растительный и животный мир, географическое положение
Бескрайние просторы, залитые ярким солнцем и поросшие густой травой — все это представляет собой степь, практически лишенная деревьев, за исключением берегов рек и водоемов. Чуть севернее, ближе к лесной зоне, начинаются лесостепи. Здесь уже к травянистой равнине начинают добавляться островки лесов и небольших рощ.
Лесостепи и степи играют огромную роль в деятельности человека, потому что почвы в этих местах отличаются наилучшей плодородностью.
Что такое природные зоны
В первую очередь следует дать определение самому понятию природных зон: это территория, для которой характерны определенные климатические условия, рельеф, почва, животный и растительный мир.
На планете существуют следующие территориальные экосистемы:
- арктическая пустыня;
- тундра и лесотундра;
- тайга;
- смешанные леса и широколиственные леса;
- степи и лесостепи;
- пустыни и полупустыни;
- жестколистные леса и кустарники;
- саванны и редколесья;
- экваториальные тропические леса.

Все биомы сформировались на основе климатических поясов: Арктика находится в арктическом поясе, тундра и лесотундра — в субарктическом, лесная зона и степи — в умеренном, жестколистные леса — в субтропическом, пустыни и полупустыни — в тропическом, саванны и редколесья — в субэкваториальном, тропические леса — в экваториальном.
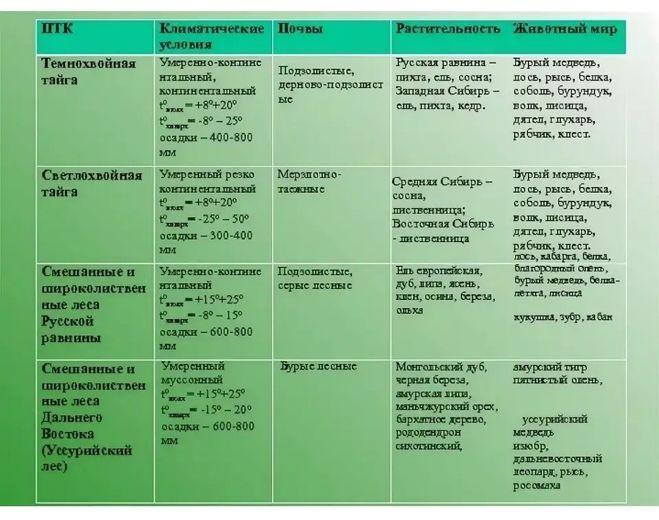
В таблице приведен перечень основных природных зон и их отличительные особенности.
Степи и лесостепи
Слово «степь» не имеет точного происхождения. Предполагается, что оно означает «вырубленное место», то есть лишенное деревьев. По мнению других, название связано со словом «топтать», иными словами, «вытоптанное место».
У разных стран данное понятие имеет свое название: в Южной Америке — пампасы, в Северной Америке — прерии, в Новой Зеландии — туссоки.
Географическое положение
Степная зона встречается практически на всех континентах. Североамериканские прерии покрывают центральную часть США, север Мексики и запад Канады. Пампасы Южной Америки расположены в юго-восточной части, между Андами и Атлантическим океаном. В Евразии травянистые равнины занимают обширные площади Монголии, Казахстана, Узбекистана, южной части России, Украины.
Пампасы Южной Америки расположены в юго-восточной части, между Андами и Атлантическим океаном. В Евразии травянистые равнины занимают обширные площади Монголии, Казахстана, Узбекистана, южной части России, Украины.
В России зона степей расположилась на юге Восточно-Европейской и Западно-Сибирской равнин. Отдельные участки встречаются в Забайкалье. С севера на юг степной регион протянулся от Тулы и рек Кама и Белая до гор Кавказа.
Лесостепи узкой полосой тянутся вдоль степных полей, отделяя их от лесных массивов.
Климат
Степи расположены в умеренном и субтропическом климате. Для этих мест характерна континентальность, то есть удаленность от морских и океанических побережий. Из этого следует, что лето здесь жаркое и засушливое, а зимнее время — холодное и малоснежное.
Климатические условия различаются в зависимости от местоположения степи. Заволжские районы являются переходной территорией между западными влажными степями и сухими континентальными областями на востоке. В районе Крыма климат становится более мягким и влажным: в зимнее время температура редко опускается ниже 0°С, а летом в среднем составляет +22°С. Однако для этих районов характерны суховеи.
В районе Крыма климат становится более мягким и влажным: в зимнее время температура редко опускается ниже 0°С, а летом в среднем составляет +22°С. Однако для этих районов характерны суховеи.
В лесостепях климат более мягкий: зимой показатели температуры составляют от -2 до -20°С.
Животный мир
Травянистые равнины населены разнообразными животными и птицами. В основном здесь живут травоядные, которым степь дает все необходимое для жизни. Но, как и в любой экосистеме, тут обитают и хищники.
Что касается лесостепей, то для них характерны как обитатели степей, так и лесные жители.
Степной волк
От обычных представителей своего вида отличается более мелкими размерами и светлой, желтовато-серой шерстью. Охотятся эти хищники на степных копытных (сайгаки и лошади), но могут питаться и более мелкой добычей: кроликами, зайцами, птицами, грызунами. Степные волки предпочитают селиться на берегах рек или в балках.
Корсак
Корсак, или степная лисица, обитает в норах, которые остались после барсуков, сусликов и других животных. Корсак охотится на грызунов и птиц, а зимой уходит на юг в поисках пропитания. Основными врагами лисицы являются волки, хищные птицы и человек, охотящийся на корсака ради пушистого меха.
Корсак охотится на грызунов и птиц, а зимой уходит на юг в поисках пропитания. Основными врагами лисицы являются волки, хищные птицы и человек, охотящийся на корсака ради пушистого меха.
Манул
Представитель семейства кошачьих, обладающий очень пушистым и густым мехом. Его шерсть окрашена в светло-серые, с рыжеватым оттенком, цвета. Этот дикий кот обитает в степях и лесостепях Центральной и Средней Азии. Может населять и горы в этих регионах. Ведет ночной образ жизни, питаясь различными грызунами, птицами и зайцами.
Сайгак
Эта уникальная разновидность антилопы обитает в степях Евразии. Отличительная особенность сайгака — большой нос, похожий на короткий хобот. Степные антилопы ведут кочевой образ жизни, постоянно перемещаясь в поисках новых пастбищ. Стадо сайгаков может насчитывать до нескольких сотен голов.
Тушканчик
Небольшой грызун, имеющий сильные задние лапы, на которых и перемещается. Также его характерной чертой является длинный хвост с большой кисточкой на конце. Тушканчики ведут ночной образ жизни, днем прячась в норах. Питаются побегами растений и семенами.
Также его характерной чертой является длинный хвост с большой кисточкой на конце. Тушканчики ведут ночной образ жизни, днем прячась в норах. Питаются побегами растений и семенами.
Степной орел
У этой птицы очень острое зрение, позволяющее ей высматривать добычу с большой высоты. Пропитанием орлу служат суслики, мелкие птицы, ящерицы, змеи. Окрас оперения темно-бурый, на крыльях имеются поперечные серые полосы.
Дрофа
Крупная птица, обитающая в ковыльных и полынных степях. Дрофа имеет пестрый окрас, состоящий из белых, серых, черных и рыжих перьев. Питается насекомыми, грызунами, ящерицами. Может употреблять в пищу побеги и семена. В настоящее время данный вид находится под угрозой.
Желтобрюхий полоз
Крупных размеров змея, чешуя которой окрашена в черный цвет, а низ, соответственно названию, имеет желтоватый оттенок. Данный вид змей не ядовит, хотя ее укусы могут причинить сильную боль. Питается грызунами, ящерицами и птицами. На зиму впадает в спячку.
Питается грызунами, ящерицами и птицами. На зиму впадает в спячку.
Растения
В степях произрастают в основном травы и кустарники. Деревья, ввиду засушливости территории, встречаются только по берегам рек и в лесостепной зоне. Характеристика главных представителей фауны представлена ниже.
Ковыль
Злаковое растение, имеющее на вершинах стеблей длинные светлые колоски. Поле, заросшее ковылем, кажется похожим на зелено-серебристый океан. Имеет слаборазвитую корневую систему, вследствие чего не произрастает на луговых полях с хорошей дерниной. Зато ковыль весьма устойчив к засухе, поэтому может встретиться на выжженных участках или сенокосах.
Шалфей степной
Шалфей покрывает луга и поля в степной и лесостепной зоне. Имеет соцветия из сиреневых и фиолетовых цветков. Это растение широко применяется в медицине и косметологии благодаря своим противовоспалительным и очищающим свойствам.
Мятлик луговой
Является растением из семейства злаковых, имеет длинные метелки с множеством колосков. Обладает хорошей устойчивостью к морозам и засухе. Мятлик высоко ценится в качестве пастбищного растения благодаря питательным веществам и витаминам.
Обладает хорошей устойчивостью к морозам и засухе. Мятлик высоко ценится в качестве пастбищного растения благодаря питательным веществам и витаминам.
Касатик безлистный
Произрастает в зарослях кустарников или по берегам рек. Виды этого растения имеют еще и другие названия — ирис безлистный или касатик вильчатый. Они произошли из-за того, что листья ириса опадают зимой, а весной появляются позднее цветоносов. Высота может достигать полуметра. Касатик — растение с красивыми крупными цветками фиолетового цвета с желтой серединой. Считается редким видом и занесен в Красную книгу.
Василек синий
Представляет собой сорняковое растение и часто соседствует с посевами ржи. Тонкие стебли украшены синими цветками, растущими поодиночке. Семена василька обладают лекарственными свойствами, их порошок используют как противовоспалительное средство.
Вишня степная
От других представителей своего вида отличается небольшой высотой. Цветки, как и у обычной вишни, белого цвета. Плоды представляют собой костянку темно-красного оттенка и имеют кисло-сладкий вкус. Любит ярко-освещенные места, устойчива к морозам и засушливой погоде.
Цветки, как и у обычной вишни, белого цвета. Плоды представляют собой костянку темно-красного оттенка и имеют кисло-сладкий вкус. Любит ярко-освещенные места, устойчива к морозам и засушливой погоде.
В лесостепях участки полей и лугов чередуются с небольшими лесами. Эти рощи состоят из мелколиственных или широколиственных пород: липа, дуб, ясень, сосна, лиственница, береза, осина.
Значение
Роль степной зоны в природе огромна. Здесь обитают уникальные виды животных и растений, которые не встречаются больше нигде.
Для хозяйственной деятельности человека травянистые просторы также имеют большое значение:
- здесь выращивается большое количество зерновых и технических культур;
- обширные пастбища для скота;
- в недрах земли содержится множество полезных ископаемых: железная руда, марганец, фосфориты, каменный уголь.
- степные озера содержат огромное количество соли, как пищевой, так и целебной.
Однако у степного земледелия есть и существенный недостаток — засушливый климат. Для защиты растений от засухи используется специальное орошение.
Для защиты растений от засухи используется специальное орошение.
Хозяйственная деятельность
Земли, покрытые степями и лесостепями являются самыми плодородными и благоприятными для выращивания различных культур: подсолнечника, кукурузы, пшеницы, бахчевых культуры. Широко развито садоводство и создание виноградников. Помимо земледелия, бескрайние поля прекрасно подходят для животноводства благодаря богатой кормовой базе.
Почвы
Почвенный покров в степной зоне — наиболее благоприятное место для выращивания культурных растений. И это неспроста, так как почвы здесь представлены черноземом. Насыщенность марганцем, железом, кальцием, магнием делает этот тип почвы самым плодородным на планете. Также в ее состав входит гумус — вещество, богатое органическими веществами. Благодаря ему чернозем окрашен в черный или темно-серый цвет.
Климат степей
Для климатических условий степной зоны характерны большие перепады температур, как сезонные, так и суточные. В Западной Сибири климат отличается большими скачками: нередко столбик термометра достигает —40 °С зимой и + 40 °С летом. Рекордно жаркая температура была зафиксирована в поселке Утта района Калмыкии в 2010 году. Столбик термометра тогда достиг +45 °С.
В Западной Сибири климат отличается большими скачками: нередко столбик термометра достигает —40 °С зимой и + 40 °С летом. Рекордно жаркая температура была зафиксирована в поселке Утта района Калмыкии в 2010 году. Столбик термометра тогда достиг +45 °С.
Среднегодовое количество осадков не превышает 200—400 мм. Дождливый период приходится на начало лета, затем наступает засуха.
Климат лесостепей характеризуется менее засушливым летом и более частыми осадками (в среднем — 600 мм).
Заповедники
Плодородные земли степных полей издавна привлекали земледельцев. Повсеместная распашка и выпас скота привели к тому, что нетронутых участков степи осталось совсем мало.
Для защиты растительного и животного мира этой уникальной экосистемы было создано несколько заповедников:
- «Оренбургский». Самый первый заповедник, созданный в России, основан в 1989 году. Сухой степной участок, граничащий с полупустынями, площадью более 21000 гектаров. Здесь охраняется множество редких видов растений и животных, среди которых степной орел, дрофа, беркут, обыкновенный бобр.
- «Черные земли». Создан в 1990 году. Под защитой находятся редкие виды растений: василек Талиева и тюльпан Шренка. Тут также обитают сайгаки, ушастые ежи, тушканчики и степные птицы вроде дрофы, жаворонка и курганника.
- «Белогорье». Созданный в 1999 году, этот заповедник расположен в лесостепной зоне, в южной части Среднерусской возвышенности. Здесь находится под охраной несколько десятков видов животных и свыше трех сотен растений.
- «Приволжская лесостепь». Еще один заповедник, под охраной которого находятся как степи, так и отдельные лесные участки. В нем произрастают редкие виды флоры, многие из которых включены в Красную книгу: четыре вида ковыля, ирис безлистный, неоттианта клобучковая, рябчик русский.
Благодаря заповедникам и национальным паркам удается сохранять многие участки степей и лесостепей в первозданном виде. Эти территории стали домом для множества животных и растений, которые встречаются только в степной зоне.
Экологические проблемы степей России
Из-за деятельности человека, а также частой засухи и сильных ветров в степях все чаще проявляются признаки опустынивания. Это означает, что некогда плодородные земли, покрытые пышной и сочной растительностью, начинают превращаться в голую высушенную пустыню.
Это означает, что некогда плодородные земли, покрытые пышной и сочной растительностью, начинают превращаться в голую высушенную пустыню.
Также на экологическое состояние степной зоны отрицательное влияние оказывают следующие факторы:
- отведение огромных площадей под посевы и урожай приводит к сокращению нетронутых земель;
- из-за перевыпаса скота почвенный покров не успевает восстановиться и дать жизнь новым степным растениям;
- частые пожары, возникающие по причине засушливого климата и неосторожности людей;
- отходы и выбросы, остающиеся от промышленной деятельности загрязняют окружающую среду;
- искусственное орошение полей приводит к засолению почв и сокращению водных ресурсов;
- бесконтрольная вырубка деревьев в лесостепях.
Чтобы защитить те нетронутые участки южных полей, которые еще сохранились, следует сократить деятельность человека в этих местах: вовремя перегонять скот на новые пастбища, ограничить количество площадей для земледелия, расширять территории заповедников.
Видео
В видео рассказывается о природе степей и лесостепей, видовом составе растительного и животного мира.
Лесостепи и степи
Природа лесостепей
Между лесной и степной зонами находится переходная лесостепная полоса, занимающая 7% территории страны или 150 млн. га.
Современные лесостепные ландшафты начали формироваться после отступления материковых ледников. Лесостепные территории до начала их освоения были покрыты лесами – в европейской части страны это были широколиственные леса, а в Западной Сибири – мелколиственные леса.
Лесостепные ландшафты в пределах Русской равнины и в Западной Сибири хорошо выражены.
Рисунок 1. Лесостепные ландшафты. Автор24 — интернет-биржа студенческих работ
По долготе зона делится на три области – Восточноевропейскую, Сибирскую, Дальневосточную.
От Карпат до Алтая лесостепь вытянута в виде сплошной полосы. Отдельными островами она представлена к востоку от Алтая.
Отдельными островами она представлена к востоку от Алтая.
Наиболее крупными лесостепными островами являются Кузнецкий, Минусинский, Ачинский, Канский, Кайтунский, Иркутский, Селенгинский, Нерчинский.
Лесостепной ландшафт на Дальнем Востоке имеют равнины Приморья и Приамурья. Есть лесостепные ландшафты на территории Крыма и Кавказа, связанные с вертикальной зональностью.
По широте лесостепь делится на северную и южную. Полевые ландшафты северной лесостепи чередуются с лесными и лугово-пастбищными.
В южной лесостепи лесных ландшафтов становится меньше.
Протяженность лесостепной полосы с севера на юг меняется в пределах от 500 до 250 км, это связано с тем, что её северная и южная границы то отступают, то приближаются друг к другу.
Причины этого связаны с климатическими факторами, рельефом местности и геологическим строением.
Климат зоны переходный от умеренно влажного лесного к засушливому степному. С запада на восток происходит увеличение континентальности, что проявляется в температурах зимнего периода и выпадающих осадках.
На западе зоны зима мягкая, умерено снежная, с январской температурой -9…-10 градусов. На востоке зоны и в Сибири зимы становятся значительно холоднее, со средней температурой января -15…-20 градусов.
На всей территории лето достаточно теплое, температура июля +20…+22 градуса.
Количество осадков меняется в направлении с запада на восток от 500 мм до 400 мм. Осадки часто носят ливневый характер, размывают грунт и способствуют почвенной эрозии.
Формирование почв происходит в условиях переменного увлажнения. На севере лесостепей преобладают серые лесные почвы, а на юге – выщелоченные и мощные черноземы. Встречаются черноземы обыкновенные.
В лесостепях азиатской части страны почвы дерново-подзолистые, обыкновенные и тучные черноземы небольшой мощности.
В Западной Сибири встречается много солонцов и солончаков.
Основная лесообразующая порода европейской лесостепи – дуб.
В Западной Сибири лесные массивы образуют березовые рощи – колки.
Современный состав лесов сильно изменен человеком. От вырубки пострадали ценные породы деревьев дуб, ясень, сосна. На смену им пришли граб, береза, осина.
От вырубки пострадали ценные породы деревьев дуб, ясень, сосна. На смену им пришли граб, береза, осина.
Лесостепь Русской равнины более теплая и влажная. В растительном покрове до распашки преобладали ковыль, овсяница, типчак, тонконог, мятлик и др.
В пределах сибирской лесостепи климат более холодный, распространены процессы заболачивания. Преобладают здесь луговые виды растений с многочисленным разнотравьем.
Лесостепные ландшафты Дальнего Востока занимают небольшую территорию.
В животном мире сочетаются представители лесостепи и степи. Таежные представители – соболь, росомаха, рысь в лесостепную зону не заходят. К востоку зоны ряд европейских видов животных – дикий лесной кот, суслик европейский – исчезают.
Природа степей
Приведем общую характеристику степи.
С продвижением к югу сокращается количество осадков и увеличивается испаряемость. Древесная растительность здесь встречается только в поймах рек – это зона степей.
Она протянулась от западной границы России до Алтайских гор на востоке. Далее к востоку степь островками вклинивается в зону тайги.
Далее к востоку степь островками вклинивается в зону тайги.
Степи на севере начинаются от южной границы лесостепей и заканчиваются в предгорьях Большого Кавказа и Крымских гор.
Степная зона занимает южную часть умеренного климатического пояса, становится значительно суше и теплее. Недостаток влаги объясняется удаленностью от океанов и редким проникновением сюда циклонов.
Лето теплое и продолжительное, зима холодная и малоснежная. Здесь часто возникают суховеи и засухи.
Средняя июльская температура +22 градуса, но в особенно жаркие дни поднимается до +40. Влажность воздуха около 50%. Дожди выпадают в виде ливней, влага не успевает пройти через почву и испаряется.
Зима холодная с температурой -30 градусов и тонким снежным покровом, но зато короткая.
Типичные степи лишены лесной растительности, за исключением речных долин.
Почвы черноземные, наибольшей тучности и мощности они достигают в северной полосе степи. Гумуса они иногда содержат до 16%. Надо сказать, что с продвижением к югу гумуса в почве становится меньше, чернозем светлеет и переходит в каштановую почву.
Надо сказать, что с продвижением к югу гумуса в почве становится меньше, чернозем светлеет и переходит в каштановую почву.
Растительный покров представлен злаками, которые растут маленькими кочками. Наиболее распространенным является обыкновенный перистый ковыль, покрывающий огромные пространства и придающий степи особый волнующий вид.
Рисунок 2. Ковыльная степь. Автор24 — интернет-биржа студенческих работ
В сухих бесплодных степях растет более мелкий ковыль.
Важную роль имеет типец, представляющий корм для овец.
Характер растительности позволяет выделить три подзоны:
- луговые степи, переходные к лесной зоне, с черноземными очень плодородными почвами и красочным разнотравьем – мятлик, костер, тимофеевка и др.;
- злаковые степи на южных черноземах и темно-каштановых почвах;
- южные полынно-злаковые степи на каштановых почвах, имеющие включения солонцов.
Луговые степи отличаются наибольшим богатством и разнообразием травянистых растений – шалфей, герань, чертополох, горицвет, лапчатка и др. До 50 видов может встречаться на 1 кв. м поверхности.
До 50 видов может встречаться на 1 кв. м поверхности.
Богата и разнообразна фауна степей, правда, под влиянием человека сильно изменившаяся. Когда-то здесь обитали дикие лошади, туры, зубры, косули, исчезнувшие ещё в прошлом веке.
Современными представителями фауны являются грызуны – суслики, тушканчики, хомяки, полевки. Из птиц – жаворонок, дрофа, стрепет.
Степь сегодня почти вся распахана и это наложило свой отпечаток на животный мир, лишив многих корма и убежищ.
На целинных и залежных степных землях животный мир более сохранился. Их обитателями являются зеленая ящерица, степная ящурка, степная гадюка, водяной уж, болотная черепаха.
Насекомых представляют луговой мотылек, полевые сверчки, прусы, саранча и др.
Экологические проблемы лесостепей и степей
Экологические проблемы появляются не сами по себе и человеческого фактора в этом вопросе не избежать.
Именно человек последовательно и настойчиво расшатывал экологический баланс и негативные последствия стали очевидными.
Изначально предназначенные для пастбищ и разведения скота, степи стали использовать не по прямому назначению. Их полностью распахали и приступили к освоению новых нужд – выращиванию кукурузы, пшеницы, сахарной свеклы.
Для этого требовалось их орошение, истребление грызунов как вредителей, использование удобрений.
Подобные действия человека привели ко второй проблеме – опустыниванию степей. Проблема тоже связана с хозяйственной деятельностью человека.
Причиной опустынивания является почвенная эрозия, являющаяся следствием высыхания рек и вырубки прилегающих лесов.
С появлением экологических проблем началось претворение в жизнь различных планов и проектов с целью обеспечения экологической сохранности степных территорий.
Для этого были приняты следующие решения:
- Экологическое зонирование оставшихся территорий степей и лесостепей;
- Составление списков редких и исчезающих представителей флоры и фауны, пополнивших Красную книгу;
- Проведение мероприятий по поддержанию плодородия почвы;
- Мероприятия по рекультивации земель, направленные на снижение негативных последствий деятельности человека.

Замечание 1
Подобных решений недостаточно и они не принесут желаемого эффекта, если не будет ответственности человека за свои действия. Ответственность и является главным фактором сохранения экологического баланса на нашей Земле.
Почвы лесостепи и степи – лесостепные и степные почвы
Почвы лесостепной и степной зон считаются самыми плодородными. Они формируются в благоприятных климатических условиях – при теплой сухой погоде и небольшом количестве осадков. Благодаря этому отмершие растительные останки быстро перегнивают. Так формируется и накапливается гумус – главное органическое вещество почвенного покрова, отвечающее за плодородие.
- Почвы лесостепи и степи
- География лесостепных и степных почв
- Условия образования лесостепных и степных почв и их особенности
- Классификация лесостепных и степных почв
- Использование лесостепных и степных почв
В этой статье мы расскажем о почвах лесостепи и степи. Вы узнаете, где они находятся, благодаря каким факторам и условиям формируются. Вы прочитаете о том, какие типы и подтипы почвенных покровов распространены на территории лесостепной и степной зон. В конце статьи мы опишем, как эти почвы используются человеком.
Вы узнаете, где они находятся, благодаря каким факторам и условиям формируются. Вы прочитаете о том, какие типы и подтипы почвенных покровов распространены на территории лесостепной и степной зон. В конце статьи мы опишем, как эти почвы используются человеком.
География лесостепных и степных почв
Лесостепь находится между лесной и степной зонами. Она считается переходной, простирается узкой полосой в южной части Русской и Западно-Сибирской равнин. Занимают лесостепи 7,5% территории России. Чем севернее местность, тем она более лесистая.
Степная зона расположилась южнее лесостепи. Она входит в состав большой Евразийской степи, которая находится в центре евразийского материка. Раньше ее также называли Великой. Она занимает действительно огромную территорию: ее начало лежит в районе Сербии и Венгрии, а конец – на Дальнем Востоке, буквально в 200-300 км от берегов Тихого океана.
Для наглядности, на карте ниже Евразийская степь выделена бирюзовым цветом:
В России степная зона охватывает 4,7% от общей площади страны.
Здесь находятся следующие степи:
- Калмыцкая (Астраханская область)
- Донская (Ростовская область)
- Ногайская (Ставропольский край, Дагестан и Чеченская республики)
- Тургайская (Оренбургская область и Казахстан)
- Ишимская (Курганская, Тюменская и Омская области в России, Северо-Казахстанская – в Казахстане)
- Курумбельская степь (Омская и Новосибирская области)
- Курайская (республика Алтай)
- Самаха (республика Алтай)
- Тажеранская (Иркутская область)
Это список не полный, его можно продолжать.
Степной район, в отличие от лесостепного, характеризуется меньшим количеством осадков и большим испарением. Именно здесь создаются благоприятные условия для формирования черноземов – самого плодородного типа почв.
Он широко распространен на территории:
- Черноземья (Воронежская, Белгородская, Курская, Липецкая и Тамбовская области)
- Поволжья (республики Калмыкия, Татарстан и Чувашия, а также Самарская, Саратовская, Пензенская, Ульяновская, Волгоградская и Астраханская области)
- Северного Кавказа (Краснодарский и Ставропольский край, республики Адыгея, Дагестан, Карачаево-Черкесия, Кабардино-Балкария, Северная Осетия, Чечня, Ингушетия)
- Южного Урала (Челябинская и Оренбургская области, республика Башкортостан)
- Западной Сибири (Тюменская, Курганская, Томская, Новосибирская, Омская, Кемеровская области, Алтайский край и республика Алтай)
 Они расположились на крошечной площади региона. Но по своим качественным характеристикам местные черноземы заметно уступают покровам вышеназванных территорий. Об этом вы можете прочитать в нашей статье Почвы Свердловской области.
Они расположились на крошечной площади региона. Но по своим качественным характеристикам местные черноземы заметно уступают покровам вышеназванных территорий. Об этом вы можете прочитать в нашей статье Почвы Свердловской области.Обычно в этой зоне выделяют еще одну – зону сухих степей. Она занимает 1,7% территории России. Тянется эта область от Прикавказья до Алтая, через Среднюю Сибирь и Забайкалье.
Если говорить о мире в целом, то там лесостепные и степные зоны встречаются на всех континентах, кроме Антарктиды. Наиболее распространены они в восточной Европе, в таких странах как Украина, Молдавия, Румыния, Болгария. Кроме того, эти почвы встречаются на территории Казахстана, Монголии, Китая, а также Северной и Южной Америк. Их положение вы можете увидеть на карте ниже:
Лесостепные и степные зоны считаются самыми изученными и освоенными. Советский ученый В.В. Докучаев, основоположник почвоведения, начал исследование почвенных покровов СССР именно с этих мест. Его интересовало, почему черноземные почвы дают такой хороший урожай, какие факторы этому способствуют.
Условия образования лесостепных и степных почв и их особенности
Факторы и условия формирования почв в лесостепи, степи и сухой степи:
- Климат
Климат в лесостепи умеренно-теплый, в июле температура колеблется от +18°С до +25°С, в январе – от -4°С до -25°. Период вегетации растений – от 90 до 180 дней. Западная часть лесостепи более теплая и влажная.
Степная зона характеризуется более сухим климатом. На протяжении года плюсовая температура здесь сохраняется около 200 дней.
Климат в зоне сухих степей континентальный и сухой, зимы холодные и бесснежные, а лето жаркое. Вегетационный период растений длится 200-210 дней. - Водный режим
В лесостепной зоне чередуется промывной и непромывной водный режимы. Осадков выпадает от 300-350 мм в год на востоке до 550-700 на западе. Весной и осенью влага с поверхности достигает грунтовых вод, летом и зимой нет. Существует баланс между поступлением влаги и ее испарением. Летом и зимой в почве замедляются процессы распада органики, интенсивно накапливается гумус. Все эти обстоятельства способствуют тому, что покровы в лесостепи обладают очень высоким плодородием, одним из лучших на планете.
Весной и осенью влага с поверхности достигает грунтовых вод, летом и зимой нет. Существует баланс между поступлением влаги и ее испарением. Летом и зимой в почве замедляются процессы распада органики, интенсивно накапливается гумус. Все эти обстоятельства способствуют тому, что покровы в лесостепи обладают очень высоким плодородием, одним из лучших на планете.
В степи ежегодно выпадает 200-500 мм осадков. Испарение тут превышает поступление влаги. Режим почвы непромывной.
В зоне сухих степей за год выпадает 200-250 мм осадков на юге, 350-400 мм – на севере. Испарение влаги здесь в 2-4 раза превышает ее поступление. - Материнская порода
Почвообразующими породами лесостепных почв являются суглинок, глина, лёссовая глина с большим содержанием пылевидных частиц. В них много кальция, поэтому реакция плодородных покровов близка к нейтральной.
Почва степной зоны образуется на лёссах и лёссовых суглинках с высоким содержанием карбонатов и растворимых солей. Ее реакция нейтральная или слабощелочная.
Ее реакция нейтральная или слабощелочная.
