и субарктическом поясе. Большинство земледельческих угодий расположено в Центрально-Черноземном районе. На Дальнем Востоке земледелие развито слабо. Пастбища и пашни занимают 13% от всей площади. На луга и сенокосы приходится 1% почв. Человеческой деятельностью не затронуто примерно 45% земельных ресурсов. Они приходятся на лесной фонд страны.
Непрерывное использование земель носит негативный характер. Начиная с восьмидесятых годов прошлого столетия стали непригодными 10 млн. га пашен. Большая часть почв России была закислена, засолена, переувлажнена, а также подверглась химическому и радиоактивному загрязнению. Отрицательно на плодородии почв сказывается ветровая и водная эрозия.
Источник: http://natworld.info/raznoe-o-prirode/pochvy-rossii-karta-tipy-opisanie-prirodnye-zony-i-plodorodie
Смешанные и широколиственные леса
Растения этих мест, как и животные, тоже весьма разнообразны. Помимо крупных деревьев и кустарников, встречаются и всевозможные цветы, травы, мхи. Грибы тоже можно отнести к растительной флоре. Приятно в середине лета или начале осени побродить по лесу и принести домой крепкие боровики, красноголовые подосиновики, стройные подберёзовики. Радуют своей пёстрой красотой лисички, рыжики. Эти грибы растут в почве, а вот опята выбирают для себя старые или погибшие деревья. В основном – берёзы.
Травяной покров этой зоны по большей части составляют дубравные растения: зеленчук, осока волосистая, копытня. Встречаются в переходных зонах и некоторые таежные: кислица, седмичник, грушанка, майник и другие.
Ягодных растений в этом природном оазисе тоже немало. Здесь можно набрать много земляники, костяники, черники, клюквы. Цветы — ромашки, лютики, ландыши и другие — радуют взоры.
Вот таковы животные и растения смешанных лесов России. Важно не забывать относиться к природе очень бережно и охранять ее от недобрых людей и пожаров.
Источник: autogear.ru
Источник: http://greenway-schop.ru/problemy/smeshannye-lesa-harakteristika.html
Климат
Более мягкий и влажный по сравнению с таежным. Зима не такая суровая, лето долгое и теплое. За год в зоне выпадает 600–800 мм осадков. Температура июля в среднем достигает +20 °С.
На европейскую часть зоны влияют теплые воздушные массы с Атлантики. В результате за год выпадает до 600– 700 мм осадков, которые более-менее равномерно распределены по всем месяцам.
В лесах зоны хорошо прослеживается сезонность
За Уралом континентальность климата усиливается: осадков становится меньше, летние температуры растут, зимние — понижаются.
На Дальнем Востоке, где условия диктует муссонный климат с ярко выраженной сезонностью, ситуация иная. Лето теплое и очень влажное, а зима практически без осадков.
Лето теплое и очень влажное, а зима практически без осадков.
Источник: http://sitekid.ru/planeta_zemlya/rossiya/prirodnaya_zona_smeshannyh_i_shirokolistvennyh_lesov.html
Влияние человека
Широколиственные, а также смешанные леса длительно подвергаются различным воздействиям со стороны людей. Поэтому многие массивы сильно изменились: коренная растительность либо совсем уничтожена, либо частично или полностью заменена вторичными породами. Сейчас остатки лесов широколиственного типа, которые уцелели под жестким антропогенным прессом, имеют различную структуру изменений флоры. Некоторые виды, утратив свое место в коренных сообществах, произрастают в антропогенно нарушенных местах обитания или заняли интразональные позиции.
Источник: http://sovetunion.ru/pochvy-ih-osnovnye-svoistva-smeshannyh-lesov-pochva-smeshannyh-lesov.html
↑ Характеристика смешанного леса
Лес считается смешанным, если в нём присутствует 5-7 % типов других деревьев. Эти леса устойчивы к неблагоприятному воздействию климата с тёплым летом и не слишком морозной зимой, которая длится 3,5-4 месяца.
Эти леса устойчивы к неблагоприятному воздействию климата с тёплым летом и не слишком морозной зимой, которая длится 3,5-4 месяца.
смешанный лес осенью фото
Близость полноводных рек даёт много влажности, а осадочные породы: глина и песок помогают образованию озёр и болот. Хвойно-мелколиственные леса, встречающиеся среди тайги, не долговечны. Здесь они играют роль восстановительной стадии для уничтоженного хвойного бора или широколиственной дубравы.
Источник: http://greenway-schop.ru/problemy/smeshannye-lesa-harakteristika.html
Растительность
В зонах широколиственных и смешанных лесов произрастают самые разные деревья и кустарники. Здесь можно встретить осину, березу, липу, ель, бузину, малину, калину и многие другие. Климат довольно мягкий, что позволяет активно развиваться как лиственным, так и хвойным деревьям. На самом юге хвойных деревьев практически нет, встречаются исключительно сосны. Здесь широко распространены широколиственные породы, например, дуб и клен.
Могут похвастать леса и изобилием трав. В сырых и затененных местах встречается мох. Нужно отметить, что подобные леса очень плодородны. Здесь огромное количество грибов и ягод. Часто растут земляника, черника и другие лесные ягоды.
Источник: http://stroy-podskazka.ru/pochva/shirokolistvennyh-i-smeshannyh-lesov/
Растения
В смешанных лесах Восточно-Европейской равнины распространены дуб, клен, липа, ясень и орешник. Из хвойных преобладают ель и сосна. В широколиственных хвойные породы отсутствуют.
За Уралом широколиственных лесов нет вообще. Причина кроется в климатических условиях. Дуб, липа, клен, ясень, вяз пришли на территорию России из Западной Европы с ее мягким климатом. Ни одна из перечисленных древесных пород не смогла проникнуть за Урал из-за нарастающей там континентальности и неподходящих для роста условий. Из лиственных пород, которые присутствуют в составе смешанного леса, за Уралом есть только мелколиственные — береза, осина, ольха.
Особенность смешанных лесов — многоярусность. Нижний ярус — трава, средний — кустарники, верхний — древесные породы
Маньчжурский орех — очень морозостойкая культура, родина которой Дальний Восток. Листья и плоды (орехи) дерева обладают целебными свойствами
Муссонные леса
Они не похожи на европейские и различаются по составу. Из хвойных лесообразующими здесь являются корейский кедр, цельнолистная пихта, лиственница и аянская ель. Среди широколиственных чаще встречаются дуб монгольский, липа амурская, бархатное дерево, маньчжурский орех и др. Женьшень и водяной орех находятся на грани исчезновения.
Корейский кедр — один из наиболее распространенных представителей хвойных на Дальнем Востоке
Дальневосточные леса отличаются большим разнообразием древесных и кустарниковых пород, многоярусностью и обилием лиан
Источник: http://sitekid.ru/planeta_zemlya/rossiya/prirodnaya_zona_smeshannyh_i_shirokolistvennyh_lesov. html
html
Животные
В европейской части зоны встречаются лось, кабан, волк, выхухоль (очень древний и редкий здесь вид), куница и зубр (тоже редкость). Из птиц наиболее распространенными являются дубонос, иволга, дятел и др. В муссонных лесах обитают таежные и южные животные. Многие виды представителей фауны занесены в Красную книгу: уссурийский тигр, пятнистый олень, утка-мандарика и др.
Зубр — самое тяжелое и крупное млекопитающее в Европе. Раньше ареал его обитания был очень широк, но из-за интенсивной вырубки лесов и осушения болот, а также безжалостной охоты зверь оказался на грани исчезновения
Белка — наиболее распространенный представитель животного мира смешанных и широколиственных лесов в европейской части России
Источник: http://sitekid.ru/planeta_zemlya/rossiya/prirodnaya_zona_smeshannyh_i_shirokolistvennyh_lesov.html
Экологические проблемы
В данном вопросе следует выделить, что большинство негативных моментов провоцирует деятельность человека. Ряд экологических проблем отмечается во всех хвойно-лиственных и широколиственных лесах. К ним можно отнести погодные изменения, браконьерство, лесные пожары, мусор и вырубку лесов. Атмосфера, вода и почва планомерно загрязняются. Это оказывает влияние на погодные условия.
Ряд экологических проблем отмечается во всех хвойно-лиственных и широколиственных лесах. К ним можно отнести погодные изменения, браконьерство, лесные пожары, мусор и вырубку лесов. Атмосфера, вода и почва планомерно загрязняются. Это оказывает влияние на погодные условия.
В некоторых случаях отмечаются погодные явления, которые не характерны для определенного временного периода, особенно в последнее время. Это, конечно, влияет на флору и фауну. Например, аномальное понижение температуры способно разрушить древесную кору, а палящее солнце грозит пожарами, так как быстро испаряет влагу. Кислотный дождь способен замедлить рост растений вследствие воздействия на процесс фотосинтеза.
Браконьерство приводит к сокращению популяций определенных видов животных, в том числе до полного их вымирания. При этом нарушается пищевая цепочка, а за ней и жизнедеятельность целых экологических систем. Кроме того, возможна передача вирусов от человека к животному, а это, в свою очередь, грозит серьезными заболеваниями. Что касается лесных пожаров, то подавляющее их большинство происходит по вине людей. Крупные пожары способны уничтожить огромные территории растений, а также привести к гибели множества живых существ. Это также оказывает значительное влияние на экосистему.
Что касается лесных пожаров, то подавляющее их большинство происходит по вине людей. Крупные пожары способны уничтожить огромные территории растений, а также привести к гибели множества живых существ. Это также оказывает значительное влияние на экосистему.
Мусор появляется в лесах также по вине человека. Некоторые искусственные материалы (например, пластик и полиэтилен) не выводятся из живого организма, поэтому употребившее их по ошибке в пищу животное умирает от засорения органов пищеварения. Эти предметы очень долго разлагаются, отравляя окружающую среду, засоряя почву, оказывая неблагоприятное влияние на растительность лесов.