Материнская порода сухих степей представлена карбонатными суглинками и лёссами. В таких условиях часто развиваются солончаки. Они характерны для низин, где около поверхности стоят обогащенные растворимыми солями грунтовые воды. Мы подробно рассказывали об этом в нашей статье Засоленные почвы.
Подробно об этом факторе вы можете прочитать в нашей статье Почвообразующая порода как фактор почвообразования. - Растительность
Основная растительность в лесостепи – широколиственные леса и луга. Они дают 7-9 т опада в год, при этом половина его образуется корнями травянистых растений. Органические вещества от корней поступают непосредственно в почву. Опад богат белком и азотом, за год в почву возвращается до 60-90 кг этого ценного элемента и почти 250-300 кг других минералов.
Растительность степи травянистая, с единичными деревьями и кустарниками. В зоне сухих степей она представлена ковылем, типчаком и полынью. Количество опада скудное – до 2-4 т/га.
Подробно об этом факторе вы можете прочитать в нашей статье Биологический фактор почвообразования.
Все эти факторы поспособствовали тому, что в степи и лесостепи образовались самые плодородные почвенные покровы.
Среди особенностей этих почв выделяют:
- Высокую степень плодородия за счет большого насыщения гумусом
- Мощный гумусовый горизонт, который может достигать 1 м
- Слабую минерализацию растительных останков
- Высокое содержание перегноя
- Рыхлую структуру благодаря травянистой растительности и пронизывающим почву корням
Далее мы расскажем, какие типы и подтипы почв встречаются на территории лесостепей и степей.
Классификация лесостепных и степных почв
К почвам лесостепной, степной и сухой зон относятся следующие типы и подтипы:
- Бурые лесные (буроземы) и бурые лесные глеевые
- Подзолисто-бурые лесные (в том числе глеевые)
- Серые лесные (светлые и темные; глеевые)
- Черноземы (типичные, оподзоленные, выщелоченные)
- Сероземы
- Каштановые (светлые и темные)
- Луговые
- Лугово-черноземные
- Лугово-болотные
- Лугово-каштановые
- Луговые подбелы (лугово-бурые)
- Коричневые
- Лугово-коричневые
- Лугово-лесные серые
- Лугово-серо-коричневые
Для вашего удобства, все основные данные об этих почвах, кроме черноземных, мы собрали в одну сводную таблицу. В ней мы разместили всю основную информацию о каждом типе и подтипе.
В ней мы разместили всю основную информацию о каждом типе и подтипе.
В таблице указано:
- Где формируются те или иные почвенные покровы
- Какие условия и факторы особенно важны в формировании этих почв
- Какова степень плодородия каждого почвенного типа и подтипа
- В чем заключаются главные особенности почв лесостепной и степной зон
- Как используют тот или иной почвенный покров, какие меры необходимо принимать для улучшения качественных характеристик почв и их степени плодородия
Все эти данные помогут вам составить достаточно полное представление о каждом типе и подтипе.
Классификация лесостепных и степных почв
Как можно заметить из таблицы, все почвы лесостепной и степной зон активно используют в сельском хозяйстве. На них выращивают всевозможные культуры, от злаковых до корнеплодов. Ниже мы также подобрали для вас несколько иллюстраций, на которых видны профили разных степных и лесостепных покровов. Слева направо – буроземы, каштановые, лугово-каштановые и лугово-болотные почвы:
Слева направо – буроземы, каштановые, лугово-каштановые и лугово-болотные почвы:
Особняком от этих почв стоит чернозем. О нем мы бы хотели поговорить чуть подробнее.
Чернозем
Черноземные почвы обладают самым высоким плодородием. Они образовались на территориях с развитой травянистой растительностью. За год в покров попадает около 10-20 т растительных останков, при этом 40-60% приходится на корни. Интенсивное разложение органики идет весной и в самом начале осени. Летом и зимой процессы приостанавливаются, что благоприятно сказывается на процессе образования гумуса.
На протяжении года в землю попадает такое же количество осадков, что и испаряется. Промывной режим отсутствует. То есть влага из верхних слоев не попадает в подземные воды, питательные вещества не вымываются.
Образуются черноземы на породах, которые богаты кальцием. Его соли с влагой поднимаются в гумусовый слой. В травяном опаде много азота и оснований, в гумусе гуминовые кислоты преобладают над фульвокислотами. Поэтому реакция почвы нейтральная или слабощелочная, в редких случаях слабокислая.
Поэтому реакция почвы нейтральная или слабощелочная, в редких случаях слабокислая.
Основные признаки черноземов:
- Мощный гумусовый горизонт (от 40 см до 1 м)
- Слабая дифференциация профиля, практически отсутствует горизонт выноса (элювиальный)
- В горизонте вноса (иллювиальном) сконцентрированы карбонаты
- Верхние слои зернистые или зернисто-комковые, нижние слои – крупнокомковые
В лесостепной зоне выделяют такие подтипы черноземов:
- Оподзоленные
Они образовались во влажных широколиственных лесах. Карбонатный слой залегает на глубине 1-1,5 м. В гумусовом горизонте выявляются участки с кремнеземом белого цвета, свободные карбонаты. Реакция почвы бывает слабокислой и даже кислой. - Выщелоченные
Они образовались в более сухих лесах и степях. Плотный слой карбонатов залегает на глубине 60-80 см, в иллювиальном горизонте. Он лишь изредка поднимается в верхние слои. Реакция грунта слабокислая. - Типичные
Почвы отличаются самой большой мощностью гумусового слоя (около 1-2 м) и высоким плодородием. В почве практически отсутствуют фульвокислоты. Карбонатный слой залегает на глубине 60-70 см. Реакция покрова нейтральная. Цвет у него черный, она хорошо поглощает тепло.
Черноземы лесостепной зоны – это самые ценные грунты сельского хозяйства. На них можно собирать большие урожаи после внесения минимального количества удобрений. Единственный недостаток таких грунтов – дефицит влаги. Поэтому возле полей высаживают лесополосы для задержки снега и осадков, оборудуют систему полива.
В степной зоне также образуются черноземы, но их плодородие ниже, чем у лесостепных. Это связано с низкой влажностью, высоким содержанием солей в материнской породе. В некоторых местах это приводит к образованию солончаков.
Черноземы степной зоны:
- Обыкновенные
Почвы распространены в северной части зоны, в довольно засушливых условиях. Растительность здесь развита слабее, чем в лесостепи. Мощность гумусового слоя едва достигает 50 см. Он обеднен гуминовыми кислотами, имеет коричневый цвет. Из материнской породы в верхние горизонты попадают белые зернистые включения – соли кальция. Структура почвы комковая.
Растительность здесь развита слабее, чем в лесостепи. Мощность гумусового слоя едва достигает 50 см. Он обеднен гуминовыми кислотами, имеет коричневый цвет. Из материнской породы в верхние горизонты попадают белые зернистые включения – соли кальция. Структура почвы комковая. - Южные
На юге степной зоны в год выпадает 350-400 мм осадков, большая их часть испаряется. Растительность здесь довольно скудная, количество опада меньше. В материнских породах содержится много кальция и растворимых солей. В иллювиальном горизонте нередко появляются прослойки гипса.
Южные черноземы светлее, чем обыкновенные. В них четко видны вкрапления кальцитов. Толщина гумусового слоя 40-60 см, его структура зернисто-комковая, иногда уплотненная.
Для вашего удобства, ниже мы составили таблицу распространения черноземных почв. В ней наглядно показано, в каких ландшафтных зонах и подзонах встречается тот или иной подтип.
В Свердловской области встречается два подтипа черноземных почв:
- Выщелоченные
- Оподзоленные
Подробно об этом вы можете прочитать в нашей статье Почвы Свердловской области.
Возможно, вам также будут интересны наши статьи по теме:
- Характеристики и свойства чернозема – подробно о всех свойствах черноземных почв, которые даны в ГОСТах
- Как выглядит чернозем – по каким внешним признакам можно узнать черноземную почву
- Как определить качество чернозема – как можно установить качество материала на разных этапах заказа (от звонка до выгрузки)
- Чем богат чернозем, его плюсы и минусы – все о преимуществах и недостатках этой почвы
- Применение чернозема – где и для чего используется этот покров
В завершающей части статьи мы расскажем, как и для каких нужд лесостепные и степные почвы осваиваются человеком.
Использование лесостепных и степных почв
Благодаря своим качественным характеристикам и высокому плодородию, почвы лесостепи и степи активно обрабатываются человеком. Их используют при занятии земледелием уже многие годы. Они заняты различными сельскохозяйственными угодьями, пашнями, сенокосами. Кроме того, степные покровы могут использоваться в качестве пастбищ.
Кроме того, степные покровы могут использоваться в качестве пастбищ.
На почвах лесостепных и степных зон выращивают всевозможные культуры, от зерновых до плодово-ягодных. Здесь хорошо растут различные злаки и зерновые, сахарная свекла, табак, подсолнечник, овощные и бахчевые культуры, виноград.
Почвы лесостепной и степной зон возделываются уже многие десятилетия. Все это не могло не сказаться на их качественных характеристиках – во всем мире сейчас наблюдается деградация окультуренных черноземов. Ведь при земледелии нарушается природный оборот биологических веществ, изменяется почвенная микрофлора, уменьшается толщина гумусового слоя. Все это приводит к падению показателя плодородности.
Для сохранения, в первую очередь, ценных черноземных покровов, рекомендуется проводить ряд агротехнических мероприятий.
В их числе:
- Высаживание многолетних трав, бобовых, растений-сидератов
- Внесение органических и минеральных (в основном азотных и фосфорных) удобрений
- Известкование
- Глубокая вспашка
- Правильное орошение
- Высаживание лесополос для задержки влаги
- Содержание почв под паром
- Борьба с эрозией
- Осушительная мелиорация (при необходимости)
С почвами в степной и лесостепной зонах нужно обращаться очень бережно, проводить различные мероприятия по повышению их качественных характеристик. Ведь в случае неверного использования или ухода за покровами, можно лишиться огромных пахотных территорий. Так, например, неправильная система полива может привести к засолению почв. Также покровам может навредить чрезмерное внесение минеральных удобрений. А вот органические удобрения, наоборот, восстанавливают нормальную микрофлору почвы, стимулируют процесс образования гумуса.
Ведь в случае неверного использования или ухода за покровами, можно лишиться огромных пахотных территорий. Так, например, неправильная система полива может привести к засолению почв. Также покровам может навредить чрезмерное внесение минеральных удобрений. А вот органические удобрения, наоборот, восстанавливают нормальную микрофлору почвы, стимулируют процесс образования гумуса.
растительный и животный мир, географическое положение
Бескрайние просторы, залитые ярким солнцем и поросшие густой травой — все это представляет собой степь, практически лишенная деревьев, за исключением берегов рек и водоемов. Чуть севернее, ближе к лесной зоне, начинаются лесостепи. Здесь уже к травянистой равнине начинают добавляться островки лесов и небольших рощ.
Лесостепи и степи играют огромную роль в деятельности человека, потому что почвы в этих местах отличаются наилучшей плодородностью.
Что такое природные зоны
В первую очередь следует дать определение самому понятию природных зон: это территория, для которой характерны определенные климатические условия, рельеф, почва, животный и растительный мир.
На планете существуют следующие территориальные экосистемы:
- арктическая пустыня;
- тундра и лесотундра;
- тайга;
- смешанные леса и широколиственные леса;
- степи и лесостепи;
- пустыни и полупустыни;
- жестколистные леса и кустарники;
- саванны и редколесья;
- экваториальные тропические леса.
Все биомы сформировались на основе климатических поясов: Арктика находится в арктическом поясе, тундра и лесотундра — в субарктическом, лесная зона и степи — в умеренном, жестколистные леса — в субтропическом, пустыни и полупустыни — в тропическом, саванны и редколесья — в субэкваториальном, тропические леса — в экваториальном.
В таблице приведен перечень основных природных зон и их отличительные особенности.
Арктические Пустыни Евразии
Арктические пустыни расположены на арктических островах: Новосибирские, Шпицберген, Северная Земля, Врангеля и Новая Земля, а также в Азии на берегу Северного ледовитого океана.
Такие зоны считаются наиболее суровыми природными зонами для проживания. Растительность очень скудная. Только в летний период на камнях растут лишайники. Единственным ресурсом для пропитания животных является море. В нём они добывают еду. Там обитают тюлени, моржи и белые медведи.
На юге Арктики располагается полоса тундры, пересекающая континент от запада к востоку.
Эта зона характеризуется субарктическим климатом с затяжной зимой. Воздух может охлаждаться до отметки -30-40°С. Снега в этой местности не бывает много. Летом температура не поднимается выше 10°С.
Через территорию Европы проходит наиболее узкая часть тундры. Лесов в этих зонах нет. Основная растительность проявляется лишайниками, мхами и ягодными кустарниками, такими как голубика, брусника или черника. Деревья встречаются в основном карликовых форм.
Животный мир в тундре отличается огромным видовым разнообразием. В этой зоне встречаются самые различные виды животных: как многочисленные грызуны, так и крупные парнокопытные, и даже хищники.
Многие из них на время холодов перемещаются в южные районы тундры (лесотундра, тайга).
Степи и лесостепи
Слово «степь» не имеет точного происхождения. Предполагается, что оно означает «вырубленное место», то есть лишенное деревьев. По мнению других, название связано со словом «топтать», иными словами, «вытоптанное место».
У разных стран данное понятие имеет свое название: в Южной Америке — пампасы, в Северной Америке — прерии, в Новой Зеландии — туссоки.
Географическое положение
Степная зона встречается практически на всех континентах. Североамериканские прерии покрывают центральную часть США, север Мексики и запад Канады. Пампасы Южной Америки расположены в юго-восточной части, между Андами и Атлантическим океаном. В Евразии травянистые равнины занимают обширные площади Монголии, Казахстана, Узбекистана, южной части России, Украины.
В России зона степей расположилась на юге Восточно-Европейской и Западно-Сибирской равнин. Отдельные участки встречаются в Забайкалье. С севера на юг степной регион протянулся от Тулы и рек Кама и Белая до гор Кавказа.
С севера на юг степной регион протянулся от Тулы и рек Кама и Белая до гор Кавказа.
Лесостепи узкой полосой тянутся вдоль степных полей, отделяя их от лесных массивов.
Климат
Степи расположены в умеренном и субтропическом климате. Для этих мест характерна континентальность, то есть удаленность от морских и океанических побережий. Из этого следует, что лето здесь жаркое и засушливое, а зимнее время — холодное и малоснежное.
Климатические условия различаются в зависимости от местоположения степи. Заволжские районы являются переходной территорией между западными влажными степями и сухими континентальными областями на востоке. В районе Крыма климат становится более мягким и влажным: в зимнее время температура редко опускается ниже 0°С, а летом в среднем составляет +22°С. Однако для этих районов характерны суховеи.
В лесостепях климат более мягкий: зимой показатели температуры составляют от -2 до -20°С.
Животный мир
Травянистые равнины населены разнообразными животными и птицами. В основном здесь живут травоядные, которым степь дает все необходимое для жизни. Но, как и в любой экосистеме, тут обитают и хищники.
В основном здесь живут травоядные, которым степь дает все необходимое для жизни. Но, как и в любой экосистеме, тут обитают и хищники.
Что касается лесостепей, то для них характерны как обитатели степей, так и лесные жители.
Степной волк
От обычных представителей своего вида отличается более мелкими размерами и светлой, желтовато-серой шерстью. Охотятся эти хищники на степных копытных (сайгаки и лошади), но могут питаться и более мелкой добычей: кроликами, зайцами, птицами, грызунами. Степные волки предпочитают селиться на берегах рек или в балках.
Корсак
Корсак, или степная лисица, обитает в норах, которые остались после барсуков, сусликов и других животных. Корсак охотится на грызунов и птиц, а зимой уходит на юг в поисках пропитания. Основными врагами лисицы являются волки, хищные птицы и человек, охотящийся на корсака ради пушистого меха.
Манул
Представитель семейства кошачьих, обладающий очень пушистым и густым мехом. Его шерсть окрашена в светло-серые, с рыжеватым оттенком, цвета. Этот дикий кот обитает в степях и лесостепях Центральной и Средней Азии. Может населять и горы в этих регионах. Ведет ночной образ жизни, питаясь различными грызунами, птицами и зайцами.
Этот дикий кот обитает в степях и лесостепях Центральной и Средней Азии. Может населять и горы в этих регионах. Ведет ночной образ жизни, питаясь различными грызунами, птицами и зайцами.
Сайгак
Эта уникальная разновидность антилопы обитает в степях Евразии. Отличительная особенность сайгака — большой нос, похожий на короткий хобот. Степные антилопы ведут кочевой образ жизни, постоянно перемещаясь в поисках новых пастбищ. Стадо сайгаков может насчитывать до нескольких сотен голов.
Тушканчик
Небольшой грызун, имеющий сильные задние лапы, на которых и перемещается. Также его характерной чертой является длинный хвост с большой кисточкой на конце. Тушканчики ведут ночной образ жизни, днем прячась в норах. Питаются побегами растений и семенами.
Степной орел
У этой птицы очень острое зрение, позволяющее ей высматривать добычу с большой высоты. Пропитанием орлу служат суслики, мелкие птицы, ящерицы, змеи. Окрас оперения темно-бурый, на крыльях имеются поперечные серые полосы.
Дрофа
Крупная птица, обитающая в ковыльных и полынных степях. Дрофа имеет пестрый окрас, состоящий из белых, серых, черных и рыжих перьев. Питается насекомыми, грызунами, ящерицами. Может употреблять в пищу побеги и семена. В настоящее время данный вид находится под угрозой.
Желтобрюхий полоз
Крупных размеров змея, чешуя которой окрашена в черный цвет, а низ, соответственно названию, имеет желтоватый оттенок. Данный вид змей не ядовит, хотя ее укусы могут причинить сильную боль. Питается грызунами, ящерицами и птицами. На зиму впадает в спячку.
Растения
В степях произрастают в основном травы и кустарники. Деревья, ввиду засушливости территории, встречаются только по берегам рек и в лесостепной зоне. Характеристика главных представителей фауны представлена ниже.
Ковыль
Злаковое растение, имеющее на вершинах стеблей длинные светлые колоски. Поле, заросшее ковылем, кажется похожим на зелено-серебристый океан. Имеет слаборазвитую корневую систему, вследствие чего не произрастает на луговых полях с хорошей дерниной. Зато ковыль весьма устойчив к засухе, поэтому может встретиться на выжженных участках или сенокосах.
Зато ковыль весьма устойчив к засухе, поэтому может встретиться на выжженных участках или сенокосах.
Шалфей степной
Шалфей покрывает луга и поля в степной и лесостепной зоне. Имеет соцветия из сиреневых и фиолетовых цветков. Это растение широко применяется в медицине и косметологии благодаря своим противовоспалительным и очищающим свойствам.
Мятлик луговой
Является растением из семейства злаковых, имеет длинные метелки с множеством колосков. Обладает хорошей устойчивостью к морозам и засухе. Мятлик высоко ценится в качестве пастбищного растения благодаря питательным веществам и витаминам.
Касатик безлистный
Произрастает в зарослях кустарников или по берегам рек. Виды этого растения имеют еще и другие названия — ирис безлистный или касатик вильчатый. Они произошли из-за того, что листья ириса опадают зимой, а весной появляются позднее цветоносов. Высота может достигать полуметра. Касатик — растение с красивыми крупными цветками фиолетового цвета с желтой серединой. Считается редким видом и занесен в Красную книгу.
Считается редким видом и занесен в Красную книгу.
Василек синий
Представляет собой сорняковое растение и часто соседствует с посевами ржи. Тонкие стебли украшены синими цветками, растущими поодиночке. Семена василька обладают лекарственными свойствами, их порошок используют как противовоспалительное средство.
Вишня степная
От других представителей своего вида отличается небольшой высотой. Цветки, как и у обычной вишни, белого цвета. Плоды представляют собой костянку темно-красного оттенка и имеют кисло-сладкий вкус. Любит ярко-освещенные места, устойчива к морозам и засушливой погоде.
В лесостепях участки полей и лугов чередуются с небольшими лесами. Эти рощи состоят из мелколиственных или широколиственных пород: липа, дуб, ясень, сосна, лиственница, береза, осина.
Географическое положение
Лесостепь протянулась непрерывной полосой по югу Русской и Западно-Сибирской равнин. Южнее этой зоны простираются степи. Они тянутся неширокой сплошной полосой на юге России от западных границ до Алтая. Далее к востоку имеют очаговое распространение.
Южнее этой зоны простираются степи. Они тянутся неширокой сплошной полосой на юге России от западных границ до Алтая. Далее к востоку имеют очаговое распространение.
Это интересно: Каким образом использует рога снежный баран?
Значение
Роль степной зоны в природе огромна. Здесь обитают уникальные виды животных и растений, которые не встречаются больше нигде.
Для хозяйственной деятельности человека травянистые просторы также имеют большое значение:
- здесь выращивается большое количество зерновых и технических культур;
- обширные пастбища для скота;
- в недрах земли содержится множество полезных ископаемых: железная руда, марганец, фосфориты, каменный уголь.
- степные озера содержат огромное количество соли, как пищевой, так и целебной.
Однако у степного земледелия есть и существенный недостаток — засушливый климат. Для защиты растений от засухи используется специальное орошение.
Тайга
Интересно:
Симбиоз — виды, примеры, описание, фото и видео
Интересный факт: в России тайга является наиболее обширной по площади зоной (62%), а формироваться она начала еще до образования ледниковых масс.
Географическое положение и рельеф
Тайга охватывает всю Сибирь – она растянулась от Балтийского моря до берегов Тихого океана. На юге граница проходит вблизи Нижнего Новгорода, Екатеринбурга, Читы и т.д. Местность сохранила максимально естественный облик, поскольку мало подвергалась воздействию человека. Рельеф преимущественно равнинный.
Почвы и климат
Множество озер, рек и других водоемов. В тайге количество осадков сильно колеблется в течение года и может составлять от 200 до 1000 мм (самый высокий показатель среди всех зон). Почва подзолистая с высоким уровнем гумуса, минералов.
Лето теплое, а зима холодная. Тепло держится около 4-5 месяцев с температурой +16℃. Зимой термометр показывает от -10 до -50℃. В целом климат можно охарактеризовать, как стабильный.
В целом климат можно охарактеризовать, как стабильный.
Флора и фауна
Преобладают сосны, ели, пихты и кедры. Распространена луговая растительность. В сравнении с северными зонами, здесь кипит жизнь. Разнообразный животный мир, представленный птицами, млекопитающими, земноводными:
- рыси;
- волки;
- медведи;
- рябчики;
- глухари.
Хозяйственная деятельность
Земли, покрытые степями и лесостепями являются самыми плодородными и благоприятными для выращивания различных культур: подсолнечника, кукурузы, пшеницы, бахчевых культуры. Широко развито садоводство и создание виноградников. Помимо земледелия, бескрайние поля прекрасно подходят для животноводства благодаря богатой кормовой базе.
Почвы
Почвенный покров в степной зоне — наиболее благоприятное место для выращивания культурных растений. И это неспроста, так как почвы здесь представлены черноземом. Насыщенность марганцем, железом, кальцием, магнием делает этот тип почвы самым плодородным на планете. Также в ее состав входит гумус — вещество, богатое органическими веществами. Благодаря ему чернозем окрашен в черный или темно-серый цвет.
Также в ее состав входит гумус — вещество, богатое органическими веществами. Благодаря ему чернозем окрашен в черный или темно-серый цвет.
Редкие млекопитающие
Среди краснокнижных млекопитающих Алтайского края есть представители следующих отрядов: рукокрылые, насекомоядные, зайцеообразные, хищные, парнокопытные и грызуны. Всего под особой охраной находятся 23 вида.
Остроухая ночница
В длину эта летучая мышь вырастает до 7-8 см, а вместе с хвостом — достигает 13 см. Размах её крыльев составляет 40 см. Тело покрыто длинным, неровным мехом светло-бурого цвета на спинке и серо-белесого на брюшке. Поедает насекомых, многих из которых ловит прямо на лету.
Степная пищуха
Самая маленькая из пищух, обитающих в России, в длину не превышает 20-21 см. Имеет коротенькие лапки и хвост, который снаружи не виден. Живёт в норах, роет сразу несколько штук для постоянного проживания и временные, чтобы прятаться от врагов.
Серый сурок
В длину вместе с хвостом достигает 78-80 см. Имеет длинную мягкую шерсть песочного цвета на спинке и коричневатого — на брюшке. Водится в основном в горах, роет сложные норы с несколькими выходами. Питается сочным побегами молодых растений, листьями и цветами.
Имеет длинную мягкую шерсть песочного цвета на спинке и коричневатого — на брюшке. Водится в основном в горах, роет сложные норы с несколькими выходами. Питается сочным побегами молодых растений, листьями и цветами.
Выдра
Крупное животное, вырастающее в длину вместе с хвостом до 1,5 метров. Живёт в реках, озерах и прудах, устраивая жилище в зарослях у воды. Основу рациона составляют рыба и моллюски. Летом выдра охотится на мелких грызунов, лягушек и ящериц, может нападать на птиц.
Перевязка
Напоминает хорька, но намного меньше его по размеру. Длина взрослой особи не превышает 60 см с учётом хвоста. Перевязки живут поодиночке. Днем сидят в норах, ближе к ночи выходят на охоту. Едят в основном грызунов, птиц и насекомых.
Кабарга
Оленевидное животное крупного размера: 1 м в длину, 70 см в высоту, около 18 кг по весу. Имеет непропорционально длинные задние ноги, рога отсутствуют. Самцы снабжены острыми выступающими наружу клыками 7-9 см, которые они используют во время боев друг с другом. Кабарги прекрасно прыгают, умеют запутывать следы. Питаются различными лишайниками.
Кабарги прекрасно прыгают, умеют запутывать следы. Питаются различными лишайниками.
Сибирская белозубка
Маленькая землеройка с носовым отделом в виде хоботка. Вырастает вместе с хвостом до 12 см. На спинке мех тёмного бурно-серого цвета, на брюшке серовато-белый. Ведёт преимущественно ночной образ жизни. Питается насекомыми и их личинками.
Краснощекий суслик
Вырастает до 28 см, имеет короткий хвост — до 65 мм. Отличительный признак этого вида — красноватые или рыжие пятна на щеках. Проживают суслики колониями в норах, которые по глубине могут достигать 3,5 метров. Поедают злаковые культуры, чем очень вредят сельскому хозяйству.
Также в красную книгу Алтайского края включены следующие виды млекопитающих: ушастый ёж, крупнознубая бурозубка, прудовая ночница, восточная ночница, сибирская ночница, ночница Иконникова, длиннохвостая ночница, ушан Огнева, рыжая вечерница, северный кожанок, двухцветный кожан, сибирский трубконос, белка-летяга, большой тушканчик, мохноногий тушканчик.
Климат степей
Для климатических условий степной зоны характерны большие перепады температур, как сезонные, так и суточные. В Западной Сибири климат отличается большими скачками: нередко столбик термометра достигает —40 °С зимой и + 40 °С летом. Рекордно жаркая температура была зафиксирована в поселке Утта района Калмыкии в 2010 году. Столбик термометра тогда достиг +45 °С.
Среднегодовое количество осадков не превышает 200—400 мм. Дождливый период приходится на начало лета, затем наступает засуха.
Климат лесостепей характеризуется менее засушливым летом и более частыми осадками (в среднем — 600 мм).
Тундра
Тундровая зона граничит с арктической пустыней (площадь – 7% от всей территории). Ее обычно делят на 3 части: арктическую, среднюю и южную. Для тундры характерно отсутствие лесов, множество болот, торфяников, многолетняя мерзлота.
Географическое положение и рельеф
Занимает Камчатку, Урал, Кольский полуостров и составляет 7% площади РФ. Выделяют также горную тундру, которая свойственна и нашей стране. Если арктическая часть более равнинная, то горная совмещает в себе и равнины, и возвышенности.
Выделяют также горную тундру, которая свойственна и нашей стране. Если арктическая часть более равнинная, то горная совмещает в себе и равнины, и возвышенности.
Почвы и климат
По сравнению с арктической пустыней почва тундры более плодородная. Однако большинству растений все равно сложно приспособиться к таким условиям. Зима длится намного дольше, чем лето: 9 морозных месяцев против 1-3 теплых, когда сходит снег. Температура в среднем около +12℃ летом, но зимы гораздо теплее, чем в арктической пустыне – около -17℃.
Интересный факт: полярные дни и ночи свойственны именно тундровой зоне. Полярная ночь продолжается на протяжении декабря-февраля, а полярный день начинается в июле.
Флора и фауна
Помимо мхов, лишайников, здесь растут карликовые деревья. Общее для местных растений – низкорослость. Животный мир представляют белые медведи, северные олени, беляки, лоси, песцы и др.
Интересно:
Первые растения на Земле — фото и видео
Заповедники
Плодородные земли степных полей издавна привлекали земледельцев. Повсеместная распашка и выпас скота привели к тому, что нетронутых участков степи осталось совсем мало.
Повсеместная распашка и выпас скота привели к тому, что нетронутых участков степи осталось совсем мало.
Для защиты растительного и животного мира этой уникальной экосистемы было создано несколько заповедников:
- «Оренбургский». Самый первый заповедник, созданный в России, основан в 1989 году. Сухой степной участок, граничащий с полупустынями, площадью более 21000 гектаров. Здесь охраняется множество редких видов растений и животных, среди которых степной орел, дрофа, беркут, обыкновенный бобр.
- «Черные земли». Создан в 1990 году. Под защитой находятся редкие виды растений: василек Талиева и тюльпан Шренка. Тут также обитают сайгаки, ушастые ежи, тушканчики и степные птицы вроде дрофы, жаворонка и курганника.
- «Белогорье». Созданный в 1999 году, этот заповедник расположен в лесостепной зоне, в южной части Среднерусской возвышенности. Здесь находится под охраной несколько десятков видов животных и свыше трех сотен растений.
- «Приволжская лесостепь». Еще один заповедник, под охраной которого находятся как степи, так и отдельные лесные участки.
 В нем произрастают редкие виды флоры, многие из которых включены в Красную книгу: четыре вида ковыля, ирис безлистный, неоттианта клобучковая, рябчик русский.
В нем произрастают редкие виды флоры, многие из которых включены в Красную книгу: четыре вида ковыля, ирис безлистный, неоттианта клобучковая, рябчик русский.
Благодаря заповедникам и национальным паркам удается сохранять многие участки степей и лесостепей в первозданном виде. Эти территории стали домом для множества животных и растений, которые встречаются только в степной зоне.
Растения
В идеале лесостепь представляет собой травянисто-древесное пространство, а степь — полностью травянистое, но таких природных зон в России почти не осталось (деревья вырублены, а земли распаханы). Участки степи, не затронутые хозяйственной деятельностью людей (главным образом в Сибири), заселены большим числом злаков: ковылем, типчаком и тонконогом, а также ярко цветущим разнотравьем (ковылем, ирисами). В более засушливых южных районах распространены полынь, грудница, лапчатка. Весной много тюльпанов. В азиатской части страны преобладают пижма и злаки.
В азиатской части страны преобладают пижма и злаки.
Луговые степи — редкое для России явление. Большая их часть распахана
В зоне степных черноземов расположен Краснодарский край — главная житница России. Здесь собирают не только богатый урожай зерновых, но и выращивают арбузы, подсолнечник, виноград и другие теплолюбивые культуры
Экологические проблемы степей России
Из-за деятельности человека, а также частой засухи и сильных ветров в степях все чаще проявляются признаки опустынивания. Это означает, что некогда плодородные земли, покрытые пышной и сочной растительностью, начинают превращаться в голую высушенную пустыню.
Также на экологическое состояние степной зоны отрицательное влияние оказывают следующие факторы:
- отведение огромных площадей под посевы и урожай приводит к сокращению нетронутых земель;
- из-за перевыпаса скота почвенный покров не успевает восстановиться и дать жизнь новым степным растениям;
- частые пожары, возникающие по причине засушливого климата и неосторожности людей;
- отходы и выбросы, остающиеся от промышленной деятельности загрязняют окружающую среду;
- искусственное орошение полей приводит к засолению почв и сокращению водных ресурсов;
- бесконтрольная вырубка деревьев в лесостепях.

Чтобы защитить те нетронутые участки южных полей, которые еще сохранились, следует сократить деятельность человека в этих местах: вовремя перегонять скот на новые пастбища, ограничить количество площадей для земледелия, расширять территории заповедников.
Почвы
Главная почва степи — плодородный чернозем. Почвы северных степей содержат 8–10% гумуса, что является прекрасным показателем. К югу климат степей становится более сухим — содержание гумуса падает до 6%, а на самом юге, на границе с полупустынями, постепенно появляются каштановые почвы, где гумуса всего 4%. Вокруг озер нередко встречаются засоленные почвы: солоди, солонцы и солончаки.
Главное отличие лесостепей от степей — наличие участков древесной растительности, которые чередуются с обширными травянистыми пространствами. В степях для развития древесной растительности не хватает влаги
Урок географии 8 класс по теме «Природа лесостепей и степей»
Урок № 49
Тема урока: Природа
лесостепей и степей.
Цели урока:
Изучить особенности компонентов лесостепей, степей.
Изучить взаимосвязи между компонентами этих зон.
Сформировать образ лесостепей, степей.
Формировать умение давать характеристику природной зоны.
Выявить природные ресурсы этих зон и их хозяйственное использование, экономические проблемы и пути их решения.
Планируемые результаты обучения:
Предметные:
Формирование знаний о природных зонах нашей страны — лесостепной и степной.
Метапредметные:
Формирование умений:
— работать в группе;
— создавать образы степи и лесостепи;
— отбирать и анализировать необходимую информацию, делать выводы.
Личностные:
Формирование экологического мышления; осознание необходимости охраны степных ландшафтов.
Основное
содержание:
лесостепи и степи, особенности лесостепной и степной зон, лесостепи и степи —
главный сельскохозяйственный район страны.
Деятельность учащихся:
Сравнивать географическое положение степи и лесостепи.
Выявлять причинно-следственные связи между географическим положением, особенностями климата, растительным и животным миром.
Тип урока: комбинированный..
Методы
– частично-поисковый;
– наглядно-иллюстративный;
– словесный;
– практический.
Форма работы – групповая, фронтальная, индивидуальная.
Оборудование: карта природных зон России, атласы Курской области, слайды и фотографии по теме урока, отрывки из художественных произведений, мультимедийный проектор, компьютер.
I. Оргмомент.
II.Проверка домашнего задания.
— Как называется раздел, над которым мы работаем?
— Что такое природная зона?
Природная зона — это крупный природный комплекс, обладающий общностью температурных условий и увлажнения почв, растительности и животного мира.
— Характеристику какой зоны мы дали на прошлом уроке?
I вариант: смешанный или широколиственный лес
II вариант: тайга
Географический
диктант.
Исп. экран
Задание: определить к какой зоне (тайга или смешанный, широколиственный лес) относятся следующие признаки:
З) Природная зона имеет более разнообразный растительный состав.
С) Эта природная зона занимает самую большую площадь.
О) Почвы дерново-подзолистые, серые лесные и бурые.
Т) Почвы подзолистые.
Е) В природной зоне деревья образуют один ярус.
Н) Эта зона отсутствует в Сибири.
П) Деревья более холодоустойчивы.
А) Условия зоны более благоприятны для земледелия.
Ь) В зоне много болот.
Самоконтроль:
I вариант: зона
II вариант: степь
III. Постановка цели и задач урока. Мотивация учебной деятельности учащихся.
Мотивация через постановку проблемного вопроса.
— Ребята, обратите внимание на ключ проверки
домашнего задания (степи).
— А сейчас, обратите внимание на изображение природных зон. фото.
— Какую особенность вы заметили?
Ответ. В изображении природных зон отсутствуют деревья.
— Скажите, а можно выйдя из лесной зоны сразу попасть в зону степей?
— Какая зона следует за лесной?
— Какие природные зоны нам предстоит сегодня исследовать?
— В какой из этих природных зон вы бывали?
— Какова будет цель нашего урока?
Задачи: Выявить главные причины, влияющие на размещение лесостепной и степной зоны по территории нашей страны.
Изучить особенности компонентов природы в лесостепях и степях в их взаимосвязях. На примере природных зон показать взаимоотношения природы и человека, выявить роль природы в жизни людей.
Тема урока « Природа лесостепей и степей »
записывается в тетрадь.
— Чтобы решить поставленные задачи, предлагаю совершить научную экспедицию по этим зонам.
— Перед вами маршрут нашего путешествия. На доске
(каждый ученик получает маршрутный лист)
Работа по карте
— Определить географическое положение лесостепей и степей.
1. Географическое положение
Лесостепь — это переходная зона от леса к степи. Лесостепная зона протянулась сплошной полосой от западной границы России до Алтая и Саян.
Зона степей невелика по площади, занимает юг европейской части России и Западной Сибири. В Восточной Сибири степи занимают межгорные котловины (Минусинскую, Тувинскую), т. к. при достаточно теплом лете, здесь недостаточно выпадает осадков (хребты закрывают доступ влажному воздуху, да и сами котловины находятся в центре материка, удалены от океанов).
III. Актуализация знаний.
Актуализация знаний.
Станция «Портретная»
1. Давайте «нарисуем»словесный портрет лесостепи и степи.
— Скажите, портрет какой зоны вы можете создать самостоятельно.
— А что такое лесостепь?
фото
…..– это переходная природная зона, которая сочетает в себе участки леса и участки степей. В лесостепи чередуются широколиственные и мелколиственные леса, а также разнотравные степи, поэтому в этой природной зоне есть животные и лесов и степей. Эта ландшафтная зона сильно изрезана оврагами и балками.
— Глядя на фотографию, ответьте на вопрос: Что же такое степь?
Степь — это безлесное пространство, умеренного пояса с господством травянистых растительных сообществ.
В русский язык слово ,, степь» вошло довольно поздно –
в начале 17 века. Но впервые слово ,,step» встречается в пьесе В. Шекспира ,,Сон в летнюю ночь»
(1600 года). Происхождение термина ,,степь» до сих пор не имеет
удовлетворительного объяснения. В разных языках мира это слово звучит
по-разному, но означает плоское, ровное, открытое место.
Но впервые слово ,,step» встречается в пьесе В. Шекспира ,,Сон в летнюю ночь»
(1600 года). Происхождение термина ,,степь» до сих пор не имеет
удовлетворительного объяснения. В разных языках мира это слово звучит
по-разному, но означает плоское, ровное, открытое место.
Степи — в России
Саванны – в Африке
Прерии – в Северной Америке
Пампа (пампасы) – в Южной Америке
2. Каждый ученик получает отрывок из повести А.П. Чехова “Степь”.
— Возьмите карандаши и подчеркните, по – вашему мнению, главные признаки степи.
Ровная, словно
скатерть, расстилается необъятная равнина, и там, где на ней сохранились
участки целины, она кажется в начале лета серебряной от цветущего ковыля и волнуется словно море. Ковыльная степь не изобилует множеством красок.
Яркой, пестрой от разнообразия красок, становится степь весной. Весна здесь
длится не долго — 2-3 недели. Это время, когда появляются растения-эфемеры:
тюльпаны, ирисы, и др. Пройдет немного времени и исчезнет яркий цветной ковер.
Останутся лишь подземные части растений — луковицы, корневища и будут
ждать в земле следующей весны, перенося летнюю засуху и зимние холода.
Пройдет немного времени и исчезнет яркий цветной ковер.
Останутся лишь подземные части растений — луковицы, корневища и будут
ждать в земле следующей весны, перенося летнюю засуху и зимние холода.
Человек чувствует себя затерянным среди этого необъятного и ровного пространства. Местами настолько ровного, что глаз видит его на много километров. С многочисленных же курганов, в беспорядке разбросанных по степи, кругозор увеличивается еще более… Даже у современного человека этот простор вызывает безотчетное желание побежать вдаль.
песня «Степь широкая…»
Вывод:
Главными признаками степей являются:
1) Травяной покров
2) Степи от лугов отличаются наличием в травяном покрове ковылей
3) Внешний облик степи сильно меняется в течение теплого периода, от весны до осени.
3. ЭТАП Создание проблемной ситуации
Станция «Климатическая»
— Какая растительность в лесной зоне? (древесная растительность)
— Какая растительность в степной зоне? (травянистая растительность)
— Как вы думаете, а какая растительность в лесостепи? (травянистая и
древесная)
— А почему? С чем это связано?
— Ваши предположения.
— Выдвигается гипотеза: изменился климат.
Карточка-задание для климатологов
1. Определить в каком климатическом поясе находятся лесостепь и степь.
2. Назвать тип климата.
3. Определить среднюю температуру января и июля.
4. Определить годовое количество осадков.
5. Определить характер увлажнения этой природной зоны.
Дополнительный материал. —
Опасные явления степей — это засуха, то есть длительный
период без осадков при сохранении высоких температур воздуха; суховеи — горячие сухие ветры, иссушающие почву; пыльные бури; их
причиной является распаханность степей) Суховей – сильный ветер (со скоростью
более 5 м/с) с высокой температурой (+25 — +30°С), продолжающийся иногда
несколько недель. Усиливает испарение, что вызывает нехватку влаги в почве.
—
Опасные явления степей — это засуха, то есть длительный
период без осадков при сохранении высоких температур воздуха; суховеи — горячие сухие ветры, иссушающие почву; пыльные бури; их
причиной является распаханность степей) Суховей – сильный ветер (со скоростью
более 5 м/с) с высокой температурой (+25 — +30°С), продолжающийся иногда
несколько недель. Усиливает испарение, что вызывает нехватку влаги в почве.
Зима в степной зоне короткая, с частыми оттепелями. В редкие годы понижается до – 15 С — 20 С . Снежный покров неустойчивый. Сильные ветры часто сдувают снег в овраги и балки.
Весна
в степях наступает рано. Она короткая и бурная. Часто в этот период сильные
ветры в отсутствии растительного покрова на полях, сдувает плодородный слой
почвы – так возникают пыльные бури. Так, например, в январе 1969 года сильная
пыльная буря занесла посёлок Заречный, располагавшийся на левом берегу нашей
реки Ясени.
В некоторых районах Ставрополья после пыльных бурь у лесных полос накапливались валы выдутого мелкозёма высотой до 4 м.
Вслед за весной наступает жаркое, сухое лето. Средняя температура Июля + 22, + 24 С. Осадки выпадают редко в виде коротких, но сильных ливней. При длительном сухом периоде, при высоких температурах воздуха и сильном ветре возникают суховее – горячие сухие ветры, которые губительно влияют на растения степей.
Станция «Почвенная»
Карточка — задание для почвоведов
1. Почему В.В. Докучаев назвал почвы » зеркалом ландшафта»?
2. Какие почвы преобладают в степях?
3. Почему почвы степей очень плодородны.
4. Какие проблемы существуют с использованием чернозёмов.
— Почему почвы степей обладают высоким плодородием?
(Много гумуса, нет промыва почв, лето длинное и теплое, благоприятный ежегодный травяной опад — благоприятные условия для почвообразования)
На
севере зоны это типичные черноземы, у которых мощность гумусового горизонта
достигает от 60 до 100 см. В.В.Докучаев – русский почвовед, назвал эти почвы –
Царем почв, потому что они обладают высоким плодородием – содержат много
гумуса, который улучшает структуру почвы. На юге – южные черноземы с укороченным
до 25 см гумусовым горизонтом. В самых засушливых частях зоны – каштановые
почвы. Встречаются засоленные почвы.
В.В.Докучаев – русский почвовед, назвал эти почвы –
Царем почв, потому что они обладают высоким плодородием – содержат много
гумуса, который улучшает структуру почвы. На юге – южные черноземы с укороченным
до 25 см гумусовым горизонтом. В самых засушливых частях зоны – каштановые
почвы. Встречаются засоленные почвы.
Тип почвы зависит от климата, растительного покрова, материнской породы. Все факторы, на большей части зоны способствуют образованию большого горизонта с мощным гумусовым слоем.
Станция «Ботаническая»
Карточка- задание для ботаников
1. Выявить какая растительность преобладает в степях (Демонстрация гербария)
2. Почему в степях не растут деревья.
3. Как климатические условия отразились на растительности.
4. Почему степи зеленеют лишь ранней весной.
5. Каким образом хозяйственная деятельность человека повлияла на растительный мир.
Дополнительный
материал. Эфемеры-растения с коротким сроком вегетации (тюльпаны, пионы, ирисы,
сон-трава)
Эфемеры-растения с коротким сроком вегетации (тюльпаны, пионы, ирисы,
сон-трава)
— Какие растения можно увидеть в степи весной?
(Эфемеры — тюльпаны, пионы, ирисы, сон-траву)
— Что такое эфемеры? (Растения с коротким сроком вегетации)
— Почему у растений степи длинные и разветвленные корни, маленькие опущенные листья? (Растения добывают влагу с большой глубины, а опущенные листья испаряют меньше влаги)
— Где в степи можно увидеть деревья? (В долинах рек, где ближе расположены грунтовые воды)
Типичные представители растительного мира степей – различные виды ковылей, типчак, пырей, овсюг, виды , перекати-поле», спорыш, подорожник, шалфей, тюльпан.
Вывод: В степях, в основном, растут травы. Древесная
растительность на большей части зоны отсутствует и встречается лишь по берегам
рек или в балках. Эти леса называются урёмами. Деревьям в степи не хватает
влаги. А вот травы выработали универсальные приспособления к жизни в сухих
местах.
Ученики называют особенности степных трав – тонкие листья, стебли, мощная корневая система, луковичные виды растений.
Станция «Зоологическая»
Просмотр кинофильма Животные степи Выписать в тетрадь
Задание: Просмотрев фильм, выписать животных степей России.
Карточка — задание для зоологов
1.Какие животные обитают в степи?
2. Как животные степей приспособились к проживанию в этой природной зоне?
3. Почему многие животные степей живут стадами, а не по одному как в лесу?
4. Среда обитания степных животных очень сильно изменена человеком. Почему?
Растительный и животный мир лесостепной и степной зон.
Благодаря обилию пищи в степях много животных. Среди
млекопитающих – копытные (сайгаки), грызуны (хомяки, суслики, полевки), а также
зайцы русаки, волки, рыжие лисицы. Насекомые – главная пища
многих птиц. Большим разнообразием отличаются птицы: дрофа, стрепет, серая
куропатка, жаворонок. Хищные: степной орел ,лунь, сова. Жизнь
многих животных связанна с травами. Это насекомые: мухи, бабочки,
шмели, и пчёлы. В лесостепной зоне расселяются как лесные, так и степные
животные. Животные степей — обитатели открытых пространств.
Отсутствие естественных укрытий заставляет их приспосабливаться: — жизнь
большими группами, быстрый бег, маскирующая окраска (под цвет почвы),
способность к рытью нор и жизни под землей
Насекомые – главная пища
многих птиц. Большим разнообразием отличаются птицы: дрофа, стрепет, серая
куропатка, жаворонок. Хищные: степной орел ,лунь, сова. Жизнь
многих животных связанна с травами. Это насекомые: мухи, бабочки,
шмели, и пчёлы. В лесостепной зоне расселяются как лесные, так и степные
животные. Животные степей — обитатели открытых пространств.
Отсутствие естественных укрытий заставляет их приспосабливаться: — жизнь
большими группами, быстрый бег, маскирующая окраска (под цвет почвы),
способность к рытью нор и жизни под землей
— Назовите каких животных можно увидеть в нашей местности? (лиса, волк, заяц, хорек, уж, полоз, крот, суслик. Несекомые- оса, шмель, стрекоза, комары. Птицы — куропатка, сова, сорока, перепел)
Взаимосвязь компонентов ПК степь
Природные компоненты степи.
Станция «Экологическая»
Карточка — задание для экологов
1. Чем занимаются люди, живущие в степях?
Чем занимаются люди, живущие в степях?
2. Какие сельскохозяйственные культуры выращивают жители степей?
3. Как хозяйственная деятельность людей отразилась на природе степей?
4. Что необходимо помнить, чтобы сохранить природу степей?
5. Какие в степях созданы заповедники?
Исп. слайды
Сравнение степи сегодня с тем, что было
Послушайте, как описывает степь Н. В. Гоголь в своём произведении ,,Тарас Бульба».
Степь
чем далее, тем становилась прекраснее. Тогда весь юг, все то пространство,
которое составляет нынешнюю Новороссию, до самого Черного моря, было зеленою,
девственною пустынею. Никогда плуг не проходил по неизмеримым волнам диких
растений. Одни только кони, скрывавшиеся в них, как в лесу, вытаптывали их.
Ничего в природе не могло быть лучше. Вся поверхность земли представлялася
зелено-золотым океаном, по которому брызнули миллионы разных цветов. Сквозь
тонкие, высокие стебли травы сквозили голубые, синие и лиловые волошки; желтый
дров выскакивал вверх своею пирамидальною верхушкою; белая кашка
зонтикообразными шапками пестрела на поверхности; занесенный бог знает откуда
колос пшеницы наливался в гуще. Под тонкими их корнями шныряли куропатки,
вытянув свои шеи. Воздух был наполнен тысячью разных птичьих свистов. В небе неподвижно
стояли ястребы, распластав свои крылья и неподвижно устремив глаза свои в
траву. Крик двигавшейся в стороне тучи диких гусей отдавался бог весть в каком
дальнем озере. Из травы подымалась мерными взмахами чайка и роскошно купалась в
синих волнах воздуха. Вон она пропала в вышине и только мелькает одною черною
точкою. Вон она перевернулась крылами и блеснула перед солнцем… Черт вас
возьми, степи, как вы хороши!..
Под тонкими их корнями шныряли куропатки,
вытянув свои шеи. Воздух был наполнен тысячью разных птичьих свистов. В небе неподвижно
стояли ястребы, распластав свои крылья и неподвижно устремив глаза свои в
траву. Крик двигавшейся в стороне тучи диких гусей отдавался бог весть в каком
дальнем озере. Из травы подымалась мерными взмахами чайка и роскошно купалась в
синих волнах воздуха. Вон она пропала в вышине и только мелькает одною черною
точкою. Вон она перевернулась крылами и блеснула перед солнцем… Черт вас
возьми, степи, как вы хороши!..
Возможность использования ресурсов природной зоны человеком
Хозяйственное использование:
В настоящее время лесостепная и
степная зона практически полностью
распаханы. Степи более чем на 80% распаханы. Степь — главная
зерновая житница страны. Здесь
выращивают пшеницу,
кукурузу, подсолнечник, сахарную свеклу, просо. Развито плодоводство и бахчеводство, а в поймах — овощеводство. Сохранившиеся
участки степной растительности используются в качестве пастбищ.
Развито плодоводство и бахчеводство, а в поймах — овощеводство. Сохранившиеся
участки степной растительности используются в качестве пастбищ.
Последствия использования степи в хозяйстве – полное изменение природной зоны. Многие виды флоры и фауны занесены в Красную книгу.
Сейчас настоящий первозданных степей почти не осталось. Дикая растительность и животные почти уничтожены и встречаются лишь в плавнях, балках, обочинах дорог и в заповедниках.
Различно используются в природной зоне гидроресурсы. Загрязнение водоемов, их обмельчание. В Красную книгу занесены: русский осетр, стерлядь, белуга, севрюга.
Пути решения экологических проблем зоны
– система мер по охране природы: мероприятия по сохранению редких и
исчезающих видов и растений и животных. Необходимо создавать заповедники,
заказники, восстанавливать пойменные леса, защитные полосы; прекратить или
уменьшить добычу полезных ископаемых. Строить экологически чистые предприятия,
на уже существующие ставить биофильтры, воздухоочистители и водоочистители.
Контакт со степью наложил отпечаток на формирование духовного облика народа. Чувство доли и шири, коллективизм, стремительность, вольность, лёгкость на подъём – вот черты характера жителя степной зоны, сформированные природой степи.
Историческим продуктом степи был казак, приёмник древних киевских богатырей. На широких степных просторах нашли себе применение отвага и удаль казаков, их мужество и жажда бранных подвигов.
5. Первичное усвоение новых знаний
1. Сформулируй несколько вопросов по изученной теме.
(Помни, что вопросы делятся по категориям: ЧТО?; ГДЕ? или КАК?; ПОЧЕМУ?)
2. Итак,
«Географическая лихорадка».
Я называю представителя лесостепной зоны, а вы
подтверждаете или отрицаете его наличие в данной природной зоне.
Игра “Географическая лихорадка”
а) названия растений лесостепей:
— ковыль, пырей, тимофеевка, клевер, мышиный
горошек, береза, осина, дуб, липа, ива, боярышник, шиповник, смородина,
костяника, клубника, сосна, рогоз, камыш.
б) птицы лесостепей:
серые куропатки, грачи, сороки, дятлы, кукушки, тетерева,
перепела, жаворонки, гуси, утки, журавли.
в) животные и пресмыкающиеся:
суслики, серые полевки, тушканчики, зайцы,
ласки, лисицы, волки, белки, лоси, ондатры, ящерицы, ужи, гадюки.
3. Составление причинно– следственной цепочки.
В процессе составления причинно– следственной цепочки подвести итог
— Составить причинно– следственную цепочку из следующих утверждений:
А) Растительность — однолетние травы;
Б) Находится на юге России;
В) Главная житница страны ;
Г) Животные – грызуны, насекомые, копытные, птицы ;
Д) Почвы – черноземы;
Е) Зима холодная, лето жаркое, увлажнение недостаточное.
Обсуждение. Проверка. (БЕДАГВ)
Лесостепи и степи – наиболее измененные
человеком природные зоны. Практически все природные ландшафты преобразованы, а
участки относительно нетронутой природы можно встретить только в заповедниках.
Степи и лесостепи – житница страны. Благоприятный климат, плодородные почвы
стали причиной активного сельскохозяйственного освоения этих земель. Каждый
компонент не существует отдельно, независимо друг от друга или параллельно.
Четко прослеживается связь между всеми компонентами природы. Климат влияет на
растения и животный мир, растения на образование почвенного покрова и т. д.
Отметили, что изменение одного компонента природы влечет за собой изменение и
другого.
Благоприятный климат, плодородные почвы
стали причиной активного сельскохозяйственного освоения этих земель. Каждый
компонент не существует отдельно, независимо друг от друга или параллельно.
Четко прослеживается связь между всеми компонентами природы. Климат влияет на
растения и животный мир, растения на образование почвенного покрова и т. д.
Отметили, что изменение одного компонента природы влечет за собой изменение и
другого.
Информация о домашнее задание.
Для всех обучающихся: § 42
Индивидуально:
— Найти дополнительный материал о заповедниках зоны степей.
— Подготовить материал о растениях и животных Курской области, занесённых в Красную книгу.
— Подготовить презентацию «Природная зона моей Малой родины».
Рефлексия.
— Для чего мы изучили данную тему ? (Чтобы грамотно использовать в хозяйственных целях, Чтобы было как можно меньше экологических проблем.)
Попробуйте оценить собственные знания и умения следующим образом:
1. Я всё понял. Смогу
объяснить другому.
Я всё понял. Смогу
объяснить другому.
2.Я понял материал, смогу объяснить другому, но при некоторой помощи учителя.
3.Я понял материал частично.
4.Я ничего не понял.
Новые понятия:
Колки— березовые рощи
Уремы— пойменные леса
Эфемероиды— многолетние травянистые растения.
Ксерофиты— засухоустойчивые растения.
Эфемеры— однолетние, мелкие травянистые растения
Степные блюдца—
многочисленные неглубокие западины, заросшие невысокими осиновыми деревцами или
кустами.
Точно нет им края.
И стоит вверху, над степью,
Тишина немая.
Нестерпимою жарою
Воздух так и пышет
Как шумит трава густая,
Только ухо слышит.
Едешь, едешь — как шальные
Кони мчатся степью,
Вдаль курганы, зеленея,
Убегаю цепью.
Промелькнут перед глазами
Две три старых ивы-
И опять перед глазами,
Ветра переливы.
Едешь, едешь — степь да небо…
Степь, вся степь, как море…
И взгрустнётся поневоле
На таком просторе.
И. З. Суриков.
Природные зоны России — лесостепи и степи | Презентация к уроку по географии (8 класс) по теме:
Степи и лесостепи
Цели и задачи урока: познакомить с характеристикой зоны степей и лесостепей. Формировать представления о хозяйственном использовании ресурсов природной зоны и ее проблемах
Оборудование: карта «Природные зоны России», изображения животных и растений зоны степей и лесостепей
Ход урока
I. Организационный момент
Организационный момент
II. Проверка домашнего задания
Расставить цифры к природным зонам: тайга, смешанные и широколиственные леса
- Занимает наибольшую площадь
- Более разнообразный растительный состав
- Средняя температура января достигает до -40 °С
- Распространены серые лесные почвы
- Деревья образуют один ярус
- Много певчих птиц
- Деревья более холодоустойчивы
- Древесина — главное богатство этой зоны
- Сильная заболачиваемость
- Более благоприятна для земледелия
- Произрастают только хвойные деревья
- Численность животных сильно сокращена
- Климат мягкий, лето продолжительное
- Зона отсутствует в Сибири
Тайга — 1, 3, 5, 7, 8, 9, 11
Смешанный и широколиственный лес — 2, 4, 6, 10, 12, 13, 14
III. Изучение нового материала
Природная зона лесостепей и степей
1. Географическое положение
Лесостепь — это переходная зона от леса к степи. Лесостепная зона протянулась сплошной полосой от западной границы России до Алтая и Саян.
Лесостепная зона протянулась сплошной полосой от западной границы России до Алтая и Саян.
Зона степей невелика по площади, занимает юг европейской части России и Западной Сибири. В Восточной Сибири степи занимают межгорные котловины (Минусинскую, Тувинскую), т. к. при достаточно теплом лете, здесь не достаточно выпадает осадков (хребты закрывают доступ влажному воздуху, да и сами котловины находятся в центре материка, удалены от океанов)
2. Типичный внешний облик зоны
Портрет степи
Беспредельная степь! Ровная, словно скатерть, расстилается необъятная равнина, и там, где на ней сохранились участки степной целины, она кажется в начале лета серебряной от цветущего ковыля и волнуется словно море. Ковыльная степь не изобилует множеством красок. Яркой, пестрой от разнообразия красок, становится степь весной. Весна в степи длится не долго — 2-3 недели. Это время, когда появляются в степи растения-эфемеры: тюльпаны, ирисы, и др. Пройдет немного времени и исчезнет яркий цветной ковер. Останутся лишь подземные части растений — луковицы, корневища и будут ждать в земле следующей весны, перенося летнюю засуху и зимние холода.
Останутся лишь подземные части растений — луковицы, корневища и будут ждать в земле следующей весны, перенося летнюю засуху и зимние холода.
Человек чувствует себя затерянным среди этого необъятного и ровного пространства. Местами настолько ровного, что глаз видит его на много километров. С многочисленных же курганов, в беспорядке разбросанных по степи, кругозор увеличивается еще более… Даже у современного человека степной простор вызывает безотчетное желание побежать вдаль.
Главными признаками степей являются:
1) Травяной покров
2) Степи от лугов отличаются наличием в травяном покрове ковылей
3) Внешний облик степи сильно меняется в течение теплого периода, от весны до осени
3. Климатические условия
Умеренный климатический пояс
Климат лесостепи теплее и суше, чем климат лесных зон. Средняя температура июля составляет +19 — 22°С. Годовая сумма осадков 400—500 мм. Коэффициент увлажнения на севере зоны близок к единице, а к югу постепенно уменьшается. Соотношение тепла и влаги в лесостепи близко к оптимальному, но увлажнение неустойчивое. Бывают засухи и суховеи. В результате увеличения испарения снижается речной сток и водность рек. Глубже залегают грунтовые воды. Возрастает их минерализация.
Соотношение тепла и влаги в лесостепи близко к оптимальному, но увлажнение неустойчивое. Бывают засухи и суховеи. В результате увеличения испарения снижается речной сток и водность рек. Глубже залегают грунтовые воды. Возрастает их минерализация.