В наше время огромной проблемой является неорганизованная и бесконтрольная вырубка лесов. В результате уничтожаются огромные лесные массивы, не восполняясь молодняком. Со временем такое положение вещей способно привести к тому, что в атмосфере накопится чрезмерное количество углекислого газа. Всё это может привести к парниковому эффекту, вследствие которого разрушится почва, а многие представители животного и растительного мира просто исчезнут с нашей планеты.
Люди привыкли, что природа смешанных лесов соответствует тому, чтобы использовать ее для хозяйственных нужд. По этой причине вырубаются многие гектары, на месте которых создают поля, предназначенные для сельского хозяйства. Изменяются природные зоны, уничтожаются животные, распахиваются земли. Далеко не все виды живых организмов способны выжить в изменённых условиях.
Источник: http://stroy-podskazka.ru/pochva/shirokolistvennyh-i-smeshannyh-lesov/
Еще тесты
- Анатомия
- Английский язык
- Астрономия
- Биология
- Физика
- История
- Педсовет
- Естествознание
- Финансы и кредит
- Правоведение
- Товароведение
- Экономика
- Социология
- Маркетинг
- Обществознание
- Культурология
- Математика
- Философия
- Литература
- Психология
- Политология
- Делопроизводство
- Бухгалтерия
- ОБЖ
- Орфография
- География
- Биографии
- Пунктуация
- Краткие содержания
- Химия
- Русский язык
- Менеджмент
Источник: http://nauka. club/geografiya/smeshannye-i-shirokolistvennye-lesa-xarakteristika-prirodnoj-zony.html
club/geografiya/smeshannye-i-shirokolistvennye-lesa-xarakteristika-prirodnoj-zony.html
Человек
Зона смешанных и широколиственных лесов давно освоена людьми. Здесь добывают полезные ископаемые, вырубают зеленые массивы, эксплуатируют мощные энергетические запасы рек. Огромная часть территории отведена под земледелие и скотоводство. Кроме того, значительное пространство отдано под города, дороги, трубопроводы, водохранилища и др. Все это привело к тому, что зона широколиственных и смешанных лесов очень сильно изменилась. Сегодня здесь практически не осталось не тронутых человеком мест, а те, что сохранились, находятся под защитой.
Поделиться ссылкой
Источник: http://sitekid.ru/planeta_zemlya/rossiya/prirodnaya_zona_smeshannyh_i_shirokolistvennyh_lesov.html
Видео о широколиственных лесах
Документальный фильм про леса:
Источник: http://touristam. com/shirokolistvennye-lesa-rossii.html
com/shirokolistvennye-lesa-rossii.html
Смешанные леса Лёссового плато Центрального Китая
Территория экорегиона указана в единицах по 1000 га. Целью защиты является зона Глобальной сети безопасности (GSN1) для данного экорегиона. Уровень защиты указывает процент цели GSN, которая в настоящее время защищена, по шкале от 0 до 10. N/A означает, что в настоящее время данные недоступны.
Биорегион: Лёссовое плато и смешанные леса равнины Хуанхэ (PA49)
Область: Восточная Евразия
Ecoregion Size (1000 ha):
36,043
Ecoregion ID:
653
Protection Goal:
9%
Protection Level:
7
Государства: Китай
Лессовое плато Центрального Китая, расположенное к северо-западу от бассейна Хуанхэ, образует переходную зону между экорегионами Восточного Китая, находящимися под влиянием моря, и степями и пустынями Монголии. Это плато из уплотненной выветренной почвы (называемой лёссом) толщиной до 200 м, простирающееся с северо-востока на юго-запад более чем на 1000 км. Почвы этого уникального ландшафта были принесены из Центральной Азии ветрами, которые дуют сезонно с момента поднятия Тибетского нагорья и начала цикла муссонов более 20 миллионов лет назад.
Это плато из уплотненной выветренной почвы (называемой лёссом) толщиной до 200 м, простирающееся с северо-востока на юго-запад более чем на 1000 км. Почвы этого уникального ландшафта были принесены из Центральной Азии ветрами, которые дуют сезонно с момента поднятия Тибетского нагорья и начала цикла муссонов более 20 миллионов лет назад.
Ведущим видом экорегиона смешанных лесов Центрально-Китайского лессового плато является коричневый ушастый фазан. Изображение предоставлено: Creative Commons
Лессовые почвы богаты минералами, необходимыми для растений, и они достаточно хорошо удерживают воду, поэтому они потенциально продуктивны, несмотря на сезонно сухой климат, подверженный засухе. Поскольку Лёссовое плато хорошо подходит для сельского хозяйства, большая часть естественных лесов была преобразована в сельскохозяйственные угодья и пастбища столетия назад. Исторические записи свидетельствуют о том, что лесной покров на Лёссовом плато сократился в восемь раз за последние три тысячелетия и достиг своего пика в 1949. По мере вырубки лесов эрозия почвы увеличилась до высокого уровня, около 3700 тонн почвы на км 2 ежегодно. Лишенные растительности лёссовые почвы способствуют возникновению неприятных пыльных бурь и объясняют характерный цвет Желтой реки и Желтого моря.
По мере вырубки лесов эрозия почвы увеличилась до высокого уровня, около 3700 тонн почвы на км 2 ежегодно. Лишенные растительности лёссовые почвы способствуют возникновению неприятных пыльных бурь и объясняют характерный цвет Желтой реки и Желтого моря.
Пара уток-мандаринок. Изображение предоставлено: Фрэнсис Франклин, Creative Commons
Потенциальная растительность Лессового плато Центрального Китая представляет собой лиственный широколиственный лес из дуба и клена с березой, осиной и липой на возвышенностях. Леса когда-то были особенно богаты в северной и восточной частях экорегиона, обращенных к морю. Эта формация переходила на запад к низкой степной растительности, прилегающей к Внутренней Монголии. Адаптированные к холоду виды деревьев, такие как ель и рябина, растут в высокогорных районах, в то время как на более низких высотах, вблизи равнины Хуанхэ, растут вяз, ясень, Hovenia, и Pistacia как потенциальные виды климаксных деревьев.
Черный аист. Изображение предоставлено: Mino Zig, Creative Commons
Дикие млекопитающие нечасто встречаются на Лёссовом плато в центральном Китае. Теплые умеренные леса природного заповедника Ли Шань (248 км 2 ) на юге поддерживают редкие китайские популяции макак-резусов и находящихся под угрозой исчезновения лесных (или карликовых) кабарг. Этот олень, один из пяти видов кабарги, населяющих Китай, встречается южнее Вьетнама, но нигде не встречается, и его популяция, вероятно, сокращается. Люди из этого экорегиона будут одними из самых северных в метапопуляции, которая, по-видимому, содержит много локальных генетических вариаций, о которых пока мало что известно. Лесные кабарги умеют прыгать, что позволяет им прыгать в нижнюю крону дерева в поисках корма или уклоняться от хищников на земле.
Восстановление лесов на Лессовом плато Центрального Китая было начато в 1960-х годах для борьбы с потерей почвы. Совсем недавно на больших площадях была восстановлена растительность в рамках Инициативы «Зерно к зелени» с целью уменьшить эрозию почвы, обогатить естественную растительность и улучшить условия жизни в сельской местности. iv Эти усилия по восстановлению растительности направлены на стабилизацию склонов и стимулирование устойчивости сельского хозяйства, но не представляют собой лесовосстановление в строгом смысле.
iv Эти усилия по восстановлению растительности направлены на стабилизацию склонов и стимулирование устойчивости сельского хозяйства, но не представляют собой лесовосстановление в строгом смысле.
Беркут. Изображение предоставлено: Яркко Ярвинен, Creative Commons
Почвенные отложения Лессового плато Центрального Китая теряют объем к северо-востоку, так что там обнажается скалистая гористая местность. И, как и во многих других местах, именно эта менее продуктивная каменистая среда обитания наименее нарушена людьми. Например, природный заповедник Панцюань Гоу (105 км 2 ) в горах Лулян является важным районом размножения бурого ушастого фазана, самого редкого из фазанов семейства Crossoptilon , а также трех других охраняемых видов птиц: черного аиста, мандаринки. , и беркут. Панцюань Гоу также является типичным местом произрастания лиственницы Северного Китая, лиственного представителя семейства сосновых. Еще один заповедник – гора Луя (215 км 2 ), обеспечивает дополнительную среду обитания для бурого ушастого фазана.
Приоритетными природоохранными действиями на следующее десятилетие являются: 1) выявление и содействие сохранению оставшихся островов среды обитания в этом сильно измененном человеком экорегионе; 2) способствовать восстановлению лесов (в рамках инициативы «Зерно к зелени»), экологически близких к коренным насаждениям; и 3) соблюдать уже действующие законы, ограничивающие ловлю кабарги петлями и охоту на нее.
Цитаты
Карпентер, К. 2000. Южная Азия: Китай https://www.worldwildlife.org/ecoregions/pa0411. По состоянию на август 2018 г.
Sun X., Wang P. 2005. Сколько лет азиатской муссонной системе? — Палеоботанические записи из Китая. Палеогеография, палеоклиматология, палеоэкология : 222(3-4): 181-222.
Buckingham, K., Hanson, C. 2015. Пример диагностического случая восстановления: Лессовое плато Китая. Институт мировых ресурсов .