В степи зима короткая, но холодная. Суровость ее нарастает к востоку. Средняя температура января — – 10°С. Мощность снежного покрова мала. После короткой и бурной весны наступает жаркое, сухое лето. Средняя температура июля составляет +21…+23°С. Здесь отчетливо прослеживаются черты засушливости климата. Дожди выпадают редко, обычно в виде коротких ливней, которые слабо увлажняют почву. Осадков выпадает мало (350—450 мм), примерно столько же, сколько в зоне тундры. Но тундра заболочена и отличается избыточным увлажнением. В степях же наблюдается недостаток влаги. Испаряемость значительно превышает количество осадков. Коэффициент увлажнения изменяется от 0,6 у северной границы зоны до 0,3 на юге. Сильные ветры обусловливают значительное испарение с поверхности и периодически происходящие засухи, суховеи, пыльные бури.
Реки маловодны. Многие из них летом пересыхают. Грунтовые воды лежат глубоко и не всегда пригодны для питья, т.к. засолены.
4. Почвы и их свойства
В лесостепной зоне на междуречьях чередуются широколиственные (дубовые) и мелколиственные леса на серых лесных почвах с разнотравными степями на черноземах.
Так как осадков выпадает немного, а испаряемость в 2 раза превышает количество осадков, в степи нет условий для вымывания перегноя в глубину почвенных горизонтов. В степи распространены черноземы с очень темной окраской и с зернистой структурой. Мощность перегнойного горизонта достигает 50—80 см. В бассейне реки Кубань граница этого горизонта отмечена на глубине 1,5 м. Черноземы — самые плодородные почвы нашей страны, их русский почвовед В. В. Докучаев называл царем почв. В южной полосе степей распространены темно-каштановые почвы, менее плодородные и нередко засоленные.
5. Растения и их приспособленность к условиям зоны
В прошлом в степях господствовала разнотравно-злаковая растительность. Ранней весной на еще влажной от стаявших снегов земле в степи расцветали многочисленные яркие цветы – эфемеры (запас питательных веществ в луковицах и корневищах). Проходило несколько недель, и степь становилась иной — в это время пышно развивались злаки: ковыли, типчак, тонконог. Ныне лишь в заповедниках можно увидеть, как красивы ковыльные степи.
Ранней весной на еще влажной от стаявших снегов земле в степи расцветали многочисленные яркие цветы – эфемеры (запас питательных веществ в луковицах и корневищах). Проходило несколько недель, и степь становилась иной — в это время пышно развивались злаки: ковыли, типчак, тонконог. Ныне лишь в заповедниках можно увидеть, как красивы ковыльные степи.
Из-за длительных засух травы уже к середине лета жухнут и выгорают. Вот почему степные растения имеют глубокие, разветвленные корни. Листья у них маленькие и опущенные — приспособление для меньшей потери влаги через испарение: на таких листьях меньше устьиц.
6. Животные и их приспособленность к условиям зоны
В лесостепях обитают лесные виды зверей и птиц, но разнообразие их и количество невелико. В животном мире степей преобладают различные мелкие грызуны — суслики, сурки, тушканчики, хомяки, полевки. Они делают запасы на зиму. Грызуны роют норы, птицы вьют гнезда. Из-за недостатка укрытий другие животные быстро бегают и ведут стадный образ жизни. По доисторическим степям бродили стада диких лошадей, сайгаки, ныне оттесненные в полупустыни, быки-туры, полностью уничтоженные в Восточной Европе.
По доисторическим степям бродили стада диких лошадей, сайгаки, ныне оттесненные в полупустыни, быки-туры, полностью уничтоженные в Восточной Европе.
ФИЗКУЛЬТМИНУТКА
Практическая работа «Сравнение безлесных зон России»
Параметры сравнения | Тундра | Степь |
ГП | побережье Северного Ледовитого океана | юг Русской и Западно-Сибирской равнины |
Причины безлесия | суровый климат, избыток влаги | недостаток влаги |
Жизненные формы растений | подушкообразные, стелющиеся формы, небольшой рост | травы |
Растения | мхи, лишайники, ягодные кустарнички и карликовые деревья | ковыль, злаки, тюльпаны |
Вывод: и тундра, и степь — безлесные зоны. Но вызвано это разными причинами: разным соотношением тепла и влаги
7. Возможность использования ресурсов природной зоны человеком
Возможность использования ресурсов природной зоны человеком
В настоящее время лесостепная и степная зона практически полностью распаханы. Степь — главная зерновая житница страны. Здесь выращивают пшеницу, кукурузу, подсолнечник, сахарную свеклу, просо. Развито плодоводство и бахчеводство, а в поймах — овощеводство. Сохранившиеся участки степной растительности используются в качестве пастбищ.
8. Проблемы охраны природной зоны
Распаханность территории и ливневый характер осадков способствуют смыву верхнего плодородного горизонта почв и образованию оврагов на полях. В степях развита и ветровая эрозия. Суховеи иногда переходят в пыльные бури. Для защиты полей от губительного действия суховеев и задержания снега в степной зоне создаются лесные полосы. Первые опытные лесные полезащитные полосы были созданы в Каменной степи на юге Среднерусской возвышенности учениками В. В. Докучаева.
IV. Закрепление
Почему отсутствуют смешанные и широколиственные леса в Западной Сибири? (Здесь более холодные зимы, низкие температуры зимой не позволяют расти более теплолюбивым породам, в степи дефицит влаги, деревья более влаголюбивые растения, чем травы)
Почему почвы степей обладают высоким плодородием? (Много гумуса, нет промыва почв, лето длинное и теплое, благоприятный ежегодный травяной опад — благоприятные условия для почвообразования)
Какие растения можно увидеть в степи весной? (Эфемеры — тюльпаны, пионы, ирисы, сон-траву)
Что такое эфемеры? (Растения с коротким сроком вегетации)
Почему у растений степи длинные и разветвленные корни, маленькие опущенные листья? (Растения добывают влагу с большой глубины, а опущенные листья испаряют меньше влаги)
Где в степи можно увидеть деревья? (В долинах рек, где ближе расположены грунтовые воды)
Каков современный ландшафт степей? (Степи более чем на 80% распаханы)
Какие опасные явления могут быть в степях? Что является их причиной? (Опасные явления степей — это засуха, то есть длительный период без осадков при сохранении высоких температур воздуха; суховеи — горячие сухие ветры, иссушающие почву; пыльные бури; их причиной является распаханность степей)
На к/карту: закрасить светло-желтым и желтым цветом зону степей
V. Домашнее задание: подготовить рассказ о природной зоне степей и лесостепей, найти и выписать значение слов: урёма, суховей, ксерофиты, сделать рисунок любого представителя растительного или животного мира этой природной зоны (по желанию)
Домашнее задание: подготовить рассказ о природной зоне степей и лесостепей, найти и выписать значение слов: урёма, суховей, ксерофиты, сделать рисунок любого представителя растительного или животного мира этой природной зоны (по желанию)
Урёма – лиственные леса, расположенные в поймах рек в степной зоне
Суховей – сильный ветер (со скоростью более 5 м/с) с высокой температурой (+25 — +30°С), продолжающийся иногда несколько недель. Усиливает испарение, что вызывает нехватку влаги в почве
Ксерофиты – засухоустойчивые растения, переносящие длительный недостаток влаги в почве
Восточно-Европейская Лесостепь | One Earth
Площадь территории экорегиона указана в единицах по 1000 га. Целью защиты является зона Глобальной сети безопасности (GSN1) для данного экорегиона. Уровень защиты указывает процент цели GSN, которая в настоящее время защищена, по шкале от 0 до 10. N/A означает, что в настоящее время данные недоступны.
Биорегион: Европейские внутренние смешанные леса (PA12)
Царство: Западная Евразия
Ecoregion Size (1000 ha):
72,884
Ecoregion ID:
661
Protection Goal:
20%
Protection Level:
3
Государства: Россия, Украина, Румыния, Молдова, Болгария
Пологие холмистые низменности Восточно-Европейской Лесостепи представляют собой сочетание зеленых лесов и великолепных лугов. Глубокие овраги и овраги прорезают тропы через склоны холмов, украшенные множеством ярких цветов. Лоси и олени находят убежище в лесах, а открытые пастбища — рай для разнообразных грызунов. Суслики и хомяки являются излюбленной добычей величественных восточных орлов-могильников; когда-то почитаемые как священные птицы, эти недавно преследуемые хищники теперь сохраняют свою европейскую цитадель в лесостепи Российской Федерации.
Глубокие овраги и овраги прорезают тропы через склоны холмов, украшенные множеством ярких цветов. Лоси и олени находят убежище в лесах, а открытые пастбища — рай для разнообразных грызунов. Суслики и хомяки являются излюбленной добычей величественных восточных орлов-могильников; когда-то почитаемые как священные птицы, эти недавно преследуемые хищники теперь сохраняют свою европейскую цитадель в лесостепи Российской Федерации.
Флагманский вид экорегиона Восточно-Европейская Лесостепь — восточный орел-могильник. Изображение предоставлено: Creative Commons
Большая часть этого экорегиона находится в пределах Российской Федерации, простираясь на Украину, Молдову, предгорья Карпат в Румынии и, наконец, на северо-восток Болгарии. Эта мозаика лесов и степей представляет собой переходную зону, соединяющую широколиственные леса умеренного пояса на севере со степными лугами на юге. Это граница между влажной и засушливой средой, где наблюдается умеренно-континентальный климат с теплым летом и холодной зимой.
Леса в основном состоят из дуба обыкновенного, граба восточного, сосны обыкновенной, клена остролистного и ясеня европейского, хотя также распространены осина и береза повислая. На лугах много красочных видов: степной сорняк, узколистный мятлик, ковыль украшают лютик многоцветковый, шалфей луговой, лабазник.
Этот переходный ландшафт порождает смешение лесных и степных видов. Лесной лось, косуля, ястреб и дятел сосуществуют с типичной степной фауной: обыкновенным хомяком, европейским сусликом и почти эндемичным большим слепышем. Популяции некоторых грызунов и птиц увеличиваются вокруг пахотных земель, поскольку они приспособились использовать зерно в качестве дополнительного источника пищи; Восточный орел-могильник — один из многих видов, которые в настоящее время населяют почти исключительно сельскохозяйственные ландшафты.
Первоначально леса покрывали до одной трети территории лесостепи, но вырубка лесов значительно сократила эту долю до примерно 10%. Пахотные угодья в настоящее время составляют 42% ландшафта, в основном для выращивания озимой пшеницы и кукурузы, хотя также широко выращиваются сахарная свекла и табак. Большинство оставшихся территорий были преобразованы под инфраструктуру, промышленность или поселения; Лесостепь имеет самую высокую плотность населения в Российской Федерации.
Большинство оставшихся территорий были преобразованы под инфраструктуру, промышленность или поселения; Лесостепь имеет самую высокую плотность населения в Российской Федерации.
Остались только небольшие фрагменты нетронутой среды обитания, либо там, где топография препятствует развитию, либо там, где заповедники обеспечивают защиту. Однако такие охраняемые территории занимают очень небольшую часть суши. В природном заповеднике Калужские засеки в Российской Федерации находятся девственные дубовые леса, в которых обитает большое количество диких и хищных птиц, а особый заповедник Хорга-Зорлени в Румынии имеет жизненно важное значение для множества видов птиц.
Косуля. Изображение предоставлено: Ян Бо Кристенсен, Creative Commons
Широко распространенная эрозия представляет собой серьезную угрозу, вызванную обширной вырубкой лесов и распашкой. В результате увеличение стока воды приводит к потере верхнего слоя почвы; В лесостепи Поволжья за последние 100 лет утрачено 69% органического вещества. Лесостепные системы также особенно чувствительны к инвазии неместных видов, таких как молочай обыкновенный и маслина русская; последний был завезен в регион в рамках научно необоснованного степного лесоразведения.
Лесостепные системы также особенно чувствительны к инвазии неместных видов, таких как молочай обыкновенный и маслина русская; последний был завезен в регион в рамках научно необоснованного степного лесоразведения.
В последнее время расширение добычи и бурения нефтяных и газовых скважин в Российской Федерации представляет угрозу для природных экосистем. Усилия по сохранению включали акцент на увеличение представленности охраняемых территорий в Украине, в то время как в Российской Федерации пропагандируются более экологически безопасные методы ведения сельского хозяйства; это наряду с мерами по сокращению обезлесения за счет реализации защитного законодательства и регулярной инвентаризации.
Приоритетными природоохранными действиями на следующее десятилетие будут: 1) обеспечение соблюдения законодательства, направленного на предотвращение дальнейшего обезлесения или преобразования лесостепной среды обитания; 2) расширить охват охраняемых территорий, уделяя особое внимание непрерывному охвату за пределами страны; и 3) инвестировать в научные исследования эффективности методов управления сохранением, чтобы избежать неуместных усилий.
Цитаты
1. Чибилёв А., 2002. 1 1 Степь и Лесостепь. Физическая география северной Евразии , 3 , стр.248
Дулай Г., Лю Х., Магнес М., Молнар З. и Накинежад А., 2018 г. iГрань двух миров: новый обзор и синтез по евразийским лесостепям. Прикладная наука о растительности , 21 (3),
3. Стратегия и исполнительный план сохранения биоразнообразия в Российской Федерации. (2014). Москва, Российская Федерация
Степь (луга) · Университет Пьюджет-Саунд
Назад СледующийКлимат
Луга (степи) представляют собой среду с умеренным климатом, с теплым или жарким летом и прохладной или очень холодной зимой; температуры часто бывают экстремальными в этих среднеконтинентальных районах. Они часто располагаются между умеренными лесами и пустынями, а годовое количество осадков выпадает между характерными для этих зон количествами. Ветры играют важную роль в этих очень открытых средах. Осадки варьируются от очень сезонных до круглогодичных. В северной части зоны наблюдается значительное накопление снега, которое значительно уменьшается к югу.
Осадки варьируются от очень сезонных до круглогодичных. В северной части зоны наблюдается значительное накопление снега, которое значительно уменьшается к югу.
Почвы
Луга обычно подстилаются черноземами, почвами, которые являются щелочными, поскольку чистое движение воды в них направлено вверх, неся с собой кальций, который осаждается в виде карбоната кальция. Степные черноземы в верхних горизонтах черноватые из-за постоянного перегнивания злаков в темный гумус. Высокотравные прерии имеют более коричневые почвы, более богатые гумусом и суглинистые по структуре.
Растительность
В пастбищах в основном преобладают травы, но однолетние и многолетние травы смешаны в разных пропорциях на разных участках. Средняя высота травы коррелирует с количеством осадков, поэтому в Северной Америке есть зоны прерий с высокой, средней и низкой травой по продольному градиенту с востока на запад. Внешний вид значительно меняется между весной, когда травы зеленеют и цветут разнотравье, и серединой лета, когда преобладают коричневые, семенные и отмирающие растения. В некоторых степях, особенно в более засушливых районах, преобладают кустарники (их обозначают как кустарниково-степные) и они отличаются от пустынь прежде всего более высокими широтами, более низкими средними температурами и меньшим разнообразием.
В некоторых степях, особенно в более засушливых районах, преобладают кустарники (их обозначают как кустарниково-степные) и они отличаются от пустынь прежде всего более высокими широтами, более низкими средними температурами и меньшим разнообразием.
Разнообразие
Разнообразие растений и животных довольно низкое в этой структурно простой зоне с умеренным климатом. Например, обычно на типичных пастбищах умеренного пояса встречается не более двух-трех видов крупных пастбищных млекопитающих, по сравнению с дюжиной или более на некоторых тропических (саванных) пастбищах. Птицы разнообразны только в заболоченных местах и в прибрежной растительности вдоль рек. Луга южного полушария, прилегающие к тропическим лесам и саваннам, могут иметь более высокое разнообразие животных, чем в северном полушарии. Единственными крупными группами позвоночных, особенно характерными для пастбищ, являются воробьиные птицы — жаворонки, коньки и овсянки. Семейство злаков является одним из крупнейших в мире, а травы как отдельные растения, вероятно, являются самыми многочисленными растениями в мире.
Адаптация растений
Травы великолепно приспособлены для покрытия открытого грунта, относительно устойчивы как к огню, так и к выпасу, поскольку их листья растут от основания, в отличие от большинства растений, у которых новые листья постоянно растут с кончиков ветвей. Таким образом, поскольку продуктивная часть травянистого растения сгорает или выедается, пока его основание остается неповрежденным, немедленно следует новый рост. Листья трав также богаты кремнеземом, который изнашивает зубы пасущихся животных, а некоторые разнотравье этой зоны (например, локови) очень токсичны для травоядных. Большинство трав размножаются с помощью корневищ, стеблей, которые проходят прямо под землей и через равные промежутки времени выпускают новые листья. Другие, особенно в более засушливых районах, растут в виде пучков трав, растений, которые сопротивляются высыханию ветром благодаря густоте своей формы роста; многие разнотравья этой зоны имеют аналогичное строение. Травы опыляются ветром, что очень эффективно в этой открытой среде.
Адаптации для животных
Копание нор является важной адаптацией для мелких животных, позволяющей избежать нападения хищников в этой открытой среде. Некоторые из нор являются колониальными, вероятно, также адаптацией для раннего обнаружения хищников. Некоторые хищники среднего размера хорошо приспособлены для выкапывания мелких нор. Из-за своей открытости и, следовательно, легкости передвижения эта среда поддерживает большие популяции пасущихся млекопитающих, которые образуют стада для защиты от крупных хищников, следующих за ними. Гнездящиеся и кормящиеся на земле птицы очень подвержены хищничеству, и почти все они загадочны, с коричневыми прожилками наверху, с хорошо развитыми отвлекающими проявлениями; без приподнятых насестов большинство поет с воздуха.
Воздействие на человека
Поскольку все зерновые культуры являются травами, эта среда хорошо подходит для них; таким образом, огромные области лугов умеренного пояса были изменены для выращивания зерна для питания людей и скота. Выпас скота оказывает более интенсивное воздействие на многие районы, чем местные копытные, изменяя представленность видов в растительных сообществах, поскольку одни из них более восприимчивы к выпасу, чем другие. Многие растения этой среды адаптированы к ранней сукцессии и при интродукции на другие континенты становятся инвазивными сорняками; они быстро колонизируют нарушенные участки и часто вытесняют местные виды. Вместе с выпасом скота это повсеместно изменило состав пастбищ, особенно в Северной Америке, так что «естественные» пастбища стали редкостью.
Выпас скота оказывает более интенсивное воздействие на многие районы, чем местные копытные, изменяя представленность видов в растительных сообществах, поскольку одни из них более восприимчивы к выпасу, чем другие. Многие растения этой среды адаптированы к ранней сукцессии и при интродукции на другие континенты становятся инвазивными сорняками; они быстро колонизируют нарушенные участки и часто вытесняют местные виды. Вместе с выпасом скота это повсеместно изменило состав пастбищ, особенно в Северной Америке, так что «естественные» пастбища стали редкостью.
Леса | Бесплатный полнотекстовый | Климатически обусловленная голоценовая миграция лесостепного экотона гор Тянь
1. Введение
Засушливые и полузасушливые районы чрезвычайно чувствительны к изменению климата и деятельности человека, и лесостепной экотон в этих районах является одним из наиболее уязвимых экосистем в мире [1,2,3]. До настоящего времени изменение климата представляло серьезную угрозу экотону лесостепи в засушливых и полузасушливых районах, вызывая снижение роста и гибель деревьев [2,4,5]. Последние прогнозы предполагают, что расширение засушливых и полузасушливых районов потенциально усугубит региональное потепление и фрагментацию среды обитания в будущем [3,6]. Поэтому возникает острая необходимость обратить внимание на экотон лесостепи в таких экологически уязвимых районах.
Таксоны пыльцы концентрируются в горах засушливых регионов, которые также являются ключевыми районами для экосистемных услуг. Горные районы обычно имеют богатые и уникальные таксоны пыльцы [7]. Как правило, пояса горной растительности в засушливой Центральной Азии имеют различные типы: от пустынных до пастбищных, лесных и альпийских лугов по мере подъема высоты [8]. Среди них горные лесостепи, обычно расположенные между лесным и степным поясами, располагаются на самом сухом краю распространения леса по вертикали [9].,10]. В связи с этим важно понимать динамику экотона горной лесостепи при изменении климата в аридных регионах. Тем не менее, как изменение пыльцы отражает миграцию лесостепного экотона, остается неясным, несмотря на серию традиционных исследований пыльцевых комплексов, проведенных в некоторых горячих точках, включая Тибетское нагорье, Алтайские горы и горы Тайбая [11,12]. ,13].
Разнообразие таксонов пыльцы может служить комплексным индикатором, отражающим динамику растительных поясов [4]. Увеличило или уменьшило разнообразие таксонов пыльцы в прошлом изменение климата, еще не полностью определено, и изменения в разнообразии таксонов пыльцы потенциально могут включать ключи к изменению растительного пояса, особенно для миграции горно-лесостепного экотона. Таким образом, голоцен является идеальным периодом для изучения миграции лесостепного экотона, при котором изменение климата в основном было обусловлено природной изменчивостью.
Кроме того, важно сосредоточить внимание на способах реагирования поясов горной растительности на голоценовые изменения климата и усовершенствовать соответствующие механизмы, которые являются ключевыми для изучения истории лесостепной миграции. Положение горного вертикального пояса растительности очень чувствительно к изменению климата [11,12], поскольку вертикальный пояс является воплощением горизонтального пояса (около 1/1000). Вертикальный пояс в засушливых районах уникален, так как верхняя и нижняя границы леса подвержены влиянию температуры и влажности соответственно. Горная растительность реагирует на изменение температуры миграцией положения, а на изменение влажности – изменением видового состава [4,5]. Эти два режима в основном приняты, хотя они могут не полностью объяснить механизм между поясами горной растительности и изменениями температуры и влажности. Исследования показали, что ширина пояса горной растительности и климат не находятся в полном равновесии [14,15]. Поскольку климатический режим может быть теплым и сухим, теплым и влажным, холодным и сухим, холодным и влажным, то и верхняя, и нижняя границы лесного пояса, находящиеся в разных климатических условиях, могут реагировать по-разному [12], что в конечном итоге приводит к возможным изменениям ширины и положения ремня. Таким образом, изменение ширины растительных поясов также может быть одной из наиболее типичных реакций на конкретное изменение климата и в дальнейшем может приводить к изменению экотона лесостепи. Следовательно, важно выяснить, существуют ли такие модели реагирования в поясах растительности и могут ли они быть обнаружены посредством изменения разнообразия таксонов пыльцы в засушливых горных районах.
Тяньские горы, крупнейшая горная система в засушливых районах мира, расположены в глубине Евразии, подвержены влиянию западных ветров [16,17,18], где очевидна вертикальная дифференциация растительных поясов и типичный горный лес -степь существует [19]. Из-за сложных топографических условий, ограниченного размера ареала и засушливой среды лесостепной экотон в горах Тянь, по-видимому, наиболее подвержен угрозе ухудшения климата. Однако немногие исследования были специально сосредоточены на миграции лесостепного экотона в ответ на влияние изменения климата в горах Тянь, несмотря на многие исследования, изучающие изменения климата и растительности в различных временных масштабах [20, 21, 22, 23]. ]. Таким образом, существует острая необходимость всестороннего изучения характера реакции лесостепного экотона в горах Тянь на голоценовое изменение климата на основе данных как поверхностной пыльцы, так и ископаемой пыльцы, поскольку пыльца может обеспечить долгую историю разнообразия таксонов пыльцы с хорошая временная преемственность [4].
Основываясь на приведенном выше описании, мы стремимся исследовать следующие научные вопросы: (1) Как развивалось разнообразие горных таксонов пыльцы в течение голоцена? (2) Как разнообразие таксонов пыльцы отражало изменения растительных поясов в горах Тянь в течение голоцена? 3. Как мигрировал лесостепной экотон в ответ на голоценовое изменение температуры и влажности в Тяньских горах?
2. Материалы и методы
2.1. Область исследования
Тяньские горы, крупнейшая горная система в засушливых регионах мира, расположены во внутренних районах Евразии под влиянием западных ветров (рис. 1а, б) [16]. Среднегодовое количество осадков на северных склонах изменяется от 175 мм в пустынной зоне до 465 мм в среднегорной зоне [24].
Горы Тянь имеют наиболее типичный и полный спектр вертикальных природных поясов для умеренных засушливых районов по всему миру, отражающий характеристики распространения и изменения в разнообразии таксонов горной пыльцы и экологических процессах. Северный склон Тяньских гор можно разделить на следующие растительные пояса [14] (рис. 1в): разреженная и низкоподушковидная растительность распространена в высокогорных поясах выше 3400 м над ур. м.; от 2700 до 3400 м над ур. м. преобладают альпийские и субальпийские луга; хвойные леса распространены на средних высотах от 1700 до 2700 м над ур. м.; от 700 до 1700 м над ур. м. преобладает типичная степь, представленная степью на солнечном склоне и редколесьем на тенистом склоне; пустынная растительность распространена ниже 700 м над уровнем моря. Примечательно, что экотон лесостепи, также известный как переходная зона между лесным поясом и степным поясом, в настоящее время расположен на высоте около 1700 м над уровнем моря. на северном склоне Тяньских гор [14].
2.2. Исследование современной растительности
Для исследования изменения видового богатства современных растений по градиенту высот были выбраны типовые участки, представляющие характерный состав растительности в каждом растительном поясе, исходя из природно-географической картины Тяньских гор [14]. В гористой местности на высоте от 700 до 1700 м над ур. м. пробы отбирали через каждые 100 м по градиенту высот. Из-за равнинного рельефа в оазисе и пустынной области ниже по течению через каждые 50 км в горизонтальном направлении отбирались участки. Наконец, было обследовано 40 действительных участков. При выборочном обследовании площадь лесного участка составляла 10 м × 10 м, а площадь сенокосного или лугового сообщества — 2 м × 2 м. Затем регистрировали подробную информацию об условиях произрастания растительности, видовом составе, покрытии и высоте.
2.3. Поверхностная пыльца
Образцы поверхностной почвы отбирали через каждые 100 м в диапазоне от 200 до 3500 м над уровнем моря. на северном склоне Тяньских гор, и было получено 34 образца для изучения изменений поверхностного пыльцевого состава и выравненности в каждом растительном поясе [14]. Пыльцу экстрагировали обработкой HCl (10%) и NaOH (10%), затем просеивали через сита 180 мкм и 10 мкм и флотировали тяжелой жидкостью (KI + HI + Zn) по стандартной методике [25]. В каждом образце подсчитывали наличие не менее 250 пыльцевых зерен под оптическим микроскопом при 400-кратном увеличении.
2.4. Ископаемая пыльца
Мы выбрали два типичных участка ископаемой пыльцы с высоким разрешением на северном склоне гор Тянь, чтобы изучить эволюцию разнообразия таксонов пыльцы. Мы оцифровали все таксоны пыльцы с помощью программы GetData Graph Digitizer v2.25 [26] и использовали их в качестве аналитических палеоданных. Учитывая репрезентативность местоположения, возраст, временное разрешение и идентификацию пыльцы ископаемых пыльцевых ядер, следующие два участка могут удовлетворить наши потребности в анализе.
Первый керн отложений, названный SLMH-2009 (44°35′ с.ш., 81°09′ в.д.), был взят из озера Сайрам на высоте 2072 м (рис. 1а,б) [22] и располагался в средненизкая часть современного пояса хвойных лесов. Это ядро было датировано переходом от позднего ледникового периода к раннему голоцену с датами 11 AMS 14 C и имело временное разрешение менее 100 лет для каждого образца пыльцы. Всего авторы проанализировали 150 образцов ископаемой пыльцы, при этом для каждого образца было идентифицировано не менее 400 пыльцевых зерен.
Второй керн отложений, названный А-01, был взят из оз. Айби (44°54′–45°08′ с.ш., 82°35′–83°10′ в.д.) (рис. в поясе пустынной растительности на высоте 200 м над ур. Этот керн был датирован переходом от позднего ледникового периода к раннему голоцену с использованием восьми дат AMS 14 C и имел временное разрешение менее 150 лет для каждого образца пыльцы. Всего авторы проанализировали 195 образцов ископаемой пыльцы, при этом для каждого образца было идентифицировано не менее 350 пыльцевых зерен.
2.5. Индекс разнообразия таксонов пыльцы
Мы выбрали индекс Шеннона-Винера и индекс Симпсона для расчета разнообразия таксонов пыльцы, поскольку эти два индекса включают неоднородность измеряемого растительного сообщества. Они учитывают как богатство, так и выравненность видов в сообществе. В частности, для оценки уровня разнообразия таксонов пыльцы в современной экосистеме и в голоцене использовался индекс разнообразия Шеннона-Винера, который представляет собой всеобъемлющий индекс, отражающий богатство и равномерность. Его формула такова [27].
Pi = ni/N, что указывает на относительное богатство видов.
ni представляет количество особей каждого таксона пыльцы.
N представляет общее количество особей всех таксонов пыльцы в сообществе.
Следует отметить, что сообщества с низким богатством и высокой выравненностью, а также сообщества с высоким богатством и низкой выравненностью имеют низкие индексы разнообразия.
Индекс разнообразия Симпсона также использовался для представления разнообразия таксонов пыльцы. В отличие от индекса Шеннона-Винера, индекс Симпсона измеряет вероятность того, что два случайных объекта из группы пыльцы представляют разные таксоны пыльцы, что можно количественно определить с помощью следующего уравнения [28].
Pi = ni/N, что указывает на относительное богатство видов.
ni представляет количество особей каждого таксона пыльцы.
N представляет общее количество особей всех таксонов пыльцы в сообществе.
2.6. Климатические данные
Изменение температуры в голоцене было получено из интегрированных результатов палеоклиматических записей для 30–90° северной широты [29,30], а изменение влажности в голоцене было получено из интегрированных результатов палеоклиматических записей в засушливой Центральной Азии [16].
3. Результаты
3.1. Изменения видового богатства современных растений вдоль градиента высот
Мы наблюдали увеличение разнообразия современных видов растений от низкогорья к среднегорному району (рис. 2). Что касается оазиса и пустынной территории вниз по течению от 200 до 700 м над уровнем моря, видовое богатство растений было ограниченным, от 4 до 17 (рис. 2). Диапазон от 200 до 400 м над ур. относится к оазисной зоне с относительно большим количеством видов; поэтому наблюдаемые виды растений сильно колебались (рис. 2). С 700 м над ур. к средней высоте видовое богатство растений имеет тенденцию к увеличению, с пиком в лесной полосе на высоте 1700 м над ур. (Фигура 2). Согласно нашим полевым исследованиям, влаголюбивый вид Picea schrenkiana Fisch. и C.A.Mey. доминировал в лесном поясе, а богатство лесного пояса в основном было обусловлено видовым богатством подлеска, которое может достигать почти 40 видов (рис. 2).
3.2. Состав поверхностной пыльцы
Таксоны поверхностной пыльцы в диапазоне 250–3500 м на северном склоне гор Тянь были проанализированы в соответствии с изменением их богатства по градиенту высот (рис. 3). Salix, Ephedra, Caryophyllaceae, Asteraceae, Thalictrum, Ranunculaceae и Rosaceae достигли пика в диапазоне 3500–2700 м. Picea, Poaceae и Ranunculaceae достигали пика в диапазоне 2700–1700 м. Betula, Artemisia, Salix, Tamarix, Ephedra, Thalictrum и Rosaceae достигли пика между 1700 и 700 м. Ulmus, Amaranthaceae, Nitraria и Tamarix достигли пика ниже 700 м.
Эти таксоны были дополнительно разделены на следующие четыре растительных пояса (таблица 1). Альпийские и субальпийские луга в основном включали Salix, Ephedra, Caryophyllaceae, Asteraceae, Thalictrum, Ranunculaceae и Rosaceae. Эти таксоны могут переносить холодный климат альпийской зоны. Хвойные леса в основном включали Picea, Poaceae и Ranunculaceae, где Picea schrenkiana была доминирующим видом, которому благоприятствовали влажные климатические условия. В типичной степи преобладали Betula, Artemisia, Salix, Tamarix, Ephedra, Thalictrum и Rosaceae. Betula fruticosa Pallas располагалась в полосе пустыни перед горой и относится к категории азональной растительности. Растительность пустыни включала Ulmus, Amaranthaceae, Nitraria и Tamarix. Ulmus pumila L. был распространен в низкогорных речных долинах, также относящихся к азональной классификации растительности. Amaranthaceae, Nitraria и Tamarix обычно являются засухоустойчивыми видами. Основываясь на наших критериях разделения, богатство ископаемой пыльцы в изучаемом осадочном участке, вероятно, происходит из разных поясов растительности, что вскоре будет обсуждаться.
Кроме того, в современной экосистеме индекс Шеннона-Винера и индекс Симпсона постоянно указывали на самое низкое разнообразие таксонов пыльцы в лесном поясе, в то время как самое высокое разнообразие таксонов пыльцы было в поясе альпийских и субальпийских лугов, за которыми следовали типичный степной пояс и пояс пустынной растительности (табл. 2). По результату индекса Шеннона-Винера в верхней (2800–2600 м) и нижней (1800–1600 м) границах и центре лесной полосы (2400–2100 м) разнообразие таксонов пыльцы составило 1,49., 1,65 и 1,21 соответственно (табл. 2). По результату индекса Симпсона в верхней (2800–2600 м) и нижней (1800–1600 м) границах и центре лесной полосы (2400–2100 м) разнообразие таксонов пыльцы составило 0,63, 0,70, и 0,50 соответственно (табл. 2). На основании результатов поверхностных пыльцевых анализов мы пришли к выводу, что разнообразие таксонов пыльцы лесной полосы в целом было низким в горах Тянь, что связано с очень низкой выравненностью пыльцы из-за абсолютного доминирования пыльцы Picea в лесной полосе. (от 39% до 93%, в среднем 67%) (рис. 2 и рис. 3). Таким образом, мы сделали вывод, что лесной пояс с очень низким разнообразием таксонов пыльцы мог влиять на изменения в разнообразии таксонов пыльцы, наблюдаемые на участках отложения в течение голоцена, даже несмотря на то, что в лесном поясе было больше видов растений, чем в поясах типичных степей и пустынная растительность (рис. 3).
3.3. Изменение разнообразия таксонов пыльцы вокруг озера Сайрам в голоцене
В течение голоцена пояса альпийских и субальпийских лугов, хвойных лесов и пустынной растительности вносили вклад в разнообразие таксонов пыльцы вокруг озера Сайрам, в то время как пояс типичной степи не вносил вклада , в соответствии с регрессионными отношениями между разнообразием таксонов пыльцы, указанным индексом Шеннона-Винера, и богатством пыльцы из разных растительных поясов (рис. 4). Линейная регрессия показала, что наибольший вклад в разнообразие таксонов пыльцы вокруг озера Сайрам на протяжении голоцена вносил пояс хвойных лесов (R 2 = 0,40, p < 0,01), тогда как вклад поясов альпийских и субальпийских лугов и пустынной растительности был относительно невелик (R 2 = 0,25, p < 0,01 и R 2 = 0,37, p < 0,01 соответственно) (рис. 4). Это также свидетельствует о том, что разнообразие таксонов пыльцы вокруг озера Сайрам может отражать изменение лесного пояса в течение голоцена.
В теплые и засушливые периоды раннего голоцена, от 12 000 до 8 000 л.н. до н.э. (л.н.) (рис. 5а,б), пояс хвойных лесов сместился вверх, а пояса альпийских и субальпийских лугов и типичных степь также поднялась (рис. 6а), потому что наблюдаемое разнообразие таксонов пыльцы, указанное индексом Шеннона-Винера вокруг озера Сайрам, было очень высоким, достигая в среднем 1,56 (рис. 5в), что было близко к индексу разнообразия (1,65). нижней границы современной лесной полосы (табл. 2). Кроме того, наблюдаемое разнообразие таксонов пыльцы, указанное индексом Симпсона вокруг озера Сайрам, также было очень высоким, достигая в среднем 0,69.(рис. 5г), что было близко к показателю разнообразия (0,70) нижней границы современного лесного пояса (табл. 2). Таким образом, мы делаем вывод, что Сайрамское озеро, вероятно, находилось на нижней границе хвойного леса в начале голоцена, соответственно, лесостепной экотон сместился вверх и приблизился к Сайрамскому озеру.
В среднем голоцене, с 8000 до 4000 лет л.н. (рис. 5а,б), когда климат был теплым и влажным по сравнению с последней стадией, расширялся и преобладал пояс хвойных лесов, а пояса альпийских и субальпийских луга и типичная степь, наоборот, были сжаты (рис. 6b), поскольку наблюдаемое разнообразие таксонов пыльцы, указанное индексом Шеннона-Винера вокруг озера Сайрам, было очень низким, достигая в среднем 1,22 (рис. 5c), что было близко к индексу разнообразия. (1.21) в центре современной лесной полосы (табл. 2). Кроме того, наблюдаемое разнообразие таксонов пыльцы, указанное индексом Симпсона вокруг озера Сайрам, также было очень низким, достигая в среднем 0,51 (рис. 5г), что было близко к индексу разнообразия (0,50) в центре современной лесной полосы ( Таблица 2). Следовательно, озеро Сайрам, вероятно, в среднем голоцене находилось в центре лесного пояса, и соответственно лесостепной экотон, вероятно, сместился вниз (рис. 6б).
По мере того как климат позднего голоцена становился холодным и влажным начиная с 4000 лет назад (рис. 5а,б), пояс хвойных лесов смещался вниз, а также пояса альпийских и субальпийских лугов и типичной степи (рис. 6в), так как наблюдаемое разнообразие пыльцевых таксонов, указанное индексом Шеннона-Винера вокруг озера Сайрам, было относительно высоким, достигая в среднем 1,36 (рис. 5в), что было близко к индексу разнообразия (1,49) верхней границы современного лесополоса (табл. 2). Кроме того, наблюдаемое разнообразие таксонов пыльцы, указанное индексом Симпсона вокруг озера Сайрам, также было относительно высоким, достигая в среднем 0,65 (рис. 5г), что было близко к индексу разнообразия (0,63) верхней границы современной лесной полосы. (Таблица 2). Таким образом, мы делаем вывод, что оз. Сайрам, вероятно, в позднем голоцене находилось на верхней границе хвойного леса, а лесостепной экотон отодвигался дальше вниз и в сторону от оз. Сайрам (рис. 6в).
3.4. Изменение разнообразия таксонов пыльцы вокруг озера Айби в голоцене
В течение голоцена пояса альпийских и субальпийских лугов, хвойных лесов и типичных степей способствовали разнообразию таксонов пыльцы вокруг озера Айби, на что указывала взаимосвязь между разнообразием таксонов пыльцы по индексу Шеннона-Винера, и богатство пыльцы из этих поясов было положительным (рис. 4). Примечательно, что пояс хвойных лесов внес наибольший вклад в разнообразие таксонов пыльцы вокруг озера Айби, согласно результатам нашей линейной регрессии (R 2 = 0,13, p < 0,05), за которыми следуют пояс альпийских и субальпийских лугов (R 2 = 0,12, p < 0,05) и типичный степной пояс (R 2 = 0,09, p < 0,05) (Рисунок 4). Это также предполагает, что разнообразие таксонов пыльцы вокруг озера Айби может отражать изменение лесного пояса в голоцене.
В теплые и засушливые периоды раннего голоцена с 12 000 до 8 000 лет л.н. (рис. 5а,б) пояс хвойных лесов смещался вверх (рис. 6а), а лесной пояс вносил меньший вклад в низкогорное место отложения Айби и больше не доминировало в изменении его разнообразия, поскольку наблюдаемое разнообразие таксонов пыльцы было высоким, со значением 1,35, указанным индексом Шеннона-Винера (рис. 5e). Таким образом, мы делаем вывод, что экотон лесостепи соответственно сместился вверх.
В среднем голоцене с 8000 до 4000 лет назад, когда климат был теплым и влажным по сравнению с последней стадией (рис. 5а,б), пояс хвойных лесов расширился и доминировал (рис. 6б), а лесной пояс сильно пострадал Низкое место отложения на озере Айби, поскольку наблюдаемое разнообразие таксонов пыльцы было низким, со значением 1,30, указанным индексом Шеннона-Винера, и значением 0,62, указанным индексом Симпсона (рис. 5e, f). Следовательно, мы делаем вывод, что экотон лесостепи соответственно сместился вниз.
По мере того как климат позднего голоцена становился холодным и влажным начиная с 4000 лет назад (рис. 5а,б), пояс хвойных лесов смещался вниз, но его площадь уменьшалась с небольшим вкладом по сравнению с последним этапом, поскольку наблюдаемое разнообразие таксонов пыльцы был высоким, со значением 1,46, указанным индексом Шеннона-Винера, и значением 0,64, указанным индексом Симпсона (рис. 5e, f). Следовательно, мы делаем вывод о том, что лесостепной экотон соответственно опустился ниже.
3.5. Сравнение двух озер
Регрессионные зависимости между разнообразием таксонов пыльцы, рассчитанным с использованием индекса Шеннона-Винера, и богатством пыльцы из разных поясов растительности показали, что разнообразие таксонов пыльцы вокруг озер Сайрам и Айби может отражать изменение лесных поясов в голоцене. При расширении лесного пояса преобладала пыльца Picea, что приводило к снижению степени видовой выравненности в хвойном лесном поясе, поскольку разнообразие таксонов пыльцы, наблюдаемое на двух участках, было очень низким.
В целом разнообразие таксонов пыльцы вокруг оз. Сайрам и оз. Айби претерпело одинаковые изменения в ответ на голоценовое изменение климата (рис. 5). В процессе адаптации к теплому и сухому климату раннего голоцена лесной пояс сместился вверх (рис. 5 и рис. 6а), так как в озере Сайрам наблюдалось высокое разнообразие таксонов пыльцы (рис. 5 и рис. 6а). Кроме того, разнообразие таксонов пыльцы озера Айби также было высоким, с меньшим вкладом восходящего лесного пояса (рис. 5 и рис. 6а). Таким образом, экотон лесостепи в раннем голоцене соответственно сместился вверх. В связи с теплым и влажным климатом в среднем голоцене лесной пояс расширился (рис. 6б), так как в озере Сайрам наблюдалось очень низкое разнообразие таксонов пыльцы (рис. 5 и 6б). Кроме того, лесной пояс с преобладанием Picea schrenkiana оказал сильное влияние на низкоуровневое место отложения на озере Айби, поскольку наблюдаемое разнообразие таксонов пыльцы было низким (рис. 5 и рис. 6b). Таким образом, в среднем голоцене экотон лесостепи соответственно сместился вниз. В ответ на холодный и влажный климат в конце голоцена полоса хвойных лесов сместилась вниз (рис. 6с), поскольку в озере Сайрам наблюдалось высокое разнообразие таксонов пыльцы (рис. 5 и 6с). Кроме того, в озере Айби наблюдалось высокое разнообразие таксонов пыльцы с меньшим вкладом сократившейся лесной полосы (Рисунок 5 и Рисунок 6c). Таким образом, в позднем голоцене экотон лесостепи соответственно сместился вниз.
4. Обсуждение
Полученные нами результаты показывают, что изменения экотона лесостепи следовали за голоценовыми изменениями климата в Тяньских горах. По мере изменения степени температуры и влажности менялся лесной пояс, что в дальнейшем определяет позиционную миграцию лесостепного экотона, о чем свидетельствуют изменения в разнообразии таксонов пыльцы, наблюдаемые на участках отложения в горах Тяня (рис. 5 и а). Рисунок 6). Под совместным влиянием солнечной инсоляции и западных ветров изменение климата демонстрирует уникальную картину в засушливой Центральной Азии, характеризующуюся теплым и сухим климатом в раннем голоцене, теплым и влажным климатом в среднем голоцене и умеренно холодным и влажным климатом в течение позднего голоцена (рис. 5а,б) [16,18,19,29,31]. Лесной пояс с преобладанием пыльцы Picea расширился и расширился в течение среднего голоцена, что привело к снижению степени видовой выравненности пояса хвойных лесов, поскольку наблюдаемое разнообразие таксонов пыльцы вокруг двух озер было очень низким. В частности, на северном склоне гор Тянь влажная среда среднегорья благоприятствует существованию Picea schrenkiana, которая в значительной степени влияет на горное разнообразие, что подтверждается нашими результатами по поверхностной пыльце (рис. 3; табл. 2). Действительно, такой тип распространения среднегорных лесов популярен в засушливых и полузасушливых горных районах Китая, что привлекло широкое внимание [5,12,19].,32]. Более того, как лесной пояс, так и лесостепной экотон перемещались вверх в раннем голоцене и вниз в позднем голоцене, что привело к уменьшению влияния лесного пояса на участки, поскольку наблюдаемое разнообразие таксонов пыльцы было высоким (рис. 3; табл. 2). ).
Действительно, голоценовые изменения температуры и влажности в засушливой Центральной Азии повлияли на вертикальную миграцию растительных поясов и их видовой состав в горных районах [18,19]. Верхняя граница лесополосы контролируется температурой, а нижняя – влажностью. При теплом и сухом климате лесной пояс перемещается вверх по возвышенности, чтобы получить больше влаги, и наоборот [11,33]. Видовой состав изменяется от влаголюбивых к засухоустойчивым видам, когда засуха климата превышает физиологические пределы [5]. Кроме того, мы обнаружили, что лесной пояс откликался на теплый и влажный климат через ширину лесного пояса, что в дальнейшем заставляло лесостепной экотон соответственно смещаться вниз. Такой же соответствующий режим обнаружен и в голоценовой смене растительных поясов в бассейне реки Манас на северном склоне Тяньских гор [14]. Поэтому подчеркнем, что миграция лесостепного экотона отреагировала на голоценовое изменение климата по мере изменения лесного пояса, что подтверждается изменением разнообразия таксонов пыльцы в местах отложения.
Хотя разнообразие таксонов пыльцы в засушливых горных районах может отражать изменение растительного пояса (рис. 5 и рис. 6), приводит ли сужение лесного пояса к сокращению разнообразия таксонов пыльцы гор и дополнительно влияет на оценку лесостепного экотона миграция? Если ширина или площадь лесного пояса сужается, то считается, что разнообразие таксонов пыльцы, наблюдаемое в местах депонирования, снижается. Однако подчеркнем, что если сокращается только ширина лесного пояса, а видовое богатство и выравненность леса не меняются, то не обязательно может снижаться разнообразие пыльцевых таксонов. Кроме того, стоит отметить, что некоторые таксоны в лесостепном экотоне могут исчезнуть, когда в засушливых горных районах иссушение климата превысит пороговое значение. Потеря влаги может представлять наибольшую угрозу относительно узкому лесостепному экотону, учитывая климатическую нишу пространства чувствительных к влаге таксонов и ограниченность их жизненного пространства. Например, как одна из горячих точек разнообразия в мире, узкие растительные пояса восточных Анд демонстрируют большую уязвимость к потеплению климата, поскольку ожидаемые темпы изменения климата могут вытеснить узко распространенные таксоны из пространства климатической ниши сотнями. лет [34]. Более серьезно, по мере усиления глобального потепления ожидается, что региональная засушливость станет более серьезной и продлится дольше, чем в нынешнем сухом климате в засушливых регионах [3]. Поэтому представляется, что будущее потепление климата и связанное с ним снижение влажности, вероятно, создадут неожиданные угрозы для таксонов с узким высотным распространением, особенно для таксонов в лесостепном экотоне гор Тянь. Таким образом, мы предлагаем для охраны и поддержания современного экотона лесостепи в горных районах уделять большое внимание влагочувствительным таксонам в условиях засушливого климата.
Кроме того, разнообразие таксонов пыльцы может служить важным индикатором миграции лесостепного экотона [4]. Однако существуют неопределенности, в основном связанные со следующими аспектами. Во-первых, ограничение индикаторов пыльцы — потенциальные неопределенности возникают из-за различий в продуктивности пыльцы и способности к рассеиванию среди разных видов [4,35,36,37], которые могут не отражать один к одному динамику растительных поясов и богатство растительности. ископаемая пыльца. Во-вторых, влияние структуры растительности — для пыльцы, переносимой ветром, ее способность к распространению в лесу гораздо хуже, чем на открытой местности. Это не только влияет на количество пыльцы, которую можно наблюдать на месте отложения, но также влияет на нашу способность восстанавливать растительность, используя данные о пыльце [38]. В-третьих, разные типы пыльцы имеют разную репрезентативность — если сообщество с одинаковым видовым составом имеет разное покрытие, то и выход пыльцы будет разным. При постоянном количестве статистических зерен малорепрезентативные виды могут не учитываться, что повлияет на оценку разнообразия таксонов пыльцы и в дальнейшем повлияет на оценку миграции лесостепного экотона [38]. Поскольку это исследование в основном основано на индексе Шеннона-Винера и индексе Симпсона для расчета разнообразия таксонов пыльцы, которые учитывают как богатство, так и однородность, таксоны с очень низким числом или процентным содержанием пыльцы мало влияют на результаты.
5. Выводы
Мы установили, что лесостепная миграция в целом следовала за голоценовыми изменениями климата в Тяньских горах. В частности, лесной пояс, где доминирует Picea schrenkiana, отличается очень низким разнообразием таксонов пыльцы, характеризующимся высоким богатством и низкой выравненностью, что играет ключевую роль в горном разнообразии. Обнаружив изменение разнообразия мест отложений, мы обнаружили, что в процессе адаптации к теплому и сухому климату в раннем голоцене с 12 до 8 тыс. к местам отложения на низкой высоте, поскольку наблюдаемые различия вокруг двух озер были высокими. При этом экотон лесостепи соответственно сместился вверх. Во время теплого и влажного среднего голоцена с 8000 до 4000 лет назад лесной пояс с низкой таксоновой выравненностью расширялся и расширялся, поскольку наблюдаемое разнообразие вокруг двух озер было очень низким. Таким образом, лесостепной экотон соответственно сместился вниз. Поскольку поздний голоцен стал холодным и влажным, начиная с 4000 лет назад, лесной пояс сместился вниз, что привело к уменьшению влияния лесного пояса на участки или вклада в них, что привело к наблюдаемому высокому разнообразию. Таким образом, экотон лесостепи сдвинулся вниз. Потеря влаги может представлять наибольшую опасность для узкого лесостепного экотона, учитывая пространство климатической ниши и ограниченность жизненного пространства для влагочувствительных таксонов. В данном исследовании показано, что температура и влажность совместно влияют на изменение лесного пояса, что в дальнейшем определяет миграцию лесостепного экотона.
Вклад авторов
Х.Л. и Ю.К. предложил идею и разработал исследование. Ю.К. проанализировал данные и написал первый черновик. YC, HL, HW, QH, YH, KD и З.Д. обсудили результаты и внесли свой вклад в улучшение рукописи. Все авторы прочитали и согласились с опубликованной версией рукописи.
Финансирование
Это исследование финансировалось Национальным фондом естественных наук Китая, номер гранта 417, 41
2. Оно также финансировалось Фондом фундаментальных исследований центральных университетов Китая, номер гранта GK202003069..
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов.
Ссылки
- Bailey, R.M. Пространственные и временные признаки хрупкости и пороговой близости в смоделированной полузасушливой растительности. проц. Р. Соц. Б 2010 , 278, 1064–1071. [Google Scholar] [CrossRef][Green Version]
- Liu, HY; Парк Уильямс, А .; Аллен, CD; Го, Д .; Ву, Х .; Аненхонов О.А.; Бадмаева, Н.К.; Лян, Э .; Санданов, Д.В.; Инь, Ю .; и другие. Быстрое потепление ускоряет снижение роста деревьев в полузасушливых лесах внутренней Азии. Глоб. Чанг. биол. 2013 , 19, 25:00–25:10. [Google Scholar] [CrossRef]
- Хуанг, Дж.; Ю, Х .; Гуань, X .; Ван, Г.; Гуо, Р. Ускоренное расширение засушливых земель в условиях изменения климата. Нац. Клим. Чанг. 2016 , 6, 166–171. [Google Scholar] [CrossRef]
- Weng, C.; Хугиемстра, Х .; Дуивенворден, Дж. Ф. Реакция разнообразия пыльцы на вызванное климатом высотное смещение растительности в колумбийских Андах. Фил. Транс. Р. Соц. В 2007 , 362, 253–262. [Google Scholar] [CrossRef][Зеленая версия]
- Ченг Ю.; Лю, Х .; Донг, З .; Дуан, К .; Ван, Х .; Хан, Ю. Летний муссон в Восточной Азии и топография совместно определяют голоценовую миграцию лесостепного экотона на севере Китая. Глоб. Планета. Чанг. 2020 , 187, 103135. [Google Scholar] [CrossRef]
- Franco, A.M.; Хилл, Дж. К.; Кичке, К.; Коллингем, YC; Рой, Д.Б.; Фокс, Р.; Хантли, Б.; Томас, К.Д. Влияние потепления климата и потери среды обитания на вымирание видов на границах ареала низких широт. Глоб. Чанг. биол. 2006 , 12, 1545–1553. [Google Scholar] [CrossRef]
- La Sorte, FA; Джетц, В. Прогнозируемое сокращение разнообразия видов горных растений в условиях глобального потепления. проц. Р. Соц. Б 2010 , 277, 3401–3410. [Google Scholar]
- Ву, Дж.; Чжан, С .; Цзян Ю. Ботаническая география; China Higher Education Press: Beijing, China, 1995. [Google Scholar]
- Erdos, L.; Амбарлы, Д.; Аненхонов О.А.; Батори, З .; Черхалми, Д.; Поцелуй, М .; Накинежад, А .; Креэль-Дулай, Г.; Лю, Х .; Магнес, М.; и другие. Грань двух миров: новый обзор и синтез по евразийским лесостепям. заявл. Вег. науч. 2018 , 21, 345–362. [Google Scholar] [CrossRef]
- Дай, Дж.; Лю, Х .; Сюй, С .; Ци, Ю .; Чжу, X .; Чжоу, М .; Ву, Ю.; Лю, Б. Дивергентные гидравлические стратегии объясняют межвидовые ассоциации совместно встречающихся деревьев в лесостепном экотоне. Леса 2020 , 11, 942. [Google Scholar]
- Cheng, Y.; Лю, Х .; Ван, Х .; Пяо, С .; Инь, Ю .; Сиаис, П.; Гао, Ю.; Ву, Х .; Луо, Ю .; Чжан, К.; и другие. Контрастное влияние зимнего и летнего климата на эволюцию альпийской границы леса в Восточной Азии, где преобладают муссоны. кв. науч. Откр. 2017 , 169, 278–287. [Google Scholar] [CrossRef]
- Huang, XZ; Пэн, В .; Рудая, Н . ; Гримм, EC; Чен, X .; Цао, X .; Чен, Ф .; Чжан, Дж.; Пан, X .; Лю, С .; и другие. Голоценовая растительность и динамика климата Горного Алтая и прилегающих территорий. Геофиз. Рез. лат. 2018 , 45, 6628–6636. [Google Scholar] [CrossRef]
- Чжао Ю.; Цедакис, ПК; Ли, В.; Цинь, Ф .; Цуй, В .; Лян, К.; Чжао, Х .; Биркс, HJB; Лю, Ю.; Чжан, З .; и другие. Эволюция растительности и изменчивость климата на Тибетском нагорье за последние 1,74 млн лет. науч. Доп. 2020 , 6, eaay6193. [Google Scholar] [CrossRef] [PubMed]
- Джи, З.К.; Лю, Х.Ю. Смещение зон вертикальной растительности в бассейне реки Манас на северном склоне гор Тянь-Шань после позднего ледника. Дж. Палеогео. 2009 , 11, 534–541. [Google Scholar]
- Готфрид М.; Паули, Х .; Фучик, А .; Ахалкаци, М.; Баранчок, П .; Алонсо, JLB; Колдеа, Г.; Дик, Дж.; Эршбамер, Б.; Кальзадо, MRF; и другие. Общеконтинентальная реакция горной растительности на изменение климата. Нац. Клим. Чанг. 2012 , 2, 111–115. [Google Scholar] [CrossRef]
- Чен, Ф.Х.; Ю, З .; Ян, М .; Ито, Э .; Ван, С.; Мэдсен, Д.Б.; Хуанг, X .; Чжао, Ю .; Сато, Т .; Биркс, HJB; и другие. Эволюция влаги в голоцене в засушливой Центральной Азии и ее несовпадение по фазе с историей азиатских муссонов. кв. науч. Ред. 2008 , 27, 351–364. [Google Scholar] [CrossRef]
- Лонг, Х.; Шен, Дж.; Чен, Дж.; Цукамото, С .; Ян, Л .; Ченг, Х .; Фрехен, М. Изменения влажности в голоцене в засушливой Центральной Азии, выявленные на основе всесторонней записи песчаных дюн в центральном Тянь-Шане, Северо-Западный Китай. кв. науч. Откр. 2017 , 174, 13–32. [Google Scholar] [CrossRef]
- Чжан Д.; Чен, X .; Ли, Ю .; Ван, В .; Сан, А .; Ян, Ю .; Ран, М .; Фэн, З. Реакция растительности на эволюцию западных ветров в голоцене в Центральной Азиатской аридной зоне. кв. науч. Rev. 2020 , 229, 106138. [Google Scholar] [CrossRef]
- Sun, X.; Ду Найцю, В.К. Палеорастительность и палеоокружающая среда озера Манаси, Синьцзян, Северо-Западный Китай, за последние 14 000 лет. кв. науч. 1994 , 3, 239–248. [Google Scholar]
- Ли, Дж.; Гоу, X .; Кук, ER; Чен, Ф. Реконструкция засухи на основе годичных колец для центрального района Тянь-Шаня на северо-западе Китая. Геофиз. Рез. лат. 2006 , 33, L07715. [Google Scholar] [CrossRef][Зеленая версия]
- Цзян, QF; Джи, Дж .; Шен, Дж.; Мацумото, Р .; Тонг, Г.; Цянь, П .; Ян, Д .; Рен, X. Растительные и климатические изменения в голоцене в районах Центральной Азии с преобладанием запада, полученные по озеру Сайрам в северном Синьцзяне, Китай. науч. Китайская наука о Земле. 2013 , 56, 339–353. [Google Scholar] [CrossRef]
- Ван, В.; Фэн, З .; Ран, М .; Чжан, К. Изменения климата и растительности в голоцене, сделанные на основе записей пыльцы озера Айби, северный Синьцзян, Китай: потенциальный вклад в понимание модели климата в голоцене в восточно-центральной Азии. кв. Междунар. 2013 , 311, 54–62. [Google Scholar] [CrossRef]
- Чен, X.; Луо, Г .; Ся, Дж.; Чжоу, К .; Лу, С .; Е, М. Экологическая реакция на изменение климата на северном склоне гор Тянь-Шань в Синьцзяне. науч. Китайская наука о Земле. 2005 , 48, 765–777. [Google Scholar] [CrossRef]
- Moore, PD; Уэбб, Дж. А.; Коллисон, М.Э. Анализ пыльцы; Blackwell Scientific Publications: Oxford, UK, 1991. [Google Scholar]
- Cheng, Y.; Лю, Х .; Ван, Х .; Хао, К. Дифференцированная миграция голоценовых биомов в западном и восточном Китае, обусловленная климатом, опосредованная топографией. наук о Земле. Откр. 2018 , 182, 174–185. [Google Scholar] [CrossRef]
- Гофен, М. Влияние температуры на индекс видового разнообразия растений Шеннона-Винера (BDI) зоопланктона в озере Кинерет (Израиль). Откройте J. Mod. гидрол. 2018 , 8, 39–49. [Google Scholar] [CrossRef][Зеленая версия]
- Моррис, Э.К.; Карузо, Т .; Бускот, Ф.; Фишер, М.; Хэнкок, К.; Майер, Т.С.; Майнерс, Т .; Мюллер, К.; Обермайер, Э.; Сочер, С.А.; и другие. Выбор и использование индексов разнообразия: идеи для экологических приложений из немецких исследований биоразнообразия. Экол. Эвол. 2014 , 18, 3514–3524. [Google Scholar] [CrossRef][Green Version]
- Marcott, SA; Шакун, JD; Кларк, PU; Mix, AC Реконструкция региональной и глобальной температуры за последние 11 300 лет. Наука 2013 , 339, 1198–1201. [Google Scholar] [CrossRef][Зеленая версия]
- Чен Ф.; Ву, Д.; Чен, Дж.; Чжоу, А . ; Ю, Дж.; Шен, Дж.; Ван, С.; Хуанг, X. Влажность в голоцене и эволюция летнего муссона в Восточной Азии на северо-востоке Тибетского плато, зарегистрированная у озера Цинхай и его окрестностей: обзор противоречивых косвенных данных. кв. науч. Откр. 2016 , 154, 111–129. [Google Scholar] [CrossRef]
- Чжао Ю.; Ю, З. Реакция растительности на голоценовое изменение климата в муссонно-окраинном регионе Восточной Азии. наук о Земле. Ред. 2012 , 113, 1–10. [Google Scholar] [CrossRef]
- Хао, В.; Лю, Х .; Лю, X. Обнаруженная пыльцой высотная миграция лесов в голоцене в горном лесостепном экотоне на севере Китая. Палеогеогр. Палеоэколь. 2016 , 446, 70–77. [Академия Google] [CrossRef]
- Лю, Х.Ю.; Инь, Ю .; Чжу, Дж.; Чжао, Ф .; Ван, Х. Как лес отреагировал на высыхание голоценового климата в экотоне лесостепи на севере Китая? Кватерн. Междунар. 2010 , 227, 46–52. [Google Scholar] [CrossRef]
- Sugita, S. Теория количественной реконструкции растительности II: Все, что вам нужно, это ЛЮБОВЬ. Голоцен 2007 , 17, 243–257. [Google Scholar] [CrossRef]
- Sugita, S. Теория количественной реконструкции растительности I: Пыльца с больших участков ВЫЯВЛЯЕТ региональный состав растительности. Голоцен 2007 , 17, 229–241. [Google Scholar] [CrossRef]
- Хан Ю.; Лю, Х .; Хао, В.; Лю, Х .; Го, В .; Шангуан, Х. Более надежные оценки продуктивности пыльцы и относительная площадь источника пыльцы в лесостепном экотоне с улучшенным обследованием растительности. Голоцен 2017 , 27, 1567–1577. [Google Scholar] [CrossRef]
- Liu, H.Y. Четвертичная экология и глобальные изменения; China Science Press: Beijing, China, 2002. [Google Scholar]
Рисунок 1. Расположение гор Тянь во внутренних районах Евразии, находящихся под влиянием западных ветров. ( a ) Изображение цифровой модели рельефа изучаемой территории. Красная сплошная точка обозначает расположение озера Сайрам, а синяя — расположение озера Айби. ( б ) Красная пунктирная область представляет диапазон подграфа. Синяя пунктирная линия представляет собой современный предел летнего муссона в Восточной Азии, который также иллюстрирует горизонтальный предел влияния западных ветров. ( c ) Растительные пояса на северном склоне Тяньских гор.