Пространственная неоднородность реакции дыхания почвы на импульс осадков в умеренном смешанном лесу в Центральном Китае Реакция дыхания почвы на импульс дождя
Пространственная неоднородность реакции дыхания почвы на импульс осадков в умеренном смешанном лесу в Центральном Китае реакция дыхания почвы на импульс осадков | Паб с открытым доступомПолный текст статьиАннотацияВведениеМатериалы и методыРезультатыОбсуждениеВыводыСсылки
Пространственная неоднородность реакции дыхания почвы на импульс осадков в смешанном лесу умеренного пояса в Центральном Китае
Yanchun Liu 1 Цин Шан 2 Чжунвэй Ван 2 Кешэн Чжан 3 Канкан Чжао 1
1 Международная совместная исследовательская лаборатория экологии глобальных изменений Школы естественных наук Хэнаньского университета, Кайфэн, Хэнань 475004, Китай
2 Технический институт охраны реки Хуанхэ, Кайфэн 475004, Хэнань, Китай
35 Лоянский институт науки и технологий, Лоян 471023, Хэнань, Китай
Abstract
Наличие воды является одним из основных факторов биологической активности и земного круговорота углерода. Хотя реакция почвенного дыхания на атмосферные осадки хорошо изучена в засушливых и полузасушливых экосистемах, наше понимание его характера в лесах весьма ограничено. Это исследование было проведено для изучения различий влияния осадков на дыхание почвы в различных условиях полога хвойных ( Pinus armandii Franch) и широколиственные ( Quercus Aliena var. острозубчатые ) смешанные леса в Центральном Китае. Результаты показали, что осадки значительно снизили температуру почвы, но увеличили объемную влажность почвы и дыхание почвы (6,0–35,3%). Осадки вызвали большее увеличение дыхания почвы под пологом лиственных деревьев (24,2%), чем под хвойными (13,5%). Вызванное осадками увеличение дыхания почвы было постоянно ниже под пологом мелкоразмерных классов (7,1%-32,6%), чем крупных классов (90,5%-33,3%). Среднее почвенное дыхание лесных промежутков увеличилось на 22,4% после осадков. Наше исследование подчеркивает положительную реакцию дыхания почвы на импульсы осадков в экосистемах с неограниченным количеством воды.
Хотя реакция почвенного дыхания на атмосферные осадки хорошо изучена в засушливых и полузасушливых экосистемах, наше понимание его характера в лесах весьма ограничено. Это исследование было проведено для изучения различий влияния осадков на дыхание почвы в различных условиях полога хвойных ( Pinus armandii Franch) и широколиственные ( Quercus Aliena var. острозубчатые ) смешанные леса в Центральном Китае. Результаты показали, что осадки значительно снизили температуру почвы, но увеличили объемную влажность почвы и дыхание почвы (6,0–35,3%). Осадки вызвали большее увеличение дыхания почвы под пологом лиственных деревьев (24,2%), чем под хвойными (13,5%). Вызванное осадками увеличение дыхания почвы было постоянно ниже под пологом мелкоразмерных классов (7,1%-32,6%), чем крупных классов (90,5%-33,3%). Среднее почвенное дыхание лесных промежутков увеличилось на 22,4% после осадков. Наше исследование подчеркивает положительную реакцию дыхания почвы на импульсы осадков в экосистемах с неограниченным количеством воды. Полученные данные свидетельствуют о том, что при оценке наземного круговорота углерода и его обратной связи с изменением климата следует подчеркивать пространственную неоднородность дыхания почвы по отношению к импульсу осадков в различных условиях растительного покрова.
Полученные данные свидетельствуют о том, что при оценке наземного круговорота углерода и его обратной связи с изменением климата следует подчеркивать пространственную неоднородность дыхания почвы по отношению к импульсу осадков в различных условиях растительного покрова.
Вклад авторов
Получено 12 ноября 2017 г .; Принят 08 декабря 2017 г .; Опубликовано 15 декабря 2017 г.;
Академический редактор: Sylvester Izah, Департамент биологических наук, Нигер Дельта Университет, Нигерия
Проверка на плагиат: Да
Обзор: Одно слепое
CopyRight © 2017 yanch и другие.
Лицензия
Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Конкурирующие интересы
Авторы заявили об отсутствии конкурирующих интересов.
Цитата:
Янчунь Лю, Цин Шан, Чжунвэй Ван, Кешэн Чжан, Канкан Чжао (2017) Пространственная неоднородность реакции дыхания почвы на импульс осадков в умеренном смешанном лесу в Центральном Китае. Журнал экологии растений и животных — 1(1):1-13.
Скачать как РИС, BibTeX, текст (включая аннотацию)
DOI 10.14302/issn.2637-6075.jpae-17-1863
Введение
Почвенное дыхание, первичный процесс, посредством которого CO 2 ассимилируется растениями в атмосферу1 основные компоненты цикла углерода (C) наземной экосистемы 2 . Предыдущие исследования показали, что на этот процесс влияют факторы окружающей среды 3, 4 . Температура и влажность почвы обычно определяются как два основных фактора, тесно связанных с дыханием почвы 5, 6 . Быстрый выброс CO 2 после выпадения осадков наблюдался во многих экосистемах 1 в результате быстрого микробного роста и минерализации доступного органического содержимого почвы, включая мертвую микробную биомассу 7 . В то время как наличие выбросов CO 2 является постоянным для изученных экосистем, величины отклика дыхания различаются между экосистемами из-за различных характеристик растительности, структуры почвы и условий питания. В лесо-луговой переходной зоне Лёссового плато выпадение осадков приводит к увеличению дыхания почвы в 1,6 раза для дубрав 5 . Однако выброс CO 2 увеличился до 30 раз после экспериментального повторного заболачивания в экосистеме пустыни Сонора 1 . Величина импульса CO 2 также чувствительна к количеству, времени и интенсивности осадков. Мансон и др. 8 обнаруживает, что общая потеря углерода и продолжительность повышенного потока CO 2 увеличивались с увеличением количества осадков в низкотравной степи.
Быстрый выброс CO 2 после выпадения осадков наблюдался во многих экосистемах 1 в результате быстрого микробного роста и минерализации доступного органического содержимого почвы, включая мертвую микробную биомассу 7 . В то время как наличие выбросов CO 2 является постоянным для изученных экосистем, величины отклика дыхания различаются между экосистемами из-за различных характеристик растительности, структуры почвы и условий питания. В лесо-луговой переходной зоне Лёссового плато выпадение осадков приводит к увеличению дыхания почвы в 1,6 раза для дубрав 5 . Однако выброс CO 2 увеличился до 30 раз после экспериментального повторного заболачивания в экосистеме пустыни Сонора 1 . Величина импульса CO 2 также чувствительна к количеству, времени и интенсивности осадков. Мансон и др. 8 обнаруживает, что общая потеря углерода и продолжительность повышенного потока CO 2 увеличивались с увеличением количества осадков в низкотравной степи. Считается, что гетеротрофное дыхание достигает асимптоты с увеличением размера осадков из-за кратковременной микробной активности 7, 9 , но для инициации автотрофного дыхания необходимы более крупные осадки из-за более глубокого распространения корневой системы 1 .
Считается, что гетеротрофное дыхание достигает асимптоты с увеличением размера осадков из-за кратковременной микробной активности 7, 9 , но для инициации автотрофного дыхания необходимы более крупные осадки из-за более глубокого распространения корневой системы 1 .
Многочисленные исследования, проведенные в засушливых и полузасушливых экосистемах (импульсные системы) 4, 7 , изучали влияние осадков на дыхание почвы и запасы почвенного углерода, а также соответствующую микробную активность 1, 5, 7, 10, 11 . Хотя наличие воды в почве не является ограничивающим фактором во влажных и полувлажных районах, дыхание почвы также может быть вызвано отдельными дождями. Кумулятивный смыв CO 2 в лесах, вероятно, вносит больший вклад в концентрацию CO 2 в атмосфере из-за большего поступления углерода и более частого выпадения осадков по сравнению с засушливыми и полузасушливыми ландшафтами. Например, Сюй и др. 12 обнаружил, что потери углерода в связи с летними дождями больше на участках с более высокой первичной продуктивностью и более высоким содержанием углерода в почве. Кроме того, пространственная неоднородность дыхания почвы, вероятно, больше, чем на пастбищах и пахотных землях, из-за более высокой неоднородности топографии и распределения растений. Например, сообщалось, что избыточное покрытие, виды и размерные классы могут вызывать различия в захвате растений, особенно при небольшом количестве осадков 10, 13, 14 . Следовательно, отклики почвенного дыхания на осадки в лесной экосистеме могут в значительной степени зависеть от породы деревьев и состояния полога, что является важным регулятором величины импульса из-за его биологического и физического воздействия на биотические и абиотические факторы почвы. Однако то, как частые колебания количества осадков влияют на дыхание почвы в лесных экосистемах, не оценивалось в такой степени, как в семиаридных и засушливых экосистемах 5 .
Например, Сюй и др. 12 обнаружил, что потери углерода в связи с летними дождями больше на участках с более высокой первичной продуктивностью и более высоким содержанием углерода в почве. Кроме того, пространственная неоднородность дыхания почвы, вероятно, больше, чем на пастбищах и пахотных землях, из-за более высокой неоднородности топографии и распределения растений. Например, сообщалось, что избыточное покрытие, виды и размерные классы могут вызывать различия в захвате растений, особенно при небольшом количестве осадков 10, 13, 14 . Следовательно, отклики почвенного дыхания на осадки в лесной экосистеме могут в значительной степени зависеть от породы деревьев и состояния полога, что является важным регулятором величины импульса из-за его биологического и физического воздействия на биотические и абиотические факторы почвы. Однако то, как частые колебания количества осадков влияют на дыхание почвы в лесных экосистемах, не оценивалось в такой степени, как в семиаридных и засушливых экосистемах 5 .