Рисунок 1. Расположение гор Тянь во внутренних районах Евразии, находящихся под влиянием западных ветров. ( a ) Изображение цифровой модели рельефа изучаемой территории. Красная сплошная точка обозначает расположение озера Сайрам, а синяя — расположение озера Айби. ( b ) Красная пунктирная область представляет диапазон подграфа. Синяя пунктирная линия представляет собой современный предел летнего муссона в Восточной Азии, который также иллюстрирует горизонтальный предел влияния западных ветров. ( c ) Растительные пояса на северном склоне Тяньских гор.
Рисунок 2. Богатство современных видов растений вдоль градиента высот. Оранжевая линия представляет собой линию линейной регрессии.
Рисунок 2. Богатство современных видов растений вдоль градиента высот. Оранжевая линия представляет собой линию линейной регрессии.
Рисунок 3. Богатство таксонов поверхностной пыльцы вдоль градиента высот.
Рисунок 3. Богатство таксонов поверхностной пыльцы вдоль градиента высот.
Рисунок 4. Линейные отношения между наблюдаемым процентным содержанием пыльцы из разных поясов растительности и индексом Шеннона-Винера. Примечательно, что горизонтальная ось представляет собой сумму процентного содержания таксонов пыльцы, происходящих из разных растительных поясов (от a до d), наблюдаемых в осадочных участках (конкретные таксоны, содержащиеся в каждом растительном поясе, перечислены в таблице 1). Панель ( A ) представляет озеро Сайрам, а панель ( B ) представляет озеро Айби.
Рис. 4. Линейные отношения между наблюдаемым процентным содержанием пыльцы из разных поясов растительности и индексом Шеннона-Винера. Примечательно, что горизонтальная ось представляет собой сумму процентного содержания таксонов пыльцы, происходящих из разных растительных поясов (от a до d), наблюдаемых в осадочных участках (конкретные таксоны, содержащиеся в каждом растительном поясе, перечислены в таблице 1). Панель ( А ) представляет озеро Сайрам, а панель ( B ) представляет озеро Айби.
Рисунок 5. Изменения разнообразия таксонов пыльцы в горах Тянь и климат голоцена. ( a ) Изменение температуры голоцена для 30–90° с.ш. [29,30]. ( b ) Голоценовые изменения влажности в засушливой Центральной Азии [16]. Серые линии представляют средние уровни индекса разнообразия таксонов пыльцы в разные периоды вокруг озер Сайрам и Айби ( c – f ). Желтая область представляет средний голоцен с низким разнообразием таксонов пыльцы.
Рисунок 5. Изменения разнообразия таксонов пыльцы в горах Тянь и климат голоцена. ( a ) Изменение температуры голоцена для 30–90° с.ш. [29,30]. ( b ) Голоценовые изменения влажности в засушливой Центральной Азии [16]. Серые линии представляют средние уровни индекса разнообразия таксонов пыльцы в разные периоды вокруг озер Сайрам и Айби ( c – f ). Желтая область представляет средний голоцен с низким разнообразием таксонов пыльцы.
Рисунок 6. Схематическая диаграмма изменения разнообразия таксонов пыльцы (ПТД) и миграции лесостепного экотона в условиях меняющегося климата голоцена ( а – с ). В поясе типичной степи степь в основном распространена на солнечном склоне, а лес — на тенистом. Красная точка обозначает местонахождение озера Сайрам, а синяя точка указывает местонахождение озера Айби. Оранжевым эллипсом обозначен возможный диапазон лесостепного экотона.
Рисунок 6. Схематическая диаграмма изменения разнообразия таксонов пыльцы (ПТД) и миграции лесостепного экотона в условиях меняющегося климата голоцена ( а – с ). В поясе типичной степи степь в основном распространена на солнечном склоне, а лес — на тенистом. Красная точка обозначает местонахождение озера Сайрам, а синяя точка указывает местонахождение озера Айби. Оранжевым эллипсом обозначен возможный диапазон лесостепного экотона.
Таблица 1. Списки основных таксонов поверхностной пыльцы, отнесенных к различным поясам растительности вдоль северного склона гор Тянь.
Таблица 1. Списки основных таксонов поверхностной пыльцы, отнесенных к различным поясам растительности вдоль северного склона гор Тянь.
| Альпийские и субальпийские луга 3500–2700 м над ур. | Хвойный лес 2700–1700 м над ур. | Типичная степь 1700–700 м над ур. | Растительность пустыни Ниже 700 м над ур. |
|---|---|---|---|
| Salix Ephedra Caryophyllaceae Asteraceae Thalictrum Ranunculaceae Rosaceae | Picea Poaceae Ranunculaceae | Betula Artemisia Salix Tamarix Ephedra Thalictrum Rosaceae | Ulmus Amaranthaceae Nitraria Tamarix |
Таблица 2. Растительный пояс и разнообразие таксонов пыльцы в современной экосистеме.
Таблица 2. Растительный пояс и разнообразие таксонов пыльцы в современной экосистеме.
| Растительный пояс | Высота (M A.S.L.) | Разнообразие таксонов. | 3500–2700 | 1,84 | 0,78 |
|---|---|---|---|---|---|
| Переходная зона (верхняя граница леса) | 2800–2600 | 1.49 | 0.63 | ||
| Conifer forest | 2700–1700 | 1.16 | 0.49 | ||
| The center of the forest belt | 2400–2100 | 1. 21 | 0.50 | ||
| Transition zone (forest lower boundary) | 1800–1600 | 1.65 | 0.70 | ||
| Typical steppe | 1700–700 | 1.68 | 0.72 | ||
| Desert vegetation | Below 700 | 1.52 | 0.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020, авторы. Лицензиат MDPI, Базель, Швейцария.
Эта статья находится в открытом доступе и распространяется на условиях лицензии Creative Commons Attribution (CC BY) (http://creativecommons.org/licenses/by/4.0/).Геоэкологические параметры указывают на несоответствие между потенциальной и фактической площадью лесов в лесостепи Центральной Монголии | Лесные экосистемы
- Исследования
- Открытый доступ
- Опубликовано:
- Майкл Клинге ORCID: orcid.org/0000-0001-5374-9363 1 ,
- Чоймаа Дуламсурэн 2 ,
- Florian Schneider 1 ,
- Stefan Erasmi 3 ,
- Uudus Bayarsaikhan 4 ,
- Daniela Sauer 1 &
- …
- Markus Hauck 2
Лесные экосистемы том 8 , номер статьи: 55 (2021) Процитировать эту статью
1341 Доступ
3 Цитаты
1 Альтметрика
Сведения о показателях
Абстрактный
Фон
Распространение лесов в лесостепи Монголии зависит от рельефа, вечной мерзлоты и климата и очень чувствительно к изменению климата и антропогенным воздействиям. Лесные пожары и вырубка сократили площадь лесов в лесостепи Монголии. Цель этого исследования состояла в том, чтобы определить геоэкологические параметры, которые контролируют распространение леса и биомассу живых деревьев в этой полузасушливой среде. Основываясь на этих параметрах, мы стремились очертить площадь, которую потенциально может занимать лес, и проанализировать пространственные закономерности фактической и потенциальной биомассы деревьев.
Методы
Мы использовали комбинацию различных географических методов в сочетании со статистическим анализом для определения ключевых параметров, определяющих распределение лесов. В ходе нескольких полевых кампаний мы нанесли на карту биомассу деревьев и экологические параметры в районе исследования в национальном парке Тарвагатай-Нуруу (центральная Монголия). Площадь лесов, топографические параметры и индексы растительности были получены по данным дистанционного зондирования. Существенные корреляции между распределением лесов и биомассой живых деревьев, с одной стороны, и топографическими параметрами, климатическими данными и условиями окружающей среды, с другой стороны, использовались для определения области потенциального распространения лесов и оценки общей биомассы живых деревьев для этой области. .
Результаты
Наличие леса на склонах контролировалось факторами высоты, экспозиции, уклона, среднегодовыми осадками и средней температурой вегетационного периода. Сочетание этих факторов позволило оценить потенциальную площадь лесов, но менее подходило для разграничения древесной биомассы. Не существовало существенных различий в средней биомассе живых деревьев между участками, подверженными различным местным условиям в отношении лесных пожаров, эксплуатации и свойств почвы. Биомасса деревьев снижалась на опушках (определяется как полоса шириной 30 м), в мелких фрагментарных и крупных древостоях. Биомасса деревьев на исследуемой территории составила 20 × 10 9 .0152 9 г (1086 км 2 площади леса), тогда как потенциальная биомасса деревьев может достигать 65 × 10 9 г (> 3168 км 2 ).
Выводы
Полученный прогноз свидетельствует о том, что потенциальная площадь лесов и биомасса деревьев в современных климатических и геоэкологических условиях в три раза превышают нынешнюю площадь лесов и биомассу. Лесные пожары, поразившие в основном крупные лесонасаждения в высокогорье, уничтожили 43% площади лесов и 45% биомассы живых деревьев на исследуемой территории за период86–2017.
Исходная информация
Монгольская лесостепь представляет собой переходную зону между южной границей бореальных лесов в Центральной Азии и засушливым районом пустыни Гоби. Он характеризуется полузасушливым климатом и очень уязвим к изменению климата и интенсификации землепользования (Poulter et al. 2013; Yang et al. 2016; Khansaritoreh et al. 2017b). Различные факторы экологического стресса в последнее время привели к сокращению площади лесов Монголии и, следовательно, скорее всего, биомассы деревьев (Dulamsuren et al. 2010a, 2010b; Hansen et al. 2013). В бореальных лесах Монголии хранится уникальное биоразнообразие лесных видов, которое на региональном уровне исчезает из-за обезлесения (Hauck et al. 2014). Более того, преобразование бореальных лесов в степные пастбища, по оценкам, снижает плотность запасов органического углерода примерно на 40 % не только из-за удаления биомассы, но и в результате потерь углерода из органического слоя (Dulamsuren et al. 2016). .
Неоднократно сообщалось о стрессе от засухи и связанном с ним снижении производства древесины и лесовосстановления, особенно для лиственницы сибирской ( L. sibirica Ledeb.), которая составляет примерно 80% от общей площади лесов (Dulamsuren et al. 2009, 2010a). , 2010c; Лю и др., 2013). Средняя годовая температура воздуха в Монголии увеличивалась на 0,27 К за десятилетие (всего на 1,7 К) с 1940 по 2001 г. (Батима и др., 2005 г.), что явно выше среднемирового показателя в 0,12 К за десятилетие с 1940 по 2001 гг.51 по 2012 г. (МГЭИК, 2013 г.). За период 1940–2006 гг. Dagvadorj et al. (2009) сообщили о сезонном дифференцированном повышении температуры на 0,11 К за десятилетие летом и на 0,51 К за десятилетие зимой. Для этого же периода авторы установили пространственно меняющиеся тенденции увеличения и уменьшения количества осадков.
Кроме того, за последние десятилетия опустошительные лесные пожары потревожили большие лесные массивы в Монголии (Goldammer 2002; Hansen et al. 2013; Nyamjav et al. 2007). Местные власти национального парка «Тарвагатай-Нуруу» в нашем районе исследования заявили, что лесные пожары участились с 19 века.90-х годов, при этом самые сильные пожары произошли в 1996 и 2002 годах. Голдаммер (2002) сообщил, что пожарные с воздушным флотом были установлены с 1969 года до 1990-х годов, когда прекратилась финансовая поддержка из России. Таким образом, с обширными пожарами 1996 и 2002 гг. нельзя было бороться так же эффективно, как с предыдущими пожарами. Кроме того, отсутствие систематического ведения лесного хозяйства, недостаточный контроль за лесозаготовками и выпасом лесов вблизи пастбищ способствовали деградации лесов и сокращению площади лесов (Tsogtbaatar 2004; Dulamsuren et al. 2014). Считается, что широкомасштабные лесозаготовительные работы после лесных пожаров с целью извлечения оставшейся древесины из леса задерживают лесовосстановление из-за механического повреждения молодых деревьев (Сакамото и др., 2021).
Бореальные леса представляют собой важный резервуар органического углерода и, таким образом, важны для глобального климата (Goodale et al. 2002; Pan et al. 2011). Хотя большая часть органического углерода в бореальной зоне хранится в почвах (DeLuca, Boisvenue, 2012; Швиденко, Щепащенко, 2014; Мухортова и др., 2015), значительное количество углерода также хранится в биомассе живых деревьев. Как правило, запасы углерода в общей биомассе деревьев бореальных лесов составляют 40–80 Мг C∙га − 1 (Jarvis et al. 2001; Luyssaert et al. 2007; Thurner et al. 2014). Исследования биомассы деревьев в монгольской лесостепи проводились в Горном Алтае, на юге Хангая (Дуламсурэн и др., 2016), на севере Хангая (Дуламсурэн и др., 2019).) и горы Хэнтэй (Данилин, 1995; Данилин, Цогт, 2014). Полученные значения биомассы деревьев находились в диапазоне 123–397 Мг∙га − 1 и, таким образом (из-за положения Монголии в самой южной бореальной зоне) в пределах и за пределами более высокого диапазона, типичного для бореальных лесов. средней биомассы деревьев от более влажного севера к более сухому югу монгольской лесостепи. В локальном масштабе биомасса деревьев внутри лесов L. sibirica превышает биомассу на опушках (Dulamsuren et al. 2016). Не было обнаружено устойчивых существенных различий в биомассе деревьев между лесонасаждениями разного размера и между лесами, растущими на пастбищных и лесостепных участках лесостепи (Dulamsuren et al. 2019).).
Лесозаготовки, другие виды лесопользования, такие как выпас лесов и разведение костров, с доисторических времен сокращали площадь лесов и биомассу деревьев в Центральной Азии (Miehe et al. 2007, 2014; Unkelbach et al. 2017, 2019). Воздействие этой деятельности можно оценить путем оценки потенциальной площади лесов на основе климатических и топографических параметров (Клинге и др., 2015 г.). Параметры осадков, температуры и испарения контролируют пространственный характер распределения лесов и степей в полузасушливой лесостепи (Нямяв и др., 2007; Дуламсурен и Хаук, 2008; Клинге и др., 2018). Кроме того, важную роль играет топографическое положение, поскольку леса обычно ограничены склонами, обращенными к северу (Клинге и др. , 2015; Хайс и др., 2016). Таким образом, рельеф является важным фактором существования, силы и густоты леса. Помимо природных факторов, на нынешнее распределение лесов сильное влияние оказывает антропогенное воздействие, усилившееся с доисторических времен. Лесозаготовки ведутся бессистемно на древесину и дрова и распространены в лесостепи. Его интенсивность часто возрастала после перехода от плановой экономики к рыночной в XIX в.90-е годы (Дуламсурен и др., 2014). Животноводство кочевников-скотоводов влияет на лесовосстановление на опушках и в глубине мелколесья. В Монгольском Алтае повышенная плотность скота способствовала приживанию саженцев L. sibirica за счет создания просветов в напочвенной растительности, но позднее снизила плотность регенерации деревьев на стадии саженцев, так как саженцы являются предпочтительным рационом козы (Khishigjargal et al. 2013). Поголовье коз в Монголии увеличилось с 19 века.90-х годов из-за высокой экономической значимости кашемировой шерсти для скотоводческих хозяйств (Lkhagvadorj et al. 2013a, 2013b). По сравнению с домашним скотом, выпас диких копытных имеет второстепенное значение из-за более низкой плотности и большого охотничьего давления. Успех регенерации L. sibirica в первую очередь зависит от наличия влаги и травоядности (Dulamsuren et al. 2008; Khishigjargal et al. 2013). Он лишь отдаленно связан с пожаром в монгольской лесостепи, так как сухой климат (поддерживаемый домашним скотом) создает просветы в напочвенной растительности, где могут прижиться саженцы (Данилин 19).95; Дуламсурен и др. 2010b). Хронология годичных колец часто показывает ежегодное укоренение деревьев в течение более длительных периодов с влажным климатом, но не связано с пожарами (Dulamsuren et al. 2010a, 2010c; Khansaritoreh et al. 2017a).
Основываясь на описанном выше уровне знаний, мы рассмотрели следующие гипотезы:
- (Я)
Климатические и топографические параметры, ограничивающие общее распространение лиственничных лесов на исследуемой территории, могут быть выведены путем пространственного анализа данных дистанционного зондирования.
- (II)
Совместное воздействие дополнительных факторов среды (отличных от топоклиматических факторов, контролирующих распространение леса) регулирует реальную биомассу живых деревьев на исследуемой территории.
- (III)
Частые лесные пожары, вырубка леса и выпас леса сильно сократили площадь лесов и биомассу живых деревьев с доисторических времен. Таким образом, леса лишь частично покрывают потенциальную лесную площадь, которую можно вывести из климатических и топографических условий.
Материалы и методы
Район исследований
Район исследований расположен на северной окраине Хангайских гор недалеко от города Тосонценгель в северной центральной Монголии (98°16′ в. д., 48°46′ с.ш.) (рис. 1) . Климат региона континентальный с холодными полузасушливыми условиями (рис. 2). Среднемесячные температуры в Тосонценгеле колеблются от − 31,7 °C в январе до 14,7 °C в июле. Большая часть годовых осадков выпадает летом из ячеек низкого давления, приносимых западными ветрами (Батима и др., 2005). Напротив, Сибирский антициклон зимой вызывает в основном засушливые условия. Холодный климат способствует прерывистой вечной мерзлоте, причем вечная мерзлота в основном встречается в днищах долин, в верховьях гор и частично на склонах. Существование вечной мерзлоты требует некоторой влажности почвы, тогда как сухие почвенные условия приводят к сухой вечной мерзлоте, т. Е. Многолетней мерзлоте безо льда.
Область исследования. а) Обзор Монголии с положением на карте, показанным в б) (черный прямоугольник). б) Расположение исследуемой территории в лесостепи северной центральной Монголии. Распределение лесов было адаптировано из Klinge et al. (2018), площадь выгоревших лесов (2000–2018 гг.) была адаптирована из Hansen et al. (2013). Цифровая модель рельефа (DEM) была создана на основе данных SRTM (Shuttle Radar Topography Mission). Черный прямоугольник в b) указывает положение изображения, показанного в c). в) Цветной космический снимок исследуемой территории вблизи Тосонценгеля (Landsat 8, 22 сентября 2014 г.)
Увеличить
Рис. 2Климат района исследования в районе г. Тосонценгель (черный кружок). Данные из набора данных CHELSA V1.2, период измерений 1979–2013 гг. (Karger et al. 2018), заштрихованная иллюстрация рельефа основана на данных TanDEM-X
Полноразмерное изображение
3200 м над уровнем моря происходят в его южной части. Для них характерны горные плато с террасами криопланации (Рихтер и др., 1963; Ковальковски и Старкель 19).84). Эти самые высокие районы над верхней линией деревьев на высоте ок. 2500 м над уровнем моря относятся к перигляциальному поясу с альпийской растительностью и голыми поверхностями суши, покрытыми каменными обломками (Klinge et al. 2018). В северной части горы ниже, и горная лесостепь покрывает северные склоны вплоть до вершин. Основные долины проходят с юга на север и ведут в текущую с востока на запад долину реки Идер-Гол (Гол: река по-монгольски) на высоте 1600 м над уровнем моря. Геологический фундамент состоит из пермских метаморфизованных осадочных и кислых плутонических пород и Каменноугольные основные породы (АН МНР, АН СССР, 1990). Грубый детрит этих коренных пород образует склоновые обломки, которые часто перемешаны с песчаными или илистыми эоловыми отложениями и покрыты ими.
Густые, обширные леса встречаются к югу от Идер-Гола, тогда как к северу от реки леса более фрагментированы и преобладает степная растительность (Dulamsuren et al. 2019). Для лесостепи Монголии характерна четкая пространственная структура лесов ( L. sibirica ) на склонах северной экспозиции и степи на склонах южной экспозиции (Hilbig 19).95; Третер 1996). Этот характер растительности обычно контролируется небольшим количеством осадков (< 300 мм), высокой эвапотранспирацией и регулируемыми рельефом различиями в инсоляции в средних широтах (Шлюц и др. , 2008; Хайс и др., 2016). Речные леса состоят из ивы ( Salix ), тополя ( Populus ) и лиственницы ( L. sibirica ). Поскольку эти аллювиальные леса поддерживаются грунтовыми водами, они довольно независимы от местных осадков. В котловинах обильны плейстоценовые дюнные поля с разбросанными отдельными старыми лиственницами. Летом происходит много местных лесных и степных пожаров (Goldammer 2007; Hessl et al. 2012). Сильные лесные пожары в 1996 и 2002 уничтожили обширные леса. Многие из этих бывших лесных массивов еще не отросли. В советское время в Тосонценгельском районе были построены лесозавод и лесовозные пути, что способствовало усилению лесопользования с 1960-х годов. О прежних сплошных рубках до сих пор свидетельствуют сгнившие пни в лесах. Промышленные лесозаготовки были прекращены после политических изменений в начале 1990-х годов, но в некоторой степени возобновились. Незаконные рубки ведутся выборочно внутри лесов и затрагивают отдельные остатки на гарях, но массовые сплошные рубки не проводятся. Кроме того, местное население выборочно добывает в лесах дрова.
Дистанционный анализ распределения лесов, категорий лесов и ландшафтных единиц
Этот этап работы имел решающее значение, поскольку основной целью данного исследования было выявление возможного несоответствия между фактической и потенциальной площадью лесов. Входные данные для этого этапа работы включали изображения Landsat 5, Landsat 8 и Sentinel 2 (рис. 3, первая строка). Для картографирования лесных массивов мы применили полуавтоматический подход после Klinge et al. (2015). Ручное картографирование лесных насаждений по спутниковым снимкам было подкреплено предыдущей контролируемой классификацией максимального правдоподобия, в которой обучающие выборки были выделены для леса, степи и водоемов. Мы использовали спутниковый снимок Landsat 5 от 23 сентября 19 г.86 для определения распределения лесов до масштабного уничтожения лесов в результате пожаров. Это изображение 1986 года было лучшим изображением за период до начала масштабных лесных пожаров. Реальную площадь леса мы определили путем объединения нескольких сцен Landsat 8 (14 мая 2013 г.; 20 июня 2015 г.) и Sentinel 2 (14 сентября 2016 г.; 19 сентября 2017 г.). После пересечения классификации мы провели интенсивную визуальную проверку всех полигонов леса, чтобы удалить и отредактировать неправильно классифицированные участки. В лесостепи четкие границы между участками леса и пастбищами позволили составить очень точные карты лесов. Как показано Klinge et al. (2015), классификация изображений в сочетании с ручным постредактированием приводит к общей точности > 0,9.9. По разнице площади лесов между снимками 1986 и 2017 гг. мы рассчитали площадь выгоревших лесов за этот период. Из-за разного пространственного разрешения космических снимков расхождения менее 20 м на опушках леса не учитывались.
Ход исследования
Изображение в полный размер
Мы выделили несколько категорий лесов, которые встречаются в разных ландшафтных единицах. По соотношению леса и степи мы различали лесонасаждения в лесопреобладающей зоне и степной преобладающей зоне. Район, где горные вершины возвышаются над верхней границей леса, определялся как высокогорный. Леса на равнинных участках (< 2°) вдоль рек интерпретировались как аллювиальные леса. Лесонасаждения на дюнах были непосредственно идентифицированы на космических снимках. Кроме того, на лесостепной и степной территориях выделено четыре класса полноты леса соответственно: F1, G1 ≤ 0,1 км 2 ; F2 = 0,1–1 км 2 ; F3 = 1–5 км 2 ; F4 ≥ 5 км 2 , используя пространственный буфер 30 м, чтобы отличить опушки леса от внутренней части. Эти схемы классификации были адаптированы из Dulamsuren et al. (2016, 2019).
Определение подходящих топографических условий и топографических порогов для роста лесов
Рельеф приводит к изменениям местного климата, что является особенно важным аспектом в полузасушливой Центральной Азии (Klinge et al. 2015). Поэтому мы создали цифровую модель рельефа (ЦМР) на основе данных TanDEM-X (рис. 3, первая строка). Из-за высокого горизонтального (10 м × 10 м) и вертикального (< 1 м) разрешения ЦМР расчетная поверхность местности искажалась пологом леса, особенно на опушках древостоев. Поэтому мы использовали карту реальной лесной площади, полученную по космическим снимкам, для коррекции ЦМР в лесах.
Скорректированная ЦМР позволила нам извлечь различные топографические параметры, включая высоту, экспозицию, уклон и инсоляцию. Мы применили инструмент ГИС для оценки совокупного поступления солнечной радиации за период с мая по сентябрь 2017 г., который служил средним вегетационным периодом (MGS). Эти параметры были извлечены для площади лесов с использованием карты лесов 1986 г., чтобы выявить связи между потенциальным распространением лесов и рельефом. Этот подход позволил нам определить подходящие топографические условия и топографические пороги для роста леса из соответствующего диапазона значений. Мы составили карту потенциальных лесных массивов на основе параметров рельефа (PFA r ), предполагая, что рост леса возможен во всех районах в пределах топографических порогов для роста леса, применяя подход Klinge et al. (2015).
Определение подходящих климатических условий и климатических порогов для роста леса
Аналогично описанному выше определению PFA r , мы также проанализировали PFA на основе взаимосвязей между распространением леса и климатическими параметрами (PFA c ). Для этой цели мы использовали климатические данные из набора данных CHELSA V1.2 (Karger et al. 2018), которые мы передискретизировали из первоначального разрешения 30 угловых секунд, чтобы получить разрешение 30 м (рис. 3, первая вставка). Этот повторно проанализированный набор климатических данных позволил нам учесть параметры местности и влияние ветра, что позволило нам получить улучшенное представление климатических условий на рельефной местности (Karger et al. 2017). Мы рассчитали среднегодовые осадки (MAP), среднегодовое потенциальное испарение и среднюю температуру вегетационного периода (MGST, как среднее среднемесячных температур с мая по сентябрь) для периода 1973–2013. Температура и осадки были в значительной степени независимыми (рис. 2), при этом температура следовала вертикальному градиенту, а осадки демонстрировали дополнительный продольный градиент, вызванный западными ветрами. Потенциальное испарение настолько тесно коррелировало с MGST, что мы не включили его в качестве дополнительного параметра для разграничения PFA c .
Климатические параметры были извлечены для лесных площадей с использованием классификации лесов 1986 года с целью выявления взаимосвязей между потенциальным распространением лесов и климатом, а также определения подходящих климатических условий и климатических порогов для роста лесов. Намерение состояло в том, чтобы составить карту потенциальной лесной площади на основе климатических параметров (PFA c ), предполагая, что рост леса возможен во всех районах в пределах климатических порогов роста леса, таким образом, используя тот же подход, что и для карты PFA r .
Анализ биомассы деревьев
Во время полевых работ в 2014–2018 гг. мы определяли биомассу живых деревьев на участках 20 м × 20 м (рис. 3, последний прямоугольник в первой строке) путем измерения диаметра дерева на высоте груди ( dbh) и высота всех живых деревьев, превышающая высоту 4 м. Кроме того, мы учитывали сеянцы, саженцы и деревья < 3 м, тогда как мертвую биомассу мы не учитывали. Мы использовали либо ультразвуковой клинометр Vertex IV и транспондер T3 (Haglöf, Långsele, Швеция), либо лазерный дальномер True Pulse 200 (Laser Technology, Inc., США) для измерения высоты деревьев. Диаметр стебля рассчитывали по окружности стебля, измеренной сантиметровой лентой. Размеры участка определялись рулеткой, а положение углов участка измерялось с помощью GPS с точностью ~ 3 м. Для пространственной привязки данных графика был рассчитан центр между четырьмя углами графика. Для статистического корреляционного анализа между биомассой и данными дистанционного зондирования были интерполированы средние значения пикселей, расположенных в пределах 400-метровой 2 обведите эту точку (рис. 3, верхняя левая сторона). Таким образом, было проанализировано 140 площадок, включающих лесостепные ландшафты, лесные опушки и интерьеры, подножья склонов, средние склоны, верхние склоны, девственные и эксплуатируемые леса, а также древостои на склонах разной экспозицией и различные размеры древостоев. Мы выбрали участки биомассы в соответствии с их репрезентативностью для большей окружающей территории, чтобы минимизировать расхождения между полевыми данными и данными дистанционного зондирования.