Экстремальные погодные явления часто случаются в результате изменения климата 15 . Засуха больше не бывает только в засушливых и полузасушливых экосистемах, она может иметь широкое распространение в наземных экосистемах 16, 17, 18 . Для лесов эпизодические поступления осадков могут влиять на фотосинтетическую и дыхательную активность растений 14 , а также на дыхание почвы, в том числе дыхание корнями растений и за счет гетеротрофии 5, 7 . Таким образом, изучение и количественная оценка реакции дыхания почвы на эпизодические осадки в лесных экосистемах имеют решающее значение для оценки наземного цикла углерода в условиях изменения климата.
В этом исследовании изучалась пространственная неоднородность реакции дыхания почвы на импульс осадков в умеренном смешанном лесу в Центральном Китае. Цели этого исследования состояли в том, чтобы (1) определить влияние видов деревьев с различной формой кроны на микроклимат почвы и дыхание под кроной после дождей; (2) сравнить реакцию дыхания почвы на импульс осадков среди разных классов размеров; и (3) анализировать суточную динамику дыхания почвы в условиях полога и лесных промежутков во время осадков.
Материалы и методы
Описание участка
Участок исследования, расположенный на лесоэкологической исследовательской станции Баотяньмэнь в Национальном природном заповеднике Баотяньмэнь (111°47′-112°04′ в.д., 33°20′-33°36′ с.ш.) , провинция Хэнань, Китай. Высота колеблется от 600-1800 м над уровнем моря. Среднегодовая температура составляет 15,1 ºС, месячная температура воздуха колеблется от 1,5 ºС в январе до 27,8 ºС в июле. Среднегодовое количество осадков составляет примерно 885,6 мм, из них 60% приходится на лето (с июня по август). В возвышенных почвах преобладала Haplic luvisol (классификация ФАО). Типичным лесом в этой области является лес с теплым умеренным климатом, с большим количеством лиственных широколиственных деревьев умеренного пояса и некоторыми компонентами растений из субтропических лесов. В лесах теплого умеренного пояса преобладают 9 видов деревьев.0129 Quercus spp., что составляет примерно 70% общей площади 19 . Доминирующими видами являются Toxicodendron verniciflnum (Stokes) F. A. Barkley, Carpinus cordata BI . , Acer spp., Platycarya strobilacea Sieb.ET. Zucc, Custanea seguinii Dode, Lindera obtusiloba BI..
A. Barkley, Carpinus cordata BI . , Acer spp., Platycarya strobilacea Sieb.ET. Zucc, Custanea seguinii Dode, Lindera obtusiloba BI..
Схема эксперимента
Эксперимент проводился в хвойно-широколиственном смешанном лесу на северо-восточной стороне с уклоном 28°. Для проведения эксперимента был выбран водораздел площадью ~ 1,5 га со средним уклоном 18°. Доминирующими видами были P. armandii и Q. алиена вар. острозубчатая , со средним диаметром на высоте груди 23,6 ± 7,90 см и 19,1 ± 8,62 см, что составляет 30,4% и 34,3% от общего покрова соответственно. Все растения в пределах водораздела были обследованы, и были зарегистрированы размеры, включая диаметр на высоте груди ( DBH ), высоту и состояние здоровья. Всего отобрано 16 деревьев двух классов диаметра (мелкий класс (~15 см) и большой класс (~30 см)) для P. armandii (PS и PL) и Q. Aliena var. острых серрат (QS и QL) были выбраны для исследования дыхания почвы под пологом (таблица 1). Диаметр кроны каждого выбранного дерева оценивали по двум вертикальным направлениям ( С-Ю , В-З ) с помощью метровой палки (Таблица 1). Для исследования дыхания почвы под открытым пологом были выбраны четыре лесные щели (ЛП) площадью около 5 м × 10 м.
Aliena var. острых серрат (QS и QL) были выбраны для исследования дыхания почвы под пологом (таблица 1). Диаметр кроны каждого выбранного дерева оценивали по двум вертикальным направлениям ( С-Ю , В-З ) с помощью метровой палки (Таблица 1). Для исследования дыхания почвы под открытым пологом были выбраны четыре лесные щели (ЛП) площадью около 5 м × 10 м.
Таблица 1. Характеристики кроны деревьев по размерным классам (среднее ± стандартная ошибка, n=4)
| Виды | Размерный класс | Среднее значение DBH (см) | Средний навес | Средняя корона |
| ширина (м) | глубина (м) | |||
| С. армандии | Маленький | 16,77±0,55 | 1,83×1,67 | 3,80±1,15 |
| Большой | 32,67±0,61 | 5,50×5,17 | 8,77±2,29 | |
В. алиена алиена | Маленький | 14,50±0,89 | 2,63×2,03 | 4,37±2,02 |
| Большой | 32,03±3,95 | 7,47×6,50 | 8,97±1,01 |
Измерение дыхания почвы
Дыхание почвы измеряли с помощью автоматизированной системы дыхания почвы (LI-8100, LI-COR, США) с июня по август 2014 г. Три поливинилхлоридных (ПВХ) воротника (190,6 см внутреннего диаметра и 8 см высоты) были вставлены в лесную подстилку на глубину 5 см вокруг каждого отобранного дерева под углом 120 ° на расстоянии 1 м от штамба. В каждую щель в лесу (FG) устанавливали по одному воротнику из ПВХ. Все ошейники оставались на участке на весь период исследования. Живые растения и очевидный мусор удаляли за 24 часа до измерения оттока.
Дыхание почвы без осадков (базовый уровень) выражали в виде средних значений, исследованных утром (8:30-11:00) и днем (2:30-5:00) 19 июня., а до этого не было эффективных осадков (> 1 мм) в течение почти 20 дней. Прилив дыхания почвы осадками измерялся примерно через 30 минут после каждого дождя. Всего за исследуемый период с 22 июня по 14 августа наблюдалось 9 осадков (22 июня – 5,81 мм, 24 июня – 5,01 мм, 28 июня – 3,43 мм, 6 июля – 14,03 мм, 16 июля – 4,13 мм, 19 июля). для 20,57 мм, 22 июля для 4,78 мм, 1-2 августа для 24,19 мм, 16 августа для 10,28 мм), в котором дыхание почвы двух дождей (6 июля, 16 июля) не учитывалось. Кроме того, для измерения суточной динамики дыхания почвы с 30-минутным интервалом под пологом полога и в просвете леса во время осадков были выбраны два дождевых события (22 июля, 1-2 августа).
Прилив дыхания почвы осадками измерялся примерно через 30 минут после каждого дождя. Всего за исследуемый период с 22 июня по 14 августа наблюдалось 9 осадков (22 июня – 5,81 мм, 24 июня – 5,01 мм, 28 июня – 3,43 мм, 6 июля – 14,03 мм, 16 июля – 4,13 мм, 19 июля). для 20,57 мм, 22 июля для 4,78 мм, 1-2 августа для 24,19 мм, 16 августа для 10,28 мм), в котором дыхание почвы двух дождей (6 июля, 16 июля) не учитывалось. Кроме того, для измерения суточной динамики дыхания почвы с 30-минутным интервалом под пологом полога и в просвете леса во время осадков были выбраны два дождевых события (22 июля, 1-2 августа).
Измерение температуры и влажности почвы
Температура почвы на расстоянии 10 см была измерена рядом с каждым дыхательным воротником с помощью портативного датчика температуры, подключенного к LI-8100. Объемное содержание воды в почве ( SVWC ) на глубине 0-5 см измеряли с помощью портативного рефлектометра MPKit-B (NTZT Inc., Наньтун, Китай) в трех точках, расположенных рядом с каждым воротником. Данные об осадках были собраны с ближайшей метеостанции, расположенной менее чем в 400 м от пробного участка.
Данные об осадках были собраны с ближайшей метеостанции, расположенной менее чем в 400 м от пробного участка.
Статистический анализ
Сначала были проверены нормальность и однородность дисперсии для полевых данных, а затем были использованы повторные измерения ANOVA для проверки значимости скорости дыхания почвы, объемного содержания почвы на глубине 0–5 см и температуры почвы на глубине 0–5 см. 10 см в зависимости от осадков, видов, размерного класса и их взаимодействия. Затем однофакторный дисперсионный анализ и апостериорный анализ ( LSD ) использовали для проверки значимости скорости дыхания почвы по сравнению с исходным уровнем, приращений скорости дыхания почвы после каждого дождя среди различных вариантов обработки (виды, размерный класс и лесной интервал). Данные выражены как среднее ±1 стандартная ошибка (SE). Статистический анализ был выполнен в SPSS 16.0 для Windows (SPSS Inc., Чикаго, Иллинойс, США).
Результаты
Температура и влажность почвы
Температура почвы без осадков (0 мм) была выше, чем после различных осадков для всех условий растительного покрова (рис. 1а). Все случаи осадков значительно снижали температуру почвы при PS, PL, QS, QL и FG (таблица 2, P <0,001). Температура почвы FG была значительно выше, чем у PS, PL, QS и QL при всех дождях. Температура почвы мелких классов (PS и QS) была ниже, чем у крупных (PL и QL) после каждого дождя (табл. 2, 9).0129 Р = 0,035). Кроме того, хвойные породы имели относительно более высокую температуру почвы, чем широколиственные (табл. 2, P = 0,0214).
1а). Все случаи осадков значительно снижали температуру почвы при PS, PL, QS, QL и FG (таблица 2, P <0,001). Температура почвы FG была значительно выше, чем у PS, PL, QS и QL при всех дождях. Температура почвы мелких классов (PS и QS) была ниже, чем у крупных (PL и QL) после каждого дождя (табл. 2, 9).0129 Р = 0,035). Кроме того, хвойные породы имели относительно более высокую температуру почвы, чем широколиственные (табл. 2, P = 0,0214).
Рис. 1. Температура почвы на высоте 10 см (a) и объемное содержание воды (SVWC) на высоте 5 см (b) для различных условий растительного покрова после различных осадков (среднее ± SE, n = 4). PS, PL, QS, QL и FG представляют собой P. armandii с классом мелкой и крупной размерности, Q. Aliena с классом мелкой и крупной размерности и лесной щелью соответственно.