Мы применили две аллометрические функции для лиственницы сибирской ( L. sibirica ) в Монголии, опубликованные Battulga et al. (2013) и Dulamsuren et al. (2016) и использовали среднее значение результатов обоих уравнений для оценки надземной и подземной биомассы живых деревьев. Различия в оценках двух функций обсуждаются в Dulamsuren et al. (2016). Мы предположили, что увеличение общей биомассы деревьев за четырехлетний период сбора данных о биомассе в полевых условиях было меньше, чем точность аллометрического метода. Лесонасаждения, в которых пни указывали на вырубку, были нанесены на карту как леса с «рубкой», тогда как древостои без пней были нанесены на карту как леса без рубки. Лесонасаждения, где обгоревшая кора и/или обугленная древесина указывали на бывшие пожары, были нанесены на карту как имеющие «признаки пожара», а те, у которых нет признаков «пожара отсутствуют». Структура наземной растительности, почвенные профили и обнаружение вечной мерзлоты предоставили вспомогательные данные. Структура наземной растительности была важна для оценки того, какая часть NDVI лесных участков приходится на наземную растительность, поскольку сомкнутость древесного полога составляла менее 53%. На 24 участках измеряли индекс листовой поверхности (LAI) с помощью анализатора растительного покрова LI-COR LAI-2200 C (Licor Biosciences, США). Профили почвы использовались для различения почв, образовавшихся в песчаных отложениях и обломках склонов, а также для обнаружения вечной мерзлоты, поскольку вечная мерзлота является решающим фактором для распространения лесов из-за ее сдерживающего эффекта для талой воды из сезонного грунтового льда, сохраняя талую воду доступной для деревьев. Распространение вечной мерзлоты не использовалось для разграничения биомассы и PFA, но ее экологическая обратная связь представляет собой важный вторичный параметр, как показано Klinge et al. (2021).
Классификация данных участка по другим влияющим факторам (таблица 1), помимо рельефа и климата, позволила выявить влияние этих факторов на биомассу деревьев посредством статистического анализа.
Таблица. 1 Средняя биомасса живых деревьев (надземная и подземная) для разных категорий лесов, подверженных различным факторам. Участки, где нельзя было убедиться в возможном влиянии того или иного фактора, из соответствующей части анализа исключались. SE = стандартная ошибка, n = количество участковПолноразмерная таблица
Dulamsuren et al. (2019) уже проанализировали биомассу деревьев в недрах лиственничных лесов на склонах в том же районе исследования, тем самым сосредоточив внимание на лиственничных насаждениях на оптимальной стадии цикла развития леса (Jacob et al. 2013; Feldmann et al. 2018) и исключив помехи от пожара или лесозаготовок. В дополнение к этому исследованию мы также включили лиственничные насаждения, находящиеся под влиянием различных факторов, чтобы также рассмотреть реакцию биомассы деревьев на эти факторы. При этом мы также проверили потенциал методов дистанционного зондирования для масштабирования данных на основе участков до уровня ландшафта. Мы использовали данные о биомассе деревьев 30 L. sibirica участков на склонах из Dulamsuren et al. (2019) и добавлены данные о биомассе деревьев на лесных опушках и других участках L. sibirica с разным размером древостоев (классы F1/G1–F4) и соотношением лесов и пастбищ (преобладающие леса с классами F1–F4). F4 по сравнению с территорией с преобладанием степи с классом G1).
Помимо этих лесов на склонах, мы также анализировали лиственничные леса на аллювиальных песках в поймах рек. Однако их ограниченный размер не позволял получить отдельные наборы данных по внутренней и опушечной части леса. Всего выделено 12 категорий лиственничных насаждений, в том числе дополнительные влияющие факторы внутрилесного массива/опушки склоново-лесных категорий F1, F2, F3, F4 и G1, а также пойменные леса как независимые вырубленные и сгоревшие/несгоревшие древостои как ковариаты по наличию/отсутствию пней и гари. Обычный ветровал, создающий одиночные валежники внутри естественных лесов, не считался нарушением.
Мы рассчитали среднюю биомассу живых деревьев для каждой категории леса (в зависимости от различных факторов), учитывая, например, вырубку, индикаторы пожаров и состояние верхнего слоя почвы (таблица 1, таблица S1). Мы проверили данные о биомассе деревьев для каждой категории леса на предмет нормального распределения и проверили различия в биомассе живых деревьев между категориями леса на статистическую значимость, используя критерий множественных диапазонов Дункана, рассчитанный в SPSS.
Мы умножили площадь каждой категории леса на среднюю биомассу деревьев этой категории леса, используя три сценария, а именно: i) фактическую площадь леса, ii) площадь леса 1986, и iii) потенциальная площадь лесов (PFA). Разграничить потенциальную площадь аллювиальных лесов было невозможно, потому что схема распределения аллювиальных лесов в значительной степени контролировалась динамикой эрозии и отложений разветвленных рек.
Результаты
Пространственные закономерности распространения лесных пожаров и вечной мерзлоты
Распространение лесных пожаров
Высокогорный район в южной части исследуемой территории потерял большую часть леса в результате пожаров за последние десятилетия (рис. 4). Его некогда большие леса превратились в многочисленные мелкие и разрозненные лесные остатки. В лесопреобладающих центральных и северо-восточных частях исследуемой территории наиболее обширные гари находились в высокогорных районах. На степных участках в северо-западной и восточной частях исследуемой территории сгорело лишь несколько лесных насаждений.
Рис. 4 Ландшафтные единицы на изучаемой территории с фактической площадью лесов и площадью выгоревших лесов (базовый 2018 год). Заштрихованная иллюстрация рельефа основана на данных TanDEM-X
Полноразмерное изображение
Распространение вечной мерзлоты
Вечная мерзлота была ограничена большими лесными насаждениями на склонах в области с преобладанием леса и высокогорной области, как видно из наших профилей почвы ( Клинге и др., 2021). Под крупными насаждениями (класс лесистости F4) на склонах северной экспозиции многолетняя мерзлота была богата льдом и залегала уже на небольшой глубине, тогда как на склонах восточной и западной экспозиций имелись лишь небольшие участки вечной мерзлоты, которые начинались на глубинах более чем 1 м. Полевых данных о наличии вечной мерзлоты под фрагментированными лесонасаждениями (G1, F1, F2) и выгоревшими лесами не было (Klinge et al. 2021).
Разграничение ЗПП по параметрам рельефа (ЗПП
r ) Верхняя граница леса в районе исследования поднимается с 2400 м на севере до 2600 м над ур.м. на юге (Клинге и др., 2018). С 1986 г. лесные пожары в высокогорье привели к снижению среднего диапазона высот лесов (95%) до 1600–2400 м над ур. м. Южные склоны в лесостепи в основном покрыты степной растительностью; они могут быть частично покрыты лесом выше 2100 м над уровнем моря. (рис. 5). Леса также встречаются в районах максимальной инсоляции МГС, что свидетельствует о том, что инсоляция не является лимитирующим фактором для роста лесов на исследуемой территории.
Вверху — Изменение максимальной инсоляции MGS (левая ось) с высотой (MGS = средний вегетационный период, май–сентябрь). Сплошные линии = инсоляция МГС на лесной площади, пунктирные линии = инсоляция МГС на всей поверхности земли. Внизу — площадь лесов в гектарах (правая ось) в 1986 г. в зависимости от высоты. * Площадь лесов на южных склонах показана в га × 10
Полноразмерное изображение
Мы выделили PFA r , вырезав участок, где все три параметра «экспозиция» (на склонах нет леса ниже 2100 м над ур. экспозицией 135–225°), «градиент склона» (0–25°) и «возвышение» (верхняя граница леса: 2400 м над ур. м. на севере, 2600 м над ур. м. на юге) учитывали рост деревьев (рис. 3). ). Самый яркий результат этого PFA 9Проекция 0870 r представляла собой гораздо более обширный лесной покров на подошвенных склонах и фронтонах, в настоящее время в основном покрытых степной растительностью (рис. 6). В высокогорной области расчетная PFA r превышала нынешнюю лесную площадь на крутых склонах, от долин до верхней границы леса на высоте 2600 м над уровнем моря.
Распределение лесов в 1986 г. и потенциальные площади лесов (PFA), очерченные в зависимости от климата и рельефа. Заштрихованный рельеф основан на данных TanDEM-X
Полноразмерное изображение
Разграничение PFA на основе климатических параметров (PFA
c ) Пространственная передискретизация климатических данных с помощью линейной интерполяции произвела некоторый шум, поскольку небольшие топографические вариации не могли быть учтены. Поэтому мы не использовали полученный набор климатических данных для вывода климатических порогов роста лесов с помощью анализа гистограмм. Вместо этого мы получили эти пороги путем ступенчатой корректировки климатических порогов до тех пор, пока пространственная картина температуры и осадков с положительными условиями для роста деревьев не включала все древостои 1986. Климатические пороги, полученные с помощью этого подхода, перечислены в Таблице 2. Полученный PFA c показал большие площади леса на верхних южных склонах и небольших плоских вершинах. Он хорошо совпадал с верхней линией деревьев в высокогорной местности (рис. 7). По сравнению с PFA r , PFA c не распространялся так глубоко в бассейны, что может быть связано с малым количеством осадков.
Полная таблица
Рис. 7 Кривые повторяемости распределения средней температуры вегетационного периода (СГСТ) и среднегодового количества осадков (САД) в районе исследования. Сплошные линии = площадь леса, пунктирные линии = общая площадь. Источник климатических данных: CHELSA V1.2 (Karger et al. 2017), период 1979–2013 гг., пространственная передискретизация до 30 м с помощью линейной интерполяции 12 и 20 м, тогда как максимальная высота одиночных деревьев достигала 32,7 м. Базальная площадь стояния колеблется от 5 до 91 м 2 ·га − 1 , в среднем 38,8 м 2 ·га − 1 . Средний возраст деревьев составлял 100–200 лет, при этом максимальный возраст деревьев достигал 380–413 лет. Биомасса живых деревьев в лиственничных лесах на склонах колеблется от 25 до 380 Мг·га − 1 . Максимальная биомасса деревьев 440–688 Мг·га − 1 была обнаружена в пойменных лесах, в то время как лиственницы на песчаных дюнах формировали только редколесья с меньшей биомассой деревьев (48 Мг·га − 1 , n = 1). Во всех классах древостоя лесов лесостепной зоны средние и медианы древесной биомассы находились в пределах 180–220 Мг·га − 1 ; максимальная биомасса деревьев превышала 320 Мг·га − 1 (табл. 1). Критерий множественных диапазонов Дункана не подтвердил статистически значимых различий между категориями леса, за исключением разницы между лесными опушечными участками G1 и внутренними участками F1 и F2 (рис. 8). Леса размерных классов F1, F2 и F3 имели 50–63 Мг·га − 1 меньше биомассы живых деревьев на опушках, чем в их недрах; только класс размера леса F4 не показал четкой разницы в биомассе деревьев между опушками и внутренней частью леса (табл. 1). Мелкие фрагментарные леса G1 в степной зоне имели на 70 Мг·га − 1 меньшую биомассу деревьев, чем в лесной. Вырубка леса, лесной пожар и тип отложений не оказали существенного влияния на биомассу живых деревьев. Большая доля вырубленных лесов в категориях мелкофрагментарных лесов (размерные классы F1 и G1) указывала на более высокую эксплуатационную нагрузку на эти мелколесья по сравнению с более крупными насаждениями.
Диаграммы биомассы деревьев (Мг·га − 1 ) лесов, различающихся по краевым эффектам, влиянию пожаров, рубок и типу наносов. Горизонтальная линия = медиана, столбцы = квартили, усы = диапазон, точки = выбросы. Средние значения имеют общую букву, существенно не различаются (критерий множественного диапазона Дункана, p ≤ 0,05)
Полноразмерное изображение
Анализ дистанционного зондирования на основе оценок биомассы деревьев
Мы протестировали несколько наборов данных NDVI и различные топографические параметры для значительных одиночных или множественных корреляций с измеренной биомассой деревьев, поскольку такие корреляции позволят интерполировать биомассу деревьев в масштабе ландшафта. Однако мы не обнаружили статистически значимых корреляций ( r > 0,5, p < 0,05). Одной из проблем для анализа NDVI было небольшое количество мультиспектральных спутниковых изображений с облачным покровом < 10%, сделанных летом. Еще одной проблемой была слабая корреляция между объемом хвои и биомассой дерева, поскольку листья и хвоя обеспечивают сигнал хлорофилла на мультиспектральных спутниковых снимках. Данилин и Цогт (2014) установили, что биомасса хвои не зависит от среднего возраста лиственничного древостоя, тогда как биомасса деревьев увеличивается с возрастом деревьев. Отсутствие значимой корреляции между индексом площади листьев (ИЛП) и биомассой деревьев, измеренной на 24 участках, подтвердило это утверждение (рис. 9).). В целом, статистические связи между NDVI, объемом хвои и биомассой деревьев были плохими.
Взаимосвязь между индексом листовой поверхности (LAI) и биомассой живых деревьев (левая ось, синие точки), а также между LAI и NDVI м (правая ось, красные точки) на 24 участках. NDVI м = среднее значение NDVI, полученное по семи снимкам Sentinel 2 (рис. 3)
Полноразмерное изображение
Обобщение результатов оценки лесистости и древесной биомассы
Площадь исследуемой территории составила 6355 км 2 . Закрытая треть общей площади (1898 км 2 ) все еще была покрыта лесами в 1986 году. С тех пор площадь лесов сократилась до 1086 км 2 (фактическая площадь лесов). Очерченный PFA дал 3168 (PFA c ) и 3553 км 2 (PFA r ) соответственно. Подробная информация о различиях между фактической площадью лесов, площадью лесов в 1986 г. и ПЗП приведена в Таблице 3. Наибольшая часть фактической площади лесов относится к классу размера леса F4 (8,6%) на территории с преобладанием леса. До крупных пожаров этот класс полноты леса был также широко распространен в высокогорной местности (9.2%). Всего с конца прошлого века пожарами была уничтожена лесная площадь площадью 812,5 км 2 (12,8% от общей площади исследований и 42,8% от ранее покрытой лесом площади). Площадь выгоревших лесов была ничтожно мала в степной зоне и незначительна в лесной, но составляла до 95% ранее покрытой лесом площади в высокогорной зоне. Соотношения между внутренней частью леса и лесной опушкой (F i /F e ) для каждого класса размера леса были относительно постоянными для различных единиц ландшафта (таблица 3).
Полная таблица
Срок из-за низкой корреляции между топографическими параметрами, NDVI и биомассой живых деревьев (рис. 9) оценка общей биомассы живых деревьев в районе исследования с использованием функций регрессии была невозможна. Поэтому мы оценили общую биомассу живых деревьев, умножив конкретную среднюю биомассу деревьев каждой категории леса на площадь этой категории леса в 1986 и 2018 г. (таблицы 3, 4, 5). Общая биомасса живых деревьев PFA была рассчитана на основе средней биомассы живых деревьев в районе с преобладанием леса, поскольку оба прогноза PFA привели к ландшафтам с преобладанием леса (таблица 5). Фактическая биомасса деревьев в районе исследований составила 57% от биомассы 1986 г., что соответствует 30% и 34% биомассы деревьев, оцененной для PFA r и PFA c соответственно (табл. 5). Наибольшие потери биомассы живых деревьев из-за лесных пожаров с 1986 были обнаружены в больших лесах (размерный класс F4) в лесопреобладающей зоне и высокогорной зоне, тогда как потери древесной биомассы в результате пожаров были менее значительными в степной зоне и аллювиальных лесах.
Полная таблица
Табл. 5 Потенциальная площадь лесов (PFA) и биомасса живых деревьев в зависимости от климата (PFA c ) и рельефа (PFA r )Полная таблица
Дискуссия
Распространение лесов
Основными природными факторами, определяющими пространственное распределение и жизнеспособность лесов в монгольской лесостепи, являются малое количество осадков и высокая эвапотранспирация. Последнее зависит от инсоляции, которая, в свою очередь, зависит от рельефа, что приводит к отсутствию лесов на южных склонах (Dulamsuren and Hauck, 2008; Hais et al. , 2016; Klinge et al., 2018). Там, где есть леса, полог леса способствует густой растительности и органическому поверхностному слою, который изолирует почву от теплого воздуха летом (Dashtseren et al. 2014). Таким образом, леса поддерживают прерывистую вечную мерзлоту (Klinge et al. 2021). В свою очередь, вечная мерзлота помогает деревьям пережить летние засухи, поскольку она предотвращает просачивание талой воды, которая высвобождается над поверхностью вечной мерзлоты, ниже корневой зоны (Sugimoto et al. 2002). Признание этих взаимосвязей имеет решающее значение для понимания закономерностей распределения лесов и древесной биомассы в монгольской лесостепи. Однако сама по себе эта причинно-следственная сеть не может объяснить нынешнюю модель распределения лесов, поскольку на нее дополнительно влияют другие, в основном антропогенные, факторы, которые могут привести к расхождению между фактической и потенциальной площадью лесов (PFA).
Потенциальная площадь лесов (PFA)
Потенциальная площадь лесов с ограниченным рельефом (PFA r ), полученная в результате этого исследования, предполагает возможность расширения лесов как вниз по склону к котловинам, так и вверх по склону к высокогорной местности. Фронтоны, широко покрывающие подошвенные склоны в районе исследований, в целом обеспечивают подходящие геоэкологические условия для произрастания деревьев, что подтверждается несколькими существующими здесь небольшими насаждениями. Тем не менее на фронтонах преобладает степная растительность, в основном из-за выпаса травоядных (Hilbig 1995). Потенциальная площадь леса с ограниченными климатическими условиями (PFA c ), полученная в результате этого исследования, дала более низкую линию деревьев, где сухие условия бассейнов препятствуют росту деревьев, что совпадает с нынешними нижними границами леса. Существование лесов ниже порога 160 мм ПДК можно объяснить дополнительным водоснабжением за счет боковых потоков воды, аккумулирующихся в вогнутых местах и подножиях склонов (Клинге и др., 2021). Кроме того, PFA c предполагает наличие больших лесных массивов на южных склонах. Это несоответствие еще раз подтверждает, что MAP и MGST сами по себе не могут объяснить распределение лесов, которое является результатом более сложной причинно-следственной сети, как объяснялось в начале этой главы. Короткий вегетационный период и продолжительный снежный покров препятствуют распространению лесов в верховья долин и на горные плато высокогорной области на юге. Еще одним ограничивающим фактором является широкое использование альпийских лугов в качестве летних пастбищ. Верхняя линия деревьев поднимается с высоты 2400 м над уровнем моря. на севере до 2600 м над ур. на юге изучаемой территории. Таким образом, небольшие безлесные участки на плоских вершинах северных гор могут быть результатом так называемого «эффекта вершины» (Körner 2012), т. е. особенно суровых условий вблизи вершин, а не истинной верхней границы леса. Прогнозируемые PFA предполагают более крупные леса и значительно меньше мелких, фрагментированных лесонасаждений на территории с преобладанием степей в северной части изучаемой территории, при условии, что там преобладают леса. Переход от потенциальной лесной территории к наблюдаемой в настоящее время степной зоне мог быть частично вызван такими природными факторами, как пожар, бурелом, нашествие насекомых и засуха, но вырубка леса и выпас лесных пастбищ, скорее всего, вызвали большие потери лесов в этой области. . Учитывая влияние больших лесов на вечную мерзлоту, предлагаемый сценарий территории с преобладанием леса для северной части изучаемой территории будет также включать большее распространение вечной мерзлоты в этой области.
Воздействие пожара
Площадь лесов, сгоревших в период с 1986 по 2017 год, составляет 12,8% от общей площади исследований. Потеря биомассы живых деревьев с прошлого века составляет примерно 15 миллионов тонн, что составляет более 45% прежней биомассы деревьев. Нямжав и др. (2007) заявили, что 95% фактического уничтожения лесов было вызвано лесными пожарами, а 5% — вырубкой. Авторы сообщают об увеличении пожаров в Монголии за последние десятилетия. Goldammer (2002) предположил, что большинство пожаров было вызвано деятельностью человека. Хессл и др. (2012) исследовали историю пожаров за последние 450 лет на основе анализа годичных колец деревьев. В соответствии с результатами, полученными в Тувинском регионе на юге Сибири (Иванова и др., 2010), авторы не обнаружили увеличения частоты пожаров в последние десятилетия, но пожары стали более сильными из-за более засушливых условий. Ограниченная одновременность возникновения пожаров на разных участках указывала на то, что огонь разжигали люди (Hessl et al. 2012). С другой стороны, антропогенное воздействие также может привести к уменьшению разрушительного действия пожаров, поскольку сбор дров и интенсивный выпас скота сокращают доступное топливо для пожаров (Umbanhowar et al. 2009).; Хессл и др. 2012).
Несмотря на то, что самые масштабные лесные пожары в этом районе произошли уже в 2002 г., на многих сгоревших участках лес еще не восстановился. Наиболее обширные гари расположены в крупных лесах высокогорной зоны и в высокогорье в лесостепной зоне. Напротив, в небольших и фрагментарных лесах степной зоны встречается лишь несколько выгоревших участков леса. Сделан вывод, что фрагментация лесов в степной зоне препятствует переходу лесных пожаров на соседние древостои и держит пожары в достаточной степени изолированными. Сокращение больших лесов (класс F4) в результате пожара привело к увеличению мелких, фрагментированных древостоев (класс F1), представляющих собой остатки бывших больших лесов. Это изменение вызвало потерю вечной мерзлоты в этих районах. У уцелевших лиственниц в оставшихся лесных остатках наблюдается усиленное плодоношение (Данилин, Цогт, 2014). Об их важной роли как ядер лесовосстановления свидетельствуют многочисленные сеянцы и саженцы, растущие в непосредственной близости от остатков леса, в тени старых деревьев. Таким образом, из этих лесных остатков происходит медленная, но неуклонная реиммиграция лиственниц на гари. Может пройти до 200 лет, прежде чем лес восстановится до состояния, существовавшего до пожара (Нямяв и др., 2007).
В полузасушливой среде часто возникают пожары (Hessl et al. 2012). Таким образом, L. sibirica в определенной степени приспособлен к огню. Его выживание при пожаре зависит от типа пожара (коронный, поверхностный или низовой пожар), интенсивности пожара, времени года и влажности почвы. Преобладающее сохранение лесных насаждений в понижениях, эрозионных каналах и на склонах склонов свидетельствует о важности влажности почвы для выживания деревьев.
Биомасса живых деревьев
В отличие от тесной связи распространения лесов с рельефом и климатом, биомасса живых деревьев не показала существенной связи с топографическими параметрами. Оказалось, что леса монгольской лесостепи имеют очень изменчивую биомассу живых деревьев. В дополнение к естественным воздействиям на леса (например, пожары, буреломы, бедствия, вызванные насекомыми и засухи), на биомассу живых деревьев могут влиять лесозаготовки и выпасы лесов. Аллювиальные леса существуют там, где русла рек мешают пастбищам, лесозаготовкам и лесным пожарам. Эти аллювиальные леса обычно состоят из старых лиственниц и имеют большую древесную биомассу. Редкие лиственничные леса на дюнах также состоят из очень старых деревьев, но имеют низкую полноту древостоя и древесную биомассу.
При попытках оценить биомассу в ландшафтном масштабе в качестве показателя биомассы обычно используется NDVI. Однако его пригодность зависит от масштаба и разрешения данных. Дуламсурен и др. (2016) успешно применили NDVI в региональном масштабе для оценки биомассы в Монголии. Однако в нашем локальном анализе мы не получили статистически значимой корреляции между NDVI и биомассой живых деревьев. Вместо этого NDVI оказался подходящим индикатором условий произрастания всей лесной растительности (Erasmi et al. 2021).
Различия в средней биомассе деревьев между выделенными в нашем исследовании категориями леса составили до 85 Мг·га − 1 . Наши полевые измерения показали, что наименьшая средняя биомасса деревьев приходится на лесостепной класс G1 (142 Мг·га − 1 ) в степной зоне и в классе F4 (182 Мг·га − 1 ) на степной территории. территория с преобладанием леса. Уменьшение биомассы живых деревьев в небольших, фрагментированных лесах на территории с преобладанием степей (G1) может быть объяснено более активным лесопользованием, уменьшающим базальную площадь древостоя. Так как поэтому солнечная радиация на землю повышена, под этими лесами нет вечной мерзлоты. Снижение биомассы живых деревьев крупнейших лесов (размерный класс F4) имеет другую причину. Здесь кровля вечной мерзлоты приближается к поверхности и препятствует глубокому укоренению деревьев. Таким образом, деревья очень подвержены ветровым выбросам, что объясняет снижение биомассы живых деревьев в этой категории лесов.
Кроме того, на опушках леса наблюдалось снижение биомассы живых деревьев. Лесные опушки представляют собой естественные зоны расширения и отступления леса (Sommer and Treter, 1999), при этом временные климатические изменения контролируют эти колебания. Опушки леса могут иметь опушку из мертвых деревьев на их внешней границе, а их внешняя граница также может быть расчленена. Кроме того, лесозаготовки и выпас скота более интенсивны на опушках, чем во внутренних районах. Из-за меньшей плотности деревьев биомасса живых деревьев на опушках обычно ниже, чем внутри. Однако мы обнаружили исключения из этого правила в классах размера леса G1 и F4, где недра и опушки леса имели сходную биомассу деревьев. В классе G1 биомассы деревьев малых внутренних лесных массивов столь же низки, как и на опушках. В классе F4 опушки леса имеют большую биомассу деревьев по сравнению со всеми остальными опушками леса. Это можно объяснить влиянием вечной мерзлоты. Неглубокий слой вечной мерзлоты во внутренней части лесов F4 вызывает снижение биомассы деревьев во внутренней части леса, как описано выше. К опушкам леса мощность кровли вечной мерзлоты увеличивается. Там летом вечная мерзлота поставляет талую воду деревьям, которые в остальном близки к климатическому порогу роста деревьев на более сухих опушках леса. Этот эффект, наряду с большим количеством осадков в высокогорье, может также объяснить существование лесов на южных склонах в более высоких горах (рис. 5).
Интересно, что лесонасаждения, подвергшиеся несмертельным пожарам или выборочным рубкам, имели сходную биомассу деревьев с нетронутыми лесами. Возможное объяснение состоит в том, что умеренное изреживание лесов может улучшить условия произрастания оставшихся деревьев, так как приводит к снижению конкуренции за воду и увеличению поступления питательных веществ из пепла, а таяние вечной мерзлоты приводит к временному повышению влажности почвы и позволяет проводить более глубокие посадки. укоренение.
Расчетная максимальная биомасса деревьев ЗСП (58–65 × 10 9 г) вдвое превышала биомассу дерева в 1986 г. (35 × 10 9 г) и в три раза превышала фактическую биомассу дерева, равную 20 × 10 9 г. Однако в прогнозе PFA нельзя было учесть несколько важных факторов. Например, крупные леса (размерный класс F4), преимущественно полученные в результате оконтуривания ЗПП, более подвержены сильным пожарам, чем фрагментированные древостои. Кроме того, из-за длительного влияния человека реконструкция естественного соотношения между степью и лесом в этом регионе остается серьезной исследовательской задачей (Клинге и Зауэр, 2019 г.).). Воздействие человека уже началось с вымирания крупных травоядных, таких как слоны, и сокращения стад диких животных с периода мезолита. Он продолжился разведением домашних животных и развитием пастбищ с эпохи неолита, которая началась около 4,7 тыс. л.н. с афанасьевской культуры в Горном Алтае (Ковалев и Эрдэнэбаатар, 2009).
Чебакова и др. (2009) смоделировали потенциальные изменения растительности в Сибири на основе сценариев изменения климата, предполагающих более теплый и сухой климат. Авторы прогнозируют увеличение площадей лесостепи и пастбищ. Они предполагают, что более сухие условия и большее количество топлива из-за повышенной гибели деревьев приведут к увеличению частоты и разрушительности пожаров.
Выводы
Сочетание определения биомассы деревьев, обнаружения вечной мерзлоты в профилях почвы, дистанционного зондирования и анализа климатических данных позволило нам выявить факторы, контролирующие распространение лиственничных лесов и биомассу живых деревьев в северной части Хангайских гор в центральной Монголии. Выявленные топографо-климатические пороги роста лесов позволили выделить потенциальную площадь лесов (ППЛ), которая значительно превышала фактическую площадь лесов. Лесные пожары уничтожили 43 % площади лесов и 45 % биомассы живых деревьев на изучаемой территории за период 1986–2017. В основном они затронули крупные лесонасаждения в высокогорьях. Вечная мерзлота, широко распространенная под крупными лесами, исчезла вскоре после уничтожения крупного древостоя.
В отличие от распределения лесов биомасса живых деревьев не коррелирует с топографическими и климатическими параметрами. Мы не обнаружили существенных различий в биомассе живых деревьев между лесами с разной пожарной историей, степенью эксплуатации и свойствами почвы, а также между большинством размерных классов леса. Только на опушках и небольших фрагментированных лесонасаждениях биомасса деревьев была значительно меньше, чем во всех других категориях леса. Ни несмертельные пожары, ни выборочные рубки серьезно не уменьшили биомассу живых деревьев. Мы пришли к выводу, что эти воздействия удаляют биомассу деревьев, но также стимулируют рост оставшихся деревьев за счет снижения конкуренции.
Исходя из порогов рельефа для роста леса, мы получили PFA r 3552 км 2 с биомассой деревьев 65×10 9 г, а на основании климатических порогов PFA c 31901 км 2 с биомассой деревьев 58 × 10 9 г, что соответствует 323% и 288% фактической биомассы деревьев соответственно. Однако эти оценки не учитывают несколько важных факторов, таких как выпас травоядных и конкуренция растений. Кроме того, длительное воздействие человека (в масштабе тысячелетия) также играет важную роль в структуре растительности, что требует дальнейшего изучения.