Объемное содержание воды в почве (S VWC ) было чрезвычайно низким во всех четырех условиях растительного покрова без осадков (0 мм, все < 6%, рис. 1b). Осадки привели к заметному увеличению SVWC для всех условий растительного покрова (таблица 2, P < 0,0001). Как и в случае с температурой почвы, SVWC FG постоянно выше, чем под кронами деревьев (рис. 1b). SVWC для малых классов был значительно выше, чем для больших классов (таблица 2, 9).0129 Р = 0,0003). Кроме того, широколиственные породы имели более высокий SVWC , чем хвойные породы (табл. 2, P = 0,0461). Отмечено значительное влияние взаимодействий осадков и размерных классов на SVWC (Таблица 2, P =0,0051), что указывает на то, что различия SVWC между различными размерными классами растительного покрова уменьшаются с увеличением интенсивности осадков.
1b). Осадки привели к заметному увеличению SVWC для всех условий растительного покрова (таблица 2, P < 0,0001). Как и в случае с температурой почвы, SVWC FG постоянно выше, чем под кронами деревьев (рис. 1b). SVWC для малых классов был значительно выше, чем для больших классов (таблица 2, 9).0129 Р = 0,0003). Кроме того, широколиственные породы имели более высокий SVWC , чем хвойные породы (табл. 2, P = 0,0461). Отмечено значительное влияние взаимодействий осадков и размерных классов на SVWC (Таблица 2, P =0,0051), что указывает на то, что различия SVWC между различными размерными классами растительного покрова уменьшаются с увеличением интенсивности осадков.
Таблица 2. Результаты (значение P) повторных измерений ANOVA для скорости дыхания почвы (SR), объемного содержания влаги в почве (SVWC) на глубине 0-5 см и температуры почвы (ST) на глубине 10 см в ответ на осадки. , вид и размерный класс.
, вид и размерный класс.
| Факторы | СР | СВВК | СТ |
| Осадки | < 0,001 | <0,0001 | < 0,001 |
| Виды | 0,5891 | 0,0461 | 0,0214 |
| Размерный класс | 0,2191 | 0,0003 | 0,0349 |
| Виды × Размерный класс | 0,0071 | 0,4702 | 0,4724 |
| Осадки × виды | 0,5331 | 0,3841 | 0,5818 |
| Осадки × Класс размера | 0,0378 | 0,0051 | 0,1474 |
| Осадки × Виды × Размерный класс | 0,7141 | 0,5217 | 0,0581 |
Дыхание почвы
При 0 мм осадков дыхание почвы колебалось от 2,24 мкмоль м -2 с -1 для QS до 2,99 мкмоль·м -2 с -1 для QL со значительными различиями между QS и другими четырьмя типами ( P < 0,05). Кроме того, у QL было значительно более высокое дыхание почвы, чем у PL и FG ( P <0,05, рис. 2).
Кроме того, у QL было значительно более высокое дыхание почвы, чем у PL и FG ( P <0,05, рис. 2).
Рис. 2. Почвенное дыхание исходного уровня и соответствующие повышенные значения после осадков для различных классов размера и состояния растительного покрова (среднее значение ± стандартная ошибка, n=4). Различные строчные буквы указывают на значительные различия (P < 0,05) между различными состояниями растительного покрова (PS, PL, QS, QL и FG) для одного и того же случая осадков. PS, PL, QS, QL и FG представляют собой P. armandii с классом мелкой и крупной размерности, Q. Aliena с классом мелкой и крупной размерности и лесной щелью соответственно.
Для сравнения изменения величины дыхания почвы после различных осадков было рассчитано усиление дыхания почвы на основе дыхания почвы без осадков (рис. 2). Осадки вызвали значительное увеличение дыхания почвы в диапазоне от 6,0% до 35,3% (таблица 2, 90-129 P 90-130 < 0,001). Усиление дыхания почвы для мелкоразмерных классов (7,1–11,6 % для PS и 16,1–32,6 % для QS) в целом было ниже, чем для крупных (9). 0,5%-21,5% для PL и 12,5%-33,3% для QL), за исключением третьего наблюдаемого дождя (3,43 мм; таблица 2, P = 0,2191). Кроме того, малые классы размера (0,28 мкмоль м -2 с -1 для PS и 0,39 мкмоль м -2 с -1 для QS) показали относительно более высокое дыхание почвы по сравнению с большими классами (0,25 мкмоль м -2 с -1 для ФЛ и 0,37 мкмоль м -2 с -1 для КЖ, рисунок 2). Кроме того, хвойные породы (0,37 мкмоль м -2 с -1 с 13,5%) имели относительно более низкое среднее увеличение дыхания почвы, чем широколиственные виды (0,63 мкмоль м -2 с -1 с 24,2%). Почвенное дыхание для ФГ увеличилось на 0,62 мкмоль м -2 с -1 (0,17-0,98 мкмоль м -2 с -1 ) после различных осадков со значительно более высоким значением (0,74 мкмоль м -2 с -1 ), чем под кронами (0,26-0,39 мкмоль м -2 с -1 ) при событии 3,43 мм.
0,5%-21,5% для PL и 12,5%-33,3% для QL), за исключением третьего наблюдаемого дождя (3,43 мм; таблица 2, P = 0,2191). Кроме того, малые классы размера (0,28 мкмоль м -2 с -1 для PS и 0,39 мкмоль м -2 с -1 для QS) показали относительно более высокое дыхание почвы по сравнению с большими классами (0,25 мкмоль м -2 с -1 для ФЛ и 0,37 мкмоль м -2 с -1 для КЖ, рисунок 2). Кроме того, хвойные породы (0,37 мкмоль м -2 с -1 с 13,5%) имели относительно более низкое среднее увеличение дыхания почвы, чем широколиственные виды (0,63 мкмоль м -2 с -1 с 24,2%). Почвенное дыхание для ФГ увеличилось на 0,62 мкмоль м -2 с -1 (0,17-0,98 мкмоль м -2 с -1 ) после различных осадков со значительно более высоким значением (0,74 мкмоль м -2 с -1 ), чем под кронами (0,26-0,39 мкмоль м -2 с -1 ) при событии 3,43 мм. Отмечено значительное влияние взаимодействий осадков × размерный класс и вид × размерный класс на увеличение дыхания почвы (таблица 2, P = 0,0378; P = 0,0071). В период исследования увеличение дыхания почвы после средних дождей (5,81 мм и 5,01 мм) было выше, чем после малых и сильных дождей (3,43 мм, 20,57 мм и 24,19 мм).
Отмечено значительное влияние взаимодействий осадков × размерный класс и вид × размерный класс на увеличение дыхания почвы (таблица 2, P = 0,0378; P = 0,0071). В период исследования увеличение дыхания почвы после средних дождей (5,81 мм и 5,01 мм) было выше, чем после малых и сильных дождей (3,43 мм, 20,57 мм и 24,19 мм).
Суточная реакция дыхания почвы на осадки
Для щели леса наблюдалась относительно стабильная эмиссия почвенного дыхания до начала выпадения осадков. Почвенное дыхание активировалось и обычно усиливалось в течение 1,5 часов после выпадения осадков (рис. 3). Почвенное дыхание достигло своего пикового значения 4,83 мкмоль м -2 с -1 через 3 часа после дождя, что в 1,68 раза превышало среднее значение дыхания почвы перед дождем. По сравнению с дыханием почвы, исследуемым в ясный день, за тот же период наблюдалось увеличение до 20% за счет импульса осадков. Продолжительность стимулированного дыхания почвы составляла 3 часа, а затем постепенно снижалась до преддождевого уровня. В течение суток значительного снижения температуры почвы не наблюдалось; однако его пиковое значение было смещено примерно на 3,5 часа позже, чем в ясный день (рис. 3).
В течение суток значительного снижения температуры почвы не наблюдалось; однако его пиковое значение было смещено примерно на 3,5 часа позже, чем в ясный день (рис. 3).
Рис. 3. Динамические реакции дыхания почвы и температуры почвы на высоте 10 см для просвета в лесу на 4,78 мм осадков (закрашенные кружки, с 8:30 22 июля до 8:30 23 июля) и соответствующие временные зависимости на следующий ясный день (открытые кружки, с 8:30 23 июля до 8:30 24 июля). Серый прямоугольник указывает на временной интервал осадков с 13:50 до 15:45 22 июля.
Под пологом леса дыхание почвы резко увеличилось после импульса осадков, достигнув пикового значения 3,96 мкмоль м -2 с -1 через 1 час после дождя, что в 1,63 раза больше, чем среднее дыхание почвы перед дождем. Стимулированное дыхание почвы постепенно снижалось до уровня, предшествующего импульсу, через 1 час, а затем постоянно снижалось до более низкого уровня, отчасти из-за значительного снижения температуры почвы (рис. 4).
4).
Рис. 4. Динамические реакции дыхания почвы и температуры почвы на высоте 10 см для участка под пологом леса на 10,28 мм осадков (сплошные кружки, с 18:00 14 августа до 18:00 15 августа), и соответствующие курсы времени перед ливнем (незакрашенные кружки, с 18:00 13 августа до 18:00 14 августа). Серый прямоугольник указывает на временной интервал осадков с 13:45 до 18:25 14 августа.
Обсуждение
В нескольких исследованиях сообщается о реакции дыхания почвы на импульс осадков в различных экосистемах усиление дыхания почвы после дождя 1, 5, 8 . В отличие от нашего исследования, в других исследованиях сообщалось о значительном снижении дыхания почвы после дождя 22, 23, 24 . В этом исследовании наблюдалось повышенное дыхание почвы после осадков, но прирост дыхания почвы был ниже при осадках 24,19 мм, чем при умеренных дождях. Подобно предсказаниям Huxman et al. 7 , увеличение амплитуды пульса с 12 мм до 39 мм не вызывало более сильного или более продолжительного дыхания почвы во время осадков 13 . Большинство исследований связывают это с быстрой активацией активности почвенных микробов и физическим перемещением CO 9 .0197 2 из пор почвы после поверхностного увлажнения почвы 1, 13 . Поскольку поровое пространство в почве и подстилке может хранить CO 2 примерно в количестве, равном дневной продукции, опустошение или заполнение порового пространства может увеличить или уменьшить измеренный поток относительно биологической продукции 24, 25, 26 . Кроме того, Гу и соавт. 27 указал, что кратковременная потеря почвой CO 2 вызвана небольшим, быстро меняющимся пулом лабильного органического вещества.
7 , увеличение амплитуды пульса с 12 мм до 39 мм не вызывало более сильного или более продолжительного дыхания почвы во время осадков 13 . Большинство исследований связывают это с быстрой активацией активности почвенных микробов и физическим перемещением CO 9 .0197 2 из пор почвы после поверхностного увлажнения почвы 1, 13 . Поскольку поровое пространство в почве и подстилке может хранить CO 2 примерно в количестве, равном дневной продукции, опустошение или заполнение порового пространства может увеличить или уменьшить измеренный поток относительно биологической продукции 24, 25, 26 . Кроме того, Гу и соавт. 27 указал, что кратковременная потеря почвой CO 2 вызвана небольшим, быстро меняющимся пулом лабильного органического вещества.
Влияние температуры почвы на дыхание почвы
Общепризнано, что температура и влажность почвы играют важную роль в дыхании почвы. Линейная регрессия используется для сравнения различий в чувствительности дыхания почвы между видами и размерными классами к температуре (рис. 5а, в). Величина наклона линейной регрессии интерпретируется как количественная оценка чувствительности дыхания почвы к температуре 5 . Для хвойных пород больший уклон в мелком классе предполагает, что дыхание почвы более чувствительно к температуре под кронами небольших деревьев, чем крупных (рис. 5а). И наоборот, изменчивость чувствительности незначительна для широколиственных пород с неучтенно более высоким значением в классе крупных размеров (рис. 5с). Широколиственные породы проявляют относительно большую чувствительность к температуре по сравнению с хвойными породами. Аналогично результатам Сюй и Ван 28 и Shi et al. 5 , под кронами деревьев (рис. 5а, в) и в просвете леса (рис. 6) наблюдается отрицательная линейная зависимость между интенсивностью дыхания почвы и температурой почвы. Полученные данные означают, что отрицательное влияние температуры на дыхание почвы можно ожидать в засушливые сезоны 28, 29 , что отличается от результатов других исследований 20, 30, 31 .
Линейная регрессия используется для сравнения различий в чувствительности дыхания почвы между видами и размерными классами к температуре (рис. 5а, в). Величина наклона линейной регрессии интерпретируется как количественная оценка чувствительности дыхания почвы к температуре 5 . Для хвойных пород больший уклон в мелком классе предполагает, что дыхание почвы более чувствительно к температуре под кронами небольших деревьев, чем крупных (рис. 5а). И наоборот, изменчивость чувствительности незначительна для широколиственных пород с неучтенно более высоким значением в классе крупных размеров (рис. 5с). Широколиственные породы проявляют относительно большую чувствительность к температуре по сравнению с хвойными породами. Аналогично результатам Сюй и Ван 28 и Shi et al. 5 , под кронами деревьев (рис. 5а, в) и в просвете леса (рис. 6) наблюдается отрицательная линейная зависимость между интенсивностью дыхания почвы и температурой почвы. Полученные данные означают, что отрицательное влияние температуры на дыхание почвы можно ожидать в засушливые сезоны 28, 29 , что отличается от результатов других исследований 20, 30, 31 .
Рисунок 5. Почвенное дыхание в зависимости от температуры почвы на уровне 10 см и объемного содержания влаги в почве (SVWC) на уровне 5 см для P. armandii (a и b) и Q. Aliena (c и d). PS, PL, QS и QL представляют P. armandii с классом мелких и крупных размеров, Q. Aliena с классом мелких и крупных размеров.
Рис. 6. Дыхание почвы в зависимости от температуры почвы на высоте 10 см (a) и объемного содержания влаги в почве (SVWC) на высоте 5 см (b) для щели в лесу.
Влияние влажности почвы на дыхание почвы
Зависимости между дыханием почвы и влажностью почвы аппроксимированы полиномиальными функциями, которые предполагают значительное влияние влажности почвы на дыхание почвы, особенно для участков с крупными деревьями (табл. 9).0129 Р =0,00514; QL, P =0,0368; Рисунок 5б, г). Это может отчасти объяснить, почему места под пологом крупных размерных классов имеют более высокую величину дыхания почвы. Увеличение влажности почвы после дождей вызывает положительное усиление дыхания почвы на более ранней стадии, что в основном связано с увеличением активности микробов 25, 32 . Тем не менее, это имеет тенденцию к снижению с постоянным повышением влажности почвы и снижением температуры почвы 1 . Это согласуется с отчетом, указывающим на то, что содержание влаги в почве отрицательно коррелирует с дыханием почвы при содержании воды от умеренного до высокого 1, 22, 33 , вероятно, из-за эволюционной адаптации почвенных микроорганизмов к длительному средняя метеорологическая обстановка 22 . Более того, давно установлено, что скорость аэробных процессов в почвах возрастает с увеличением содержания воды вплоть до ограничения активности за счет диффузии кислорода через заполненные водой поровые пространства 34 . Во многих предыдущих исследованиях сообщалось о результатах, подобных этому исследованию, и было обнаружено, что примерно 60% водоудерживающей способности является порогом, при котором дыхание почвы начинает снижаться с увеличением содержания влаги в почве 5, 35 .
Увеличение влажности почвы после дождей вызывает положительное усиление дыхания почвы на более ранней стадии, что в основном связано с увеличением активности микробов 25, 32 . Тем не менее, это имеет тенденцию к снижению с постоянным повышением влажности почвы и снижением температуры почвы 1 . Это согласуется с отчетом, указывающим на то, что содержание влаги в почве отрицательно коррелирует с дыханием почвы при содержании воды от умеренного до высокого 1, 22, 33 , вероятно, из-за эволюционной адаптации почвенных микроорганизмов к длительному средняя метеорологическая обстановка 22 . Более того, давно установлено, что скорость аэробных процессов в почвах возрастает с увеличением содержания воды вплоть до ограничения активности за счет диффузии кислорода через заполненные водой поровые пространства 34 . Во многих предыдущих исследованиях сообщалось о результатах, подобных этому исследованию, и было обнаружено, что примерно 60% водоудерживающей способности является порогом, при котором дыхание почвы начинает снижаться с увеличением содержания влаги в почве 5, 35 .
Мелкоразмерный класс из двух видов имел относительно более низкую температуру почвы и более высокую SVWC , чем крупные классы после каждого события. Мы связываем это с меньшей шириной полога и высотой кроны (таблица 1), которые перехватывают меньше воды во время дождя. В целом, наши результаты подтверждают предыдущие исследования, в которых наличие влаги в почве более важно, чем температура почвы, для регулирования дыхания почвы в засушливые сезоны 26 .
Пространственная неоднородность дыхания почвы в различных условиях растительного покрова
Во время небольшого количества осадков (3,43 мм) дыхание почвы на ПЗ увеличивается в большей степени, чем на участках под растительным покровом, отчасти из-за эффекта быстрого пульса и быстрого усиления активность почвенных микробов в ФГ в результате отсутствия захвата растений во время дождей. Но меньший импульсный эффект возникает под пологом из-за большей доли перехвата полога после небольшого дождя. Аналогичные результаты недавно были получены Potts et al. 21 , который обнаружил, что на открытых участках в лесу дыхание почвы, как правило, выше, чем в подпологовых (под пологом), особенно после сезона дождей. Однако среди умеренных и больших осадков различия в дыхании почвы под пологом и в лесной щели незначительны, так как воды, достигающей поверхности почвы, достаточно для регуляции активности почвенных микробов 1 .
Аналогичные результаты недавно были получены Potts et al. 21 , который обнаружил, что на открытых участках в лесу дыхание почвы, как правило, выше, чем в подпологовых (под пологом), особенно после сезона дождей. Однако среди умеренных и больших осадков различия в дыхании почвы под пологом и в лесной щели незначительны, так как воды, достигающей поверхности почвы, достаточно для регуляции активности почвенных микробов 1 .
Суточные характеристики дыхания почвы показывают быстрое увеличение и постепенное снижение после достижения пикового значения через 1-1,5 часа после импульса осадков. Это связано с тем, что ограничение субстрата после повторного увлажнения ограничивает продолжительность микробного дыхания и, следовательно, общее содержание CO 2 производство 1 .
Влияние на круговорот углерода
Хотя импульсы дыхания почвы после дождя невелики по сравнению с годовой потерей, они играют важную роль в балансе углерода в экосистеме из-за высокой частоты появления в лесных экосистемах. Подсчитано, что почти 60% случаев осадков выпадает ниже 5 мм на основе данных об осадках с 2009 по 2010 год в этом регионе (таблица 3), что предлагается в качестве нижнего порога для реакции растений на дождь 36 . Следовательно, в соответствии с этим порогом около 60% событий слишком малы, чтобы вызвать реакцию растений, но достаточны, чтобы вызвать потери CO 2 на основе микробов. В отличие от водоограниченных экосистем количество осадков выше, чем в безводном регионе с более высоким преддождевым дыханием почвы. Это означает, что выпадает больше импульса осадков и индуцируется большая эмиссия CO 2 из почвы, даже если относительные изменения увеличения CO 9019Эмиссия 7 2 (6,0-35,3%) ниже, чем в водоограниченных экосистемах (в 4-8 раз 37 ; в 4-5 раз 21 ; в 1,6-2,3 раза 5 ). В результате безводная экосистема в условиях дождей может действовать как чистый источник CO 2 в атмосферу 1 .
Подсчитано, что почти 60% случаев осадков выпадает ниже 5 мм на основе данных об осадках с 2009 по 2010 год в этом регионе (таблица 3), что предлагается в качестве нижнего порога для реакции растений на дождь 36 . Следовательно, в соответствии с этим порогом около 60% событий слишком малы, чтобы вызвать реакцию растений, но достаточны, чтобы вызвать потери CO 2 на основе микробов. В отличие от водоограниченных экосистем количество осадков выше, чем в безводном регионе с более высоким преддождевым дыханием почвы. Это означает, что выпадает больше импульса осадков и индуцируется большая эмиссия CO 2 из почвы, даже если относительные изменения увеличения CO 9019Эмиссия 7 2 (6,0-35,3%) ниже, чем в водоограниченных экосистемах (в 4-8 раз 37 ; в 4-5 раз 21 ; в 1,6-2,3 раза 5 ). В результате безводная экосистема в условиях дождей может действовать как чистый источник CO 2 в атмосферу 1 . Помимо измерений суточных режимов двух ошейников, мы не проводили непрерывных измерений для всего дыхания почвы, что может привести к завышению нашего результата.
Помимо измерений суточных режимов двух ошейников, мы не проводили непрерывных измерений для всего дыхания почвы, что может привести к завышению нашего результата.
Таблица 3. Количество осадков в 2009 и 2010 гг. в Баотяньмане. Данные за 2011 год не были включены, так как данные были пропущены из-за повреждения от грозы и проблем с аккумуляторной батареей.
| Год | Размерный класс (мм) | № случая осадков | Осадки (мм) |
| 2009 | <1 | 37 | 8,45 |
| 01 мая | 21 | 46,1 | |
| 05 октября | 11 | 76,91 | |
| 20 октября | 11 | 143,83 | |
| >20 | 19 | 655,96 | |
| Всего | 99 | 931,25 | |
| 2010 | <1 | 25 | 9,81 |
| 01 мая | 19 | 85,8 | |
| 05 октября | 5 | 41,68 | |
| 20 октября | 8 | 97,28 | |
| >20 | 15 | 752,32 | |
| Всего | 72 | 986,89 |
Выводы
В умеренном смешанном лесу в Центральном Китае мы обнаружили, что эпизодические осадки оказывают значительное влияние на дыхание почвы, хотя доступность воды не является ограничивающим фактором в этой экосистеме. Почвенное дыхание под широколиственными породами относительно сильнее реагирует на импульс осадков по сравнению с хвойными. Кроме того, прирост дыхания почвы под деревьями с малой кроной был ниже, чем с большой кроной. Наше исследование предполагает, что различия в морфологии растительного покрова между разными видами играют важную роль в регулировании величины дыхания почвы после осадков. Полученные данные показывают, что при оценке земного круговорота углерода и его обратной связи с изменением климата следует учитывать пространственную неоднородность реакции дыхания почвы на пульсацию осадков в различных условиях растительного покрова.
Почвенное дыхание под широколиственными породами относительно сильнее реагирует на импульс осадков по сравнению с хвойными. Кроме того, прирост дыхания почвы под деревьями с малой кроной был ниже, чем с большой кроной. Наше исследование предполагает, что различия в морфологии растительного покрова между разными видами играют важную роль в регулировании величины дыхания почвы после осадков. Полученные данные показывают, что при оценке земного круговорота углерода и его обратной связи с изменением климата следует учитывать пространственную неоднородность реакции дыхания почвы на пульсацию осадков в различных условиях растительного покрова.
Благодарности
Это исследование финансировалось Китайским национальным научным фондом (№ 31600379, 31640011), ключевым исследовательским проектом колледжей и университетов провинции Хэнань (№ 17A180001) и проектом Фонда научных исследований Хэнаньского университета (2015YBZR035). . Мы благодарим Ye Tian, Yin Wu и Xiaojing Liu за помощь в полевых условиях во время этого эксперимента, а также Jingxin Wang из Университета Западной Вирджинии за полезные комментарии к более ранним версиям этой рукописи.
Каталожные номера
1. R A Спонсор. (2007) Импульсы осадков и поток CO2 в почве в экосистеме пустыни Сонора. , Глоб. Изменить Биол 13, 426-436.
Поиск в Google Scholar
2. Zhang X, Tan Y, Zhang B, Li A, Daryanto S. (2017) Влияние увеличения количества осадков и добавления азота на дыхание почвы в полузасушливой умеренной степи. , Ecosphere 8, 01655.
Поиск в Google Scholar
3. M, Egea G, Martin-Gorriz B, P A Nortes, Baile A. (2017) Пространственная изменчивость оттока CO2 из почвы в старых и молодые цитрусовые сады и их зависимость от биотических и абиотических факторов. , Геодерма 294, 29-37.
ScienceDirect·Scopus·Поиск в Google Scholar
4. Rey A, Oyonarte C, Morán-López T, Raimundo J, Pegoraro E. (2017) Изменения влажности почвы прогнозируют потери углерода в почве при повторном увлажнении в многолетней полузасушливой зоне Степь на юго-востоке Испании. , Геодерма 287, 135-146.
(2017) Изменения влажности почвы прогнозируют потери углерода в почве при повторном увлажнении в многолетней полузасушливой зоне Степь на юго-востоке Испании. , Геодерма 287, 135-146.
ScienceDirect · Scopus · Поиск в Google Scholar
5. W Y Shi, Tateno R, J G Zhang, Y L Wang, Yamanaka N. (2011) Реакция дыхания почвы на осадки в засушливый сезон в двух типичных лесонасаждениях в лесно-луговая переходная зона Лёссового плато. , Агр. За. Метеорол 151, 854-863.
Поиск в Google Scholar
6. Liu Y, Liu S, Wan S, Wang J, Luan J. (2016) Дифференциальные реакции дыхания почвы на потепление почвы и экспериментальное снижение сквозного стока в переходном дубовом лесу в центральном Китае . , Агр. За. Метеорол 226, 186-198.
ScienceDirect · Scopus · Поиск в Google Scholar
7. Т. Э. Хаксман, К. А. Снайдер, Тиссью Д., А. Дж. Леффлер, Огл К. (2004) Импульсы осадков и потоки углерода в полузасушливых и засушливых экосистемах. , Экология 141, 254-268.
Э. Хаксман, К. А. Снайдер, Тиссью Д., А. Дж. Леффлер, Огл К. (2004) Импульсы осадков и потоки углерода в полузасушливых и засушливых экосистемах. , Экология 141, 254-268.
Поиск в Google Scholar
8. С. М. Мансон, Т. Дж. Бентон, В. К. Лауэнрот, И. К. Берк. (2009) Поток почвенного углерода после импульсных осадков в низкотравной степи. , экол. Рез 25, 205-211.
Поиск в Google Scholar
9. Дж. Р. Велтер, С. Г. Фишер, Н. Б. Гримм. (2005) Перенос и удержание азота в водоразделе засушливой земли: влияние характеристик шторма на наземно-водные связи. , Биогеохимия 76, 421-440.
Поиск в Google Scholar
10. Дж. М. Кейбл, Т. Э. Хаксман. (2004) Влияние размера импульса осадков на микробные корки почвы пустыни Сонора. , Экология 141, 317-324.
Поиск в Google Scholar
11. Мюр Дж., Франке Дж., Боркен В. (2010) Процессы высыхания-повторного увлажнения снижают потери углерода и азота из подстилки из ели европейской. , Почвы. биол. Биохим 42, 1303-1312.
Мюр Дж., Франке Дж., Боркен В. (2010) Процессы высыхания-повторного увлажнения снижают потери углерода и азота из подстилки из ели европейской. , Почвы. биол. Биохим 42, 1303-1312.
Поиск в Google Scholar
12. Xu L, D, Tang J. (2004) Как влажность почвы, импульсы дождя и рост изменяют реакцию дыхания экосистемы на температуру. , Глоб. Биогеохим. Цикл 18, 4002.
Поиск в Google Scholar
13. Д.Л. Поттс, Т.Э. Хаксман, Дж.М. Кейбл, Н.Б. обмен на импульсы дождя в полузасушливых пастбищах. , Новый Фитол 170, 849-860.
Поиск в Google Scholar
14. В. Э. Эммерих, К. Л. Вердуго. (2008) Пороговые значения осадков для поглощения CO2 сообществами травянистых и кустарниковых растений на экспериментальном водоразделе Орехового ущелья. , Исследование водных ресурсов 44, н/д-н/д. .
.
Поиск в Google Scholar
15. IPCC. (2014) Изменение климата 2014: Обобщающий отчет. Вклад рабочих групп I, II и III в Пятый оценочный доклад Межправительственной группы экспертов по изменению климата. , Team, C
Поиск в Google Scholar
16. Martinho F, Leitão R, Viegas I, Dolbeth M, JM Neto. (2007) Влияние экстремальной засухи на рыбное сообщество эстуария южной Европы с умеренным климатом. , Устье. Побережье. Полка Sci 75, 537-546.
Поиск в Google Scholar
17. Shen C, Wang W-C, Hao Z, Gong W. (2007) Исключительные засухи в восточном Китае за последние пять столетий. , Изменение климата 85, 453-471.
Поиск в Google Scholar
18. Дж. Л. Чен, К. Р. Уилсон, Б. Д. Тапли, З. Л. Ян, Г. Ю. Ниу. (2009) засуха в бассейне реки Амазонки, измеренная GRACE и оцененная с помощью климатических моделей. , Дж. Геофиз. Res 114, 05404.
, Дж. Геофиз. Res 114, 05404.
Поиск в Google Scholar
19. C S Song. (1994) Научный обзор национального природного заповедника Баотяньмань, Китайское лесное издательство. , Пекин
Поиск в Google Scholar
20. Chen S, Lin G, Huang J, He M. (2008) Реакция дыхания почвы на смоделированные импульсы осадков в полузасушливой степи при различных режимах выпаса скота. , J. Plant Ecol 1, 237-246.
Поиск в Google Scholar
21. Д. Л. Поттс, Р. Л. Скотт, Дж. М. Кейбл, Т. Э. Хаксман, Д. Г. Уильямс. (2008) Чувствительность обмена CO2 в мескитовых кустарниках к осадкам в контрастных ландшафтных условиях. , Экология 89, 2900-2910.
Поиск в Google Scholar
22. DÖRR H, K O Münnich. (1987) Годовой ход дыхания почвы в отдельных районах умеренного пояса. , Теллус В 39, 114-121.
, Теллус В 39, 114-121.
Поиск в Google Scholar
23. B C, Scott A, JP Parker. (1999) Полевые потоки N2O, CO2 и Ch5 в связи с обработкой почвы, уплотнением и качеством почвы в Шотландии. , Обработка почвы. Рез 53, 29-39.
Поиск в Google Scholar
24. М. Л. Томи, С. Л. Коллинз, Варгас Р., Дж. Э., Р. Ф. Браун. (2011) Влияние изменчивости осадков на чистую первичную продукцию и дыхание почвы на пастбищах пустыни Чиуауа. , Глоб. Изменение Биол 17, 1505-1515.
Поиск в Google Scholar
25. М. Г. Райан, Б. Е. Лоу. (2005) Интерпретация, измерение и моделирование дыхания почвы. , Биогеохимия 73, 3-27.
Поиск в Google Scholar
26. Liu Y, Liu S, Wang J, Zhu X, Zhang Y. (2014) Изменение дыхания почвы под кронами деревьев в смешанном лесу умеренного пояса, центральный Китай, при различных условиях почвенной воды. , экол. Рез 29, 133-142.
, экол. Рез 29, 133-142.
Поиск в Google Scholar
27. Gu L, WM Post, AW King. (2004) Быстрый лабильный оборот углерода скрывает чувствительность гетеротрофного дыхания из почвы к температуре: модельный анализ. , Глоб. Биогеохим. Цикл 18, 1022.
Поиск в Google Scholar
28. Xu W, Wan S. (2008) Опосредованные водой и растениями реакции дыхания почвы на топографию, огонь и азотные удобрения на полузасушливых пастбищах в северном Китае. , Почвы. биол. Биохим 40, 679-687.
Поиск в Google Scholar
29. Keith H, Jacobsen K, Raison R. (1997) Влияние наличия фосфора в почве, температуры и влажности на дыхание почвы в лесу Eucalyptus pauciflora. , Почва растений 190, 127-141.
Поиск в Google Scholar
30. Хашимото Т., Миура С., Ишизука С. (2009) Температура контролирует временные колебания в выделении CO2 почвой во вторичном буковом лесу в горах Аппи. , Япония, Дж. Фор. Рез 14, 44-50.
Хашимото Т., Миура С., Ишизука С. (2009) Температура контролирует временные колебания в выделении CO2 почвой во вторичном буковом лесу в горах Аппи. , Япония, Дж. Фор. Рез 14, 44-50.
Scopus·Поиск в Google Scholar
31. G M Malcolm, J C López-Gutiérrez, R T Koide. (2009) Температурная чувствительность дыхания различается между ярусами лесной подстилки на плантациях Pinus Resinosa. , Почвы. биол. Биохим 41, 1075-1079.
ScienceDirect · Scopus · Поиск в Google Scholar
32. В. А. Орчард, Кук Ф. (1983) Связь между дыханием почвы и влажностью почвы. , Почвы. биол. Биохим 15, 447-453.
Поиск в Google Scholar
33. Дэвидсон Э., Белк Э., Р. Д. Бун. (1998) Содержание влаги в почве и температура как независимые или смешанные факторы, контролирующие дыхание почвы в смешанном лиственном лесу умеренного пояса. , Глоб. Изменить Биол 4, 217-227.
, Глоб. Изменить Биол 4, 217-227.
Поиск в Google Scholar
34. Р. Д. Миллер, Джонсон Д. (1964) Влияние натяжения почвенной влаги на выделение двуокиси углерода, нитрификацию и минерализацию азота. 644-647.
Поиск в Google Scholar
35. Линн Д., Доран Дж. (1984) Влияние заполненного водой порового пространства на образование двуокиси углерода и закиси азота в обрабатываемых и необрабатываемых почвах. , Почвоведение Soc Amer J 48, 1267-1272.
Поиск в Google Scholar
36. Дж. Ф. Рейнольдс, П. Р. Кемп, Огл К., Р. Дж. Фернандес. (2004) Модификация «импульсного резерва»; парадигма для пустынь Северной Америки: импульсы осадков, почвенная вода и реакция растений. , Экология 141, 194-210.
Поиск в Google Scholar
37. WD Borken, Savage E, Gaudinski K, Trumbore J, S. (2003) Влияние высыхания и увлажнения на выброс углекислого газа из органических горизонтов. , Soil Sci Soc Amer J 67, 1888.
WD Borken, Savage E, Gaudinski K, Trumbore J, S. (2003) Влияние высыхания и увлажнения на выброс углекислого газа из органических горизонтов. , Soil Sci Soc Amer J 67, 1888.
Искать в Google Scholar
Цитируется по (1)
1. Курганова И. Н., Лопес де Гереню В. О., Мякшина Т. Н., Сапронов Д. В., Ромашеткин Д. В., Ромашеткин Д. В. 2020, Экспериментальные и модельные оценки дыхания лесной дерново-подзолистой почвы Приокско-Террасного заповедника, Современные проблемы экологии, 13(7), 813, 10.1134/S1995425520070057
Практический пример в районе Олту, Эрзурум
Детали
| Основной язык | Английский |
|---|---|
| Субъекты | Лесное хозяйство, Почвоведение |
| Дата публикации | март 2022 |
| Раздел журнала | Orman Mühendisliği / Лесная техника |
| Авторы | Эмре ЧОМАКЛИ> (основной автор) Аднан БИЛГИЛИ> Ташкин ОЗТАШ> Тугба ЧОМАКЛИ
Это я |
| Спасибо | Arazi çalışmalarında ulaşım desteğini veren Doğu Anadolu Ormancılık Araştırma Enstitüsü Müdürlüğüne teşekkür ederiz. |
| Дата публикации | 1 марта 2022 г. |
| Дата подачи заявки | 1 августа 2021 г. |
| Дата приемки | 23 декабря 2021 г. |
| Опубликовано в выпуске | 2022 год, Том 12, Выпуск 1 |
 977224},
title = {Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум},
ключ = {цитировать},
автор = {Чомаклы, Эмре и Билгили, Аднан и Озташ, Ташкин и Чомаклы, Тугба}
}
977224},
title = {Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум},
ключ = {цитировать},
автор = {Чомаклы, Эмре и Билгили, Аднан и Озташ, Ташкин и Чомаклы, Тугба}
} Определение запасов углерода, азота и фосфора и стехиометрии в почве широколиственных смешанных лесов и подстилке: тематическое исследование в районе Олту, Эрзурум
.
Журнал Института науки и технологий
, 12 (1) ,
464-475 .
DOI: 10.21597/млн.977224
Определение запасов углерода, азота и фосфора и стехиометрии в почве широколиственных смешанных лесов и подстилке: тематическое исследование в районе Олту, Эрзурум
.
Журнал Института науки и технологий
, 12 (1) ,
464-475 .
DOI: 10.21597/млн.977224 , Озташ, Т.
, Чомаклы, Т.
«Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум»
.
Журнал Института науки и технологий 12 (2022 г.)
): 464-475
, Озташ, Т.
, Чомаклы, Т.
«Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум»
.
Журнал Института науки и технологий 12 (2022 г.)
): 464-475  , Билгили, А.
, Озташ, Т.
, Чомаклы, Т.
«Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум».
Журнал Института науки и технологий 12 (2022 г.)
): 464-475
, Билгили, А.
, Озташ, Т.
, Чомаклы, Т.
«Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум».
Журнал Института науки и технологий 12 (2022 г.)
): 464-475 21597/jist.977224 DO — 10.21597/jist.977224 T2 — Journal of the Institute of Science and Technology
JF — Журнал
ДЖО — ДЖОР
СП — 464
ЭП-475
ВЛ — 12
ИС — 1
СН — -2536-4618
M3 — дои: 10.21597/jist.977224 UR — https://doi.org/10.21597/jist.977224 Y2 — 2021
Скорая помощь —
21597/jist.977224 DO — 10.21597/jist.977224 T2 — Journal of the Institute of Science and Technology
JF — Журнал
ДЖО — ДЖОР
СП — 464
ЭП-475
ВЛ — 12
ИС — 1
СН — -2536-4618
M3 — дои: 10.21597/jist.977224 UR — https://doi.org/10.21597/jist.977224 Y2 — 2021
Скорая помощь — «Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум».
Журнал Института науки и технологий
12
/
1
(март 2022 г.):
464-475
.
«Определение запасов углерода, азота и фосфора и стехиометрии в почве и опаде широколиственных смешанных лесов: тематическое исследование в районе Олту, Эрзурум».
Журнал Института науки и технологий
12
/
1
(март 2022 г.):
464-475
. https://doi.org/10.21597/млн.977224
https://doi.org/10.21597/млн.977224