Доступность данных и материалов
Наборы данных, использованные и/или проанализированные в ходе текущего исследования, можно получить у соответствующего автора по обоснованному запросу.
Климатические данные общедоступны из набора данных CHELSA (https://chelsa-climate.org/).
Сокращения
- над уровнем моря:
Над уровнем моря
- дбч:
Диаметр дерева на высоте груди
- DEM:
Цифровая модель рельефа
- LAI:
Индекс площади листьев
- Карта:
Среднегодовое количество осадков
- МГС:
Средний вегетационный период с мая по 9 сентября.
0003- МГСТ:
Средняя температура вегетационного периода
- NDVI:
Нормированный дифференцированный вегетационный индекс
- ПФА:
Потенциальная лесная площадь
- SE:
Стандартная ошибка
- СРТМ:
Миссия шаттла по радиолокационной топографии
Литература
Академия наук Монголии, Академия наук СССР (1990) Национальный атлас Монгольской Народной Республики, Улан-Батор, Москва
Батима П., Нацагдорж Л., Гомблуудев П., Эрдэнэцэцэг ( 2005) Наблюдение за изменением климата в Монголии. Рабочие документы AIACC 12:1–25
Google ученый
Баттулга П.
, Цогтбаатар Дж., Дуламсурэн Ч., Хаук М. (2013) Уравнения для оценки надземной биомассы лиственницы сибирской в лесостепи Монголии. J Forest Res 24 (3): 431–437. https://doi.org/10.1007/s11676-013-0375-4Дагвадорж Д., Нацагдорж Л., Доржпурев Дж., Намхайням Б. (2009) Отчет об оценке изменения климата в Монголии 2009, Улан-Батор, Монголия
Данилин И.М. (1995) Структура и биомасса лиственничных насаждений, естественно возобновляющихся после сплошных рубок. Вода Воздух Почва Загрязнение 82:125–131. https://doi.org/10.1007/978-94-017-0942-2_14
CAS Статья Google ученый
Данилин И.М., Цогт З. (2014) Динамика структуры и биологической продуктивности послепожарных лиственничных лесов северной Монголии. Contemp Probl Ecol 7(2):158–169. https://doi.org/10.1134/S1995425514020036
Артикул Google ученый
Дашцэрен А.
, Исикава М., Иидзима Й., Джамбалжав Й. (2014) Температурные режимы деятельного слоя и сезонномерзлого грунта под мозаикой лесостепи, Монголия. Вечная мерзлота Periglac Proc 25 (4): 295–306. https://doi.org/10.1002/ppp.1824Статья Google ученый
DeLuca TH, Boisvenue C (2012) Углерод в почве бореальных лесов: распределение, функции и моделирование. Лесное хозяйство 85 (2): 161–184. https://doi.org/10.1093/forestry/cps003
Dulamsuren Ch, Hauck M (2008) Пространственные и сезонные колебания климата на степных склонах горной тайги северной Монголии. Grassl Sci 54 (4): 217–230. https://doi.org/10.1111/j.1744-697X.2008.00128.x
Дуламсурен Ч., Хаук М., Бадер М., Осохжаргал Д., Оюнгерел С., Нямбаяр С., Рунге М., Лойшнер С. (2009) Вода отношения и фотосинтетическая активность у лиственницы сибирской , произрастающей в лесостепном экотоне северной Монголии. Физиол дерева 29(1): 99–110.
https://doi.org/10.1093/treephys/tpn008Dulamsuren Ch, Hauck M, Khishigjargal M, Leuschner HH, Leuschner C (2010a) Изменение климатических тенденций в монгольских таежных лесах влияет на рост и регенерацию Larix sibirica . Экология 163 (4): 1091–1102. https://doi.org/10.1007/s00442-010-1689-y
Dulamsuren Ch, Hauck M, Leuschner C (2010c) Недавняя засуха привела к снижению роста Larix sibirica в западной части Хентея, Монголия . Glob Change Biol 16: 3024–3035. https://doi.org/10.1111/j.1365-2486.2009.02147.x
Dulamsuren Ch, Hauck M, Leuschner HH, Leuschner C (2010b) Индуцированное непарным шелкопрядом снижение роста Larix sibirica в лесостепном экотоне. Дендрохронология 28(4):207–213. https://doi.org/10.1016/j.dendro.2009.05.007
Dulamsuren Ch, Hauck M, Mühlenberg M (2008) Насекомые и мелкие травоядные млекопитающие ограничивают посадку деревьев в северной монгольской степи. Завод Экол 195(1):143–156.
https://doi.org/10.1007/s11258-007-9311-zДуламсурэн Ч., Хишигжаргал М., Лойшнер С., Хаук М. (2014) Реакция ширины годичных колец на потепление климата и выборочные рубки в лиственничных лесах Монгольского Алтая. J Plant Ecol 7(1):24–38. https://doi.org/10.1093/jpe/rtt019
Дуламсурэн Ч., Клинге М., Бат-Энерел Б., Ариунбаатар Т., Туя Д. (2019) Влияние фрагментации леса на плотность пула органического углерода в монгольских лесах. степь. Forest Ecol Manag 433:780–788. https://doi.org/10.1016/j.foreco.2018.10.054
Дуламсурен Ч., Клинге М., Дегенер Дж., Хишигджаргал М., Ченлемуге Т., Бат-Энерел Б., Йеруулт Й., Сайндовдон Д., Ганбаатар К., Цогтбаатар Дж., Лойшнер С., Хаук М. (2016) Плотность углеродного пула и первое оценка общего запаса углерода в монгольской лесостепи. Glob Change Biol 22 (2): 830–844. https://doi.org/10.1111/gcb.13127
Эрасми С., Клинге М., Дуламсурен Ч., Шнайдер Ф., Хаук М. (2021) Моделирование продуктивности сибирских лиственничных лесов по временным рядам Landsat NDVI во фрагментированных лесных насаждениях монгольской лесостепи.
Оценка окружающей среды 193(4):200. https://doi.org/10.1007/s10661-021-08996-1Feldmann E, Glatthorn J, Hauck M, Leuschner C (2018) Новый эмпирический подход к определению продолжительности стадий развития лесов в умеренных старых -ростовые леса. Eur J Forest Res 137(3):321–335. https://doi.org/10.1007/s10342-018-1105-4
Статья Google ученый
Голдаммер Г. (2002 г.) Пожарная ситуация в Монголии. Международные новости о лесных пожарах 26: 75–83
Google ученый
Голдаммер Г. (редактор) (2007 г.) Международные новости о лесных пожарах. ЕЭК ООН и ФАО 36
Goodale CL, Apps MJ, Birdsey RA, Field CB, Heath LS, Houghton RA, Jenkins JC, Kohlmaier GH, Kurz W, Liu S, Nabuurs G-J, Nilsson S, Shvidenko AZ (2002) Forest углерод поглощается в северном полушарии. Приложение Ecol 12(3):891–899. https://doi.org/10.1890/1051-0761(2002)012[0891:FCSITN]2.
0.CO;2Статья Google ученый
Хайс М., Хитрый М., Горсак М. (2016) Лесостепь, связанная с экспозицией: разнообразный тип ландшафта, определяемый топографией и климатом. J Засушливая среда 135: 75–84. https://doi.org/10.1016/j.jaridenv.2016.08.011
Статья Google ученый
Хансен М.С., Потапов П.В., Мур Р., Ханчер М., Турубанова С.А., Тюкавина А., Тау Д., Стехман С.В., Гетц С.Дж., Лавленд Т.Р., Коммаредди А., Егоров А., Чини Л., Джастис Ко, Таунсенд JRG (2013 ) Глобальные карты высокого разрешения 21 ст -в. Лесопокров. Наука 342 (6160): 850–853. https://doi.org/10.1126/science.1244693
CAS Статья пабмед Google ученый
Хаук М., Дуламсурен Ч., Баяртогтох Б., Улыкпан К., Буркитбаева Ю.Д., Отгонжаргал Е., Титов С.В., Энхбаяр Т., Сундетпаев А.К., Бекет Ю., Лойшнер С. (2014) Взаимосвязь между моделями разнообразия сосудистых растений, лишайников и беспозвоночные в экотоне среднеазиатской лесостепи.
Biodivers Conserv 23 (5): 1105–1117. https://doi.org/10.1007/s10531-014-0648-zХессл А.Э., Ария У., Браун П., Бьямбасурен О., Грин Т., Джейкоби Г., Сазерленд Э.К., Начин Б., Максвелл Р.С., Педерсон Н., де Гранпре Л., Саладыга Т., Тардиф Дж. К. (2012) Реконструкция истории пожаров в Центральная Монголия из годичных колец. Int J Wildland Fire 21 (1): 86. https://doi.org/10.1071/WF10108
Статья Google ученый
Hilbig W (1995) Растительность Монголии. SPB Acad Publ, Амстердам
Google ученый
МГЭИК (2013 г.) Изменение климата 2013 г. — основы физической науки. Издательство Кембриджского университета, Кембридж
Google ученый
Иванова Г.А., Иванов В.А., Кукавская Е.А., Соя А.Ю. (2010) Повторяемость лесных пожаров в сосновых насаждениях Тувы, Россия. Environ Res Lett 5(1):15002. https://doi.
org/10.1088/1748-9326/5/1/015002Статья Google ученый
Jacob M, Bade C, Calvete H, Dittrich S, Leuschner C, Hauck M (2013) Значение перезрелых и гниющих деревьев для запасов углерода в естественном еловом лесу Центральной Европы. Экосистемы 16 (2): 336–346. https://doi.org/10.1007/s10021-012-9617-0
CAS Статья Google ученый
Джарвис П., Сожье Б., Шульце Э.-Д. (2001) Продуктивность бореальных лесов. В: Рой Дж., Сожье Б., Муни Х. (ред.) Глобальная продуктивность Земли. Academic Press, Сан-Диего, стр. 211–244. https://doi.org/10.1016/B978-012505290-0/50011-9
Глава Google ученый
Каргер Д.Н., Конрад О., Бёнер Дж., Каволь Т., Крефт Х., Сориа-Ауза Р.В., Циммерманн Н.Е., Линдер Х.П., Кесслер М. (2017) Климатология с высоким разрешением для участков земной поверхности. Научные данные 4(1):170122.
https://doi.org/10.1038/sdata.2017.122Статья пабмед ПабМед Центральный Google ученый
Хансариторе Э., Дуламсурен Ч., Клинге М., Ариунбаатар Т., Бат-Энерел Б., Батсайхан Г., Ганбаатар К., Сайндовдон Д., Йеруулт Й., Цогтбаатар Дж., Туя Д., Лойшнер С., Хаук М. (2017b) Более высокая чувствительность сибирских животных к потеплению климата лиственницы на мелких, чем на крупных лесных островах в раздробленной монгольской лесостепи. Glob Change Biol 23 (9): 3675–3689. https://doi.org/10.1111/gcb.13750
Хансариторе Э., Эльдаров М., Ганбаатар К., Сайндовдон Д., Лойшнер С., Хаук М., Дуламсурен С. (2017a) Возрастная структура и тенденции годового прироста стебля лиственницы sibirica в двух соседних лесостепных районах Монголии, различающихся историей землепользования. Деревья 31(6):1973–1986. https://doi.org/10.1007/s00468-017-1601-z
Статья Google ученый
Khishigjargal M, Dulamsuren C, Lkhagvadorj D, Leuschner C, Hauck M (2013) Контрастная реакция плотности рассады и саженца на плотность скота в монгольской лесостепи.
Завод Экол 214(11):1391–1403. https://doi.org/10.1007/s11258-013-0259-xСтатья Google ученый
Клинге М., Бёнер Дж., Эразми С. (2015) Моделирование границ леса и моделей распределения лесов с использованием данных дистанционного зондирования в горном регионе полузасушливой Центральной Азии. Биогеонауки 12 (10): 2893–2905. https://doi.org/10.5194/bg-12-2893-2015
Статья Google ученый
Клинге М., Дуламсурен С., Эрасми С., Каргер Д.Н., Хаук М. (2018) Влияние климата на жизнеспособность растительности на границе бореальных лесов Монголии. Биогеонауки 15(5):1319–1333. https://doi.org/10.5194/bg-15-1319-2018
Статья Google ученый
Клинге М., Зауэр Д. (2019) Пространственная модель позднеледникового и голоценового климатического и экологического развития в Западной Монголии — критический обзор и синтез.
Quaternary Sci Rev 210: 26–50. https://doi.org/10.1016/j.quascirev.2019.02.020Статья Google ученый
Клинге М., Шнайдер Ф., Дуламсурен Ч., Арндт К., Баярсайхан У., Зауэр Д. (2021) Взаимосвязи между рельефом, растительностью, нарушениями и вечной мерзлотой в лесостепи Центральной Монголии. Earth Surf Process Landforms 46 (9)): 1766–1782. https://doi.org/10.1002/esp.5116
Körner C (2012) Альпийские линии деревьев: функциональная экология глобальных высотных пределов деревьев. Springer, Basel, Heidelberg, New York, Dordrecht, London
Ковалев А.А., Эрдэнэбаатар Д. (2009) Открытие новых культур бронзового века в Монголии по данным международной центральноазиатской археологической экспедиции. В: Bemmann J, Parzinger H, Pohl E, Tseveendorj D (eds) Текущие археологические исследования в Монголии, Бонн, стр. 149.–170
Ковальковский К., Старкель Л. (1984) Высотные пояса геоморфологических процессов в южной части Чангайских гор.
(Монголия). Конезавод Геоморфол Карпато-Балкан 18:95–115Google ученый
Liu H, Park Williams A, Allen CD, Guo D, Wu X, Anenkhonov OA, Liang E, Sandanov DV, Yin Y, Qi Z, Badmaeva NK (2013) Быстрое потепление ускоряет снижение роста деревьев в полузасушливых районах леса внутренней Азии. Глоб изменить биол 19(8): 2500–2510. https://doi.org/10.1111/gcb.12217
Статья Google ученый
Лхагвадорж Д., Хаук М., Дуламсурэн Ч., Цогтбаатар Дж. (2013a) Кочевой скотоводческий образ жизни в лесостепи Монгольского Алтая в условиях меняющейся экономики и потепления климата. J Засушливая среда 88: 82–89. https://doi.org/10.1016/j.jaridenv.2012.07.019
Лхагвадорж Д., Хаук М., Дуламсурэн Ч., Цогтбаатар Дж. (2013b) Двадцать лет после деколлективизации: Мобильное животноводство и его воздействие на окружающую среду в лесостепь. Гум Экол 41 (5): 725–735. https://doi.
org/10.1007/s10745-013-9599-3Луйссарт С., Инглима И., Юнг М., Ричардсон А.Д., Райхштейн М., Папале Д., Пьяо С., Шульце Э.-Д., Вингейт Л., Маттеуччи Г., Арагао Л., Обине М., Бир С., Бернхофер С., Блэк К., Бональ Д., Боннефон Дж., Чемберс Дж., Сиаис П., Кук Б., Дэвис К., Долман А., Гилен Б., Гулден М., Грейс Дж., Гранье А., Грелль А., Гриффис Т., Грюнвальд Т., Гвидолотти Г., Хэнсон А., Хардинг Р., Холлинджер Д., Хутира Л., Колари П., Круйт Б., Кутч В., Лагергрен Ф., Лаурила Т., Лоу Б., Ле Мэр Г., Линдрот А., Лустау Д., Малхи Ю., Матеус Дж., Мильявакка М., Миссон Л., Монтаньяни Л., Монкрифф Дж., Мурс Э., Мангер В., Никинмаа Э., Оллингер С., Пита Г., Ребманн С., Рупсард О., Сайгуса Н., Санс М., Зойферт Г., Сьерра С., Смит М., Тан Дж., Валентини Р., Весала Т., Янссенс I (2007) CO 2 баланс бореальных, умеренных и тропических лесов, полученный из глобальной базы данных. Glob Change Biol 13 (12): 2509–2537. https://doi.org/10.1111/j.1365-2486.2007.01439.x
Статья Google ученый
Miehe G, Miehe S, Böhner J, Kaiser K, Hensen I, Madsen D, Liu J, Opgenoorth L (2014) Сколько лет человеческому следу в крупнейшей в мире альпийской экосистеме? Обзор мультипрокси записей с Тибетского нагорья с точки зрения экологов.
Четвертичная научная версия 86:190–209. https://doi.org/10.1016/j.quascirev.2013.12.004Статья Google ученый
Михе Г., Михе С., Уилл М., Опгеноорт Л., Дуо Л., Дорге Т., Лю Дж. (2007) Инвентаризация лесных реликтов на пастбищах южного Тибета (Сизанг, Китай). Завод Экол 194(2):157–177. https://doi.org/10.1007/s11258-007-9282-0
Статья Google ученый
Мухортова Л., Щепащенко Д., Швиденко А., МакКаллум И., Кракснер Ф. (2015) Вклад почвы в углеродный баланс российских лесов. Агрик Фор Метеорол 200:97–108. https://doi.org/10.1016/j.agrformet.2014.09.017
Статья Google ученый
Нямжав Б., Голдаммер Г., Уибриг Х. (2007 г.) Пожарная ситуация в Монголии. Международные новости о лесных пожарах 36: 46–66
Google ученый
Пан Ю., Бердси Р.
А., Фанг Дж., Хоутон Р., Кауппи П.Е., Курц В.А., Филлипс О.Л., Швиденко А., Льюис С.Л., Канаделл Дж.Г., Сиаис П., Джексон Р.Б., Пакала С.В., Макгуайр А.Д., Пиао С., Раутиайнен A, Sitch S, Hayes D (2011) Большой и постоянный поглотитель углерода в лесах мира. Наука 333 (6045): 988–993. https://doi.org/10.1126/science.1201609CAS Статья пабмед Google ученый
Поултер Б., Педерсон Н., Лю Х., Чжу З., Д’Арриго Р., Сиаис П., Дави Н., Франк Д., Леланд С., Минени Р., Пиао С., Ван Т. (2013) Последние тенденции в лесах внутренней Азии динамика к температуре и осадкам свидетельствует о высокой чувствительности к изменению климата. Agric For Meteorol 178–179: 31–45. https://doi.org/10.1016/j.agrformet.2012.12.006
Артикул Google ученый
Рихтер Х., Хаазе Г., Бартель Х. (1963) Die Golezterrassen. Petermanns Geogr Mitt 107: 183–192
Google ученый
Сакамото К.
, Томонари М., Ария У., Накагири Э., Мацумото Т.К., Акадзи Й., Отода Т., Хиробе М., Начин Б. (2021 г.) Влияние крупномасштабного лесного пожара с последующей незаконной вырубкой на восстановление бореальных лесов в Монголии. Пейзаж Ecol Eng 17 (3): 267–279. https://doi.org/10.1007/s11355-021-00457-8Статья Google ученый
Schlütz F, Dulamsuren Ch, Wieckowska M, Mühlenberg M, Hauck M (2008) История растительности в позднем голоцене предполагает естественное происхождение степей в северной монгольской горной тайге. Палеогеогр Палеоклиматол Палеоэколь 261(3-4):203–217. https://doi.org/10.1016/j.palaeo.2007.12.012
Швиденко А.З., Щепащенко Д. (2014) Углеродный баланс лесов России. Сибирский лес J 1:69–92
Google ученый
Sommer M, Treter U (1999) Die Lärchenwälder der Gebirgswaldsteppe in den Randgebieten des Uvs Nuur-Beckens. Эрде 130:173–188
Google ученый
Сугимото А.
, Янагисава Н., Найто Д., Фудзита Н., Максимов Т.С. (2002) Значение вечной мерзлоты как источника воды для растений восточно-сибирской тайги. Экологический рез. 17(4):493–503. https://doi.org/10.1046/j.1440-1703.2002.00506.xАртикул Google ученый
Чебакова Н.М., Парфенова Е., Соя А.Ю. (2009) Влияние климата, вечной мерзлоты и пожаров на изменение растительности Сибири в условиях меняющегося климата. Environ Res Lett 4(4):45013. https://doi.org/10.1088/1748-9326/4/4/045013
Статья Google ученый
Тернер М., Бир С., Санторо М., Карвалье Н., Вутцлер Т., Щепащенко Д., Швиденко А., Комптер Э., Аренс Б., Левик С.Р., Шмуллиус С. (2014) Запасы углерода и плотность северных бореальных и умеренных лесов. Глоб Эколь Биогеогр 23(3):297–310. https://doi.org/10.1111/geb.12125
Статья Google ученый
Третер У (1996) Гебиргс-Вальдстепп ин дер Монголей.
Геогр Рундш 48: 655–661Google ученый
Цогтбаатар Дж. (2004) Потребности в обезлесении и лесовосстановлении в Монголии. Forest Ecol Manag 201(1):57–63. https://doi.org/10.1016/j.foreco.2004.06.011
Статья Google ученый
Умбанховар К.Э., Шиннеман А.Л., Церенкханд Г., Хаммон Э.Р., Лор П., Наил К. (2009) История региональных пожаров на основе угольного анализа отложений девяти озер в западной Монголии. Голоцен 19 (4): 611–624. https://doi.org/10.1177/095968360
39Статья Google ученый
Ункельбах Дж., Дуламсурен С., Пунсалпаамуу Г., Сайндовдон Д., Белинг Х. (2017) Растительность, климат, человек и пожары в лесостепной экосистеме в позднем голоцене, полученные из керна G2-a в Алтайском Таванском Богде Заповедник в Монголии. Veget Hist Archaeobot 27 (5): 665–677. https://doi.org/10.1007/s00334-017-0664-5
Артикул Google ученый
Ункельбах Дж.
, Кашима К., Энтерс Д., Дуламсурен С., Пунсалпаамуу Г., Бехлинг Х. (2019) Палеоэкологическая эволюция в позднем голоцене (Мегхалаи), полученная на основе мультипрокси-исследований озерных отложений из региона Даян-Нур в Монголии. Палеогеог Палеоклиматол Палеоэколь 530:1–14. https://doi.org/10.1016/j.palaeo.2019.05.021Статья Google ученый
Yang Y, Wang Z, Li J, Gang C, Zhang Y, Zhang Y, Odeh I, Qi J (2016) Сравнительная оценка динамики деградации пастбищ в ответ на изменение климата и деятельность человека в Китае, Монголии, Пакистане и Узбекистан с 2000 по 2013 год. J Arid Environ 135:164–172. https://doi.org/10.1016/j.jaridenv.2016.09.004
Статья Google ученый
Ссылки на скачивание
Благодарности
Мы благодарим г-жу Дарамрагчаа Туя из Национального парка Тарвагатай-Нуруу (Тосонценгел Сум, Завханский аймак, Монголия) за неоценимую поддержку нашего исследования. Выражаем благодарность нашим монгольским коллегам г-ну Амарбаясгалану, г-ну Энхжаргалу, г-ну Энх-Агар, г-же Мунхтуя. Мы высоко ценим их гостеприимство и помощь в полевых работах. Мы также выражаем благодарность немецким студентам Мартине Кооб, Тино Пеплау, Янину Клаассену и Тиму Роллваге за их большую поддержку в измерениях биомассы во время полевых работ в Монголии.
Немецкий аэрокосмический центр (DLR) предоставил данные TanDEM-X для исследуемой области (DEM_FOREST 1106). Полевые работы в 2014 г. выполнены при финансовой поддержке Фонда Фольксваген в рамках проекта 87175 «Восстановление лесов и биоразнообразие на лесостепной границе Алтая и Хангая в условиях контрастного развития поголовья скота в Казахстане и Монголии», предоставленного М. Хауку. , Ч. Дуламсурен и К. Лейшнер. Последующая работа финансировалась Deutsche Forschungsgemeinschaft (DFG), проект № 385460422, одобренный М. Клинге, Д. Зауэр и М. Фрехен.
Финансирование
Это исследование финансировалось Фондом Volkswagen (проект № 871759) и Немецким исследовательским советом (Deutsche Forschungsgemeinschaft, DFG) (проект № 385460422).
Информация об авторе
Авторы и организации
Факультет физической географии, Институт географии, Геттингенский университет, Goldschmidtstraße 5, 37077, Геттинген, Германия
Michael Klinge, 00ela Sanider 9 Florian Schneider & Dan0167
Прикладная экология растительности, Факультет окружающей среды и природных ресурсов, Университет Фрайбурга, Tennenbacher Str. 4, 79106, Freiburg, Germany
Choimaa Dulamsuren & Markus Hauck
Institute of Farm Economics, Thünen Institute, Bundesallee 63, 38116, Braunschweig, Germany
Stefan Erasmi
Department of Biology, School of Arts and наук, Национальный университет Монголии, Бага тойруу 47, Сухэ-Батор дуурег, Улан-Батор, Монголия
Уудус Баярсайхан
Авторы
- Майкл Клинге
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Choimaa Dulamsuren
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Florian Schneider
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Стефан Эрасми
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Уудус Баярсайхан
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Daniela Sauer
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Markus Hauck
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
Вклады
MK задумал идеи; MK, ChD, FS, MH, UB и DS участвовали в полевых работах и собирали данные; МК, ЧД и СЭ проанализировали данные; MK, ChD, MH и DS написали статью. Автор(ы) прочитал(и) и утвердил окончательный вариант рукописи.
Автор, ответственный за переписку
Майкл Клинге.
Декларация этики
Одобрение этики и согласие на участие
Неприменимо.
Согласие на публикацию
Неприменимо.
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Дополнительный файл 1: Таблица S1 Средняя биомасса живых деревьев (над и под землей) для различных категорий леса и условий участка. Участки, условия на которых не были четко определены, были исключены из соответствующей части анализа. Два более низких и более высоких класса размера леса в зоне с преобладанием леса были объединены для статистического анализа из-за небольшого набора данных по опушкам леса. SE = стандартная ошибка, n = количество участков. Подчеркнутые данные не являются репрезентативными из-за недостаточного размера соответствующего набора данных. Рис. S1 NDVI м лесов в районе исследования. Средние арифметические значения NDVI по семи спутниковым снимкам Sentinel 2 (17.05.2018, 11.06.2018, 25.08.2018, 4.09.2018, 14.09.2018, 19.09.2017, 16.07.2016). Заштрихованная рельефная иллюстрация основана на данных TanDEM-X.
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате, при условии, что вы укажете соответствующую ссылку на оригинальный автор(ы) и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons на статью, если иное не указано в кредитной строке материала. Если материал не включен в лицензию Creative Commons статьи, а ваше предполагаемое использование не разрешено законом или выходит за рамки разрешенного использования, вам необходимо получить разрешение непосредственно от правообладателя. Чтобы просмотреть копию этой лицензии, посетите http://creativecommons.org/licenses/by/4.0/.
Перепечатки и разрешения
Об этой статье
За пределами дихотомии лес-луг: Градиентная организация местообитаний в лесостепях , саванны, лесные пастбища и лесостепи) покрывают значительную часть земной поверхности (House et al., 2003), и их динамика (например, Innes et al., 2013), модели биоразнообразия (например, Erdős et al. ., 2018a, b), а также природоохранное значение (например, Bergmeier et al., 2010; Prevedello et al., 2018) находятся в центре внимания экологических исследований. Компоненты таких систем можно условно разделить на два основных типа: леса и пастбища, которые существенно различаются по нескольким биотическим (например, видовому составу и площади листьев) и абиотическим (например, солнечному излучению и влажности почвы) параметрам (Breshears, 2006). . Наличие структурно несхожих участков увеличивает пространственную неоднородность и способствует поддержанию видового разнообразия, экосистемных услуг и экологической стабильности (Manning et al.
, 2009).; Сантана и др., 2017; Толгиеси и др., 2018).Лесостепи образуют контактную зону между сомкнутыми лесами умеренного пояса и безлесными степями и представляют собой хрестоматийный пример мозаики лес-луг (Erdős et al., 2018a). В этих районах чередование участков леса и луга является неотъемлемой чертой экосистемы (Erdős et al., 2014; Hais et al., 2016; Lashchinskiy et al., 2017; Bátori et al., 2018). Было подчеркнуто, что такие сложные экосистемы нельзя понять, просто изучая компоненты лесов и пастбищ по отдельности (House et al., 2003; Erdős et al., 2018b). Вместо этого необходим целостный подход с интегрированным взглядом на всю мозаику, поскольку эти компоненты экологически взаимосвязаны несколькими способами. Например, некоторым видам животных необходимы оба компонента для их полного жизненного цикла, в то время как некоторые виды растений могут менять предпочтения между компонентами в годы с разными погодными условиями (Bartha et al., 2008; Luza et al., 2014).
Леса и пастбища имеют различные экологические, структурные и композиционные характеристики, и взаимодействие этих характеристик приводит к возникновению специфических пограничных сообществ (Erdős et al. , 2013, 2014). Опушки являются основными местами обитания деревьев; таким образом, они играют важную роль в динамике лесостепи (Erdős et al., 2015). Эти результаты подтверждают идею о том, что, помимо лесных и пастбищных компонентов, опушки леса также должны быть признаны важными местами обитания в этих мозаичных экосистемах.
Общеизвестно, что безлесные территории оказывают значительное влияние на микроклиматические параметры периферийных участков леса, что может затрагивать всю площадь небольших участков леса (Schmidt et al., 2017). Таким образом, можно предположить, что разноразмерные участки леса существенно различаются как по структуре, так и по видовому составу.
Поскольку опушки являются переходными местообитаниями между лесным и луговым компонентами, а небольшие участки леса могут по некоторым характеристикам быть переходными между более крупными участками леса и лугами, мы предполагаем, что местообитания лесостепных мозаик могут располагаться по градиенту. Однако этому явлению в лесостепи до сих пор уделялось мало внимания со стороны ученых (Erdős et al. , 2018b).
В этом исследовании мы оценили шесть типов местообитаний песчаных лесостепей: большие участки леса, средние участки леса, малые участки леса, северные опушки леса, южные опушки леса и луга. Наш вопрос заключался в том, можно ли, основываясь на видовом составе, функциональных группах видов и значениях экологических индикаторов, сгруппировать эти шесть типов местообитаний в две отдельные категории местообитаний (что поддержало бы дихотомическое описание), или же для этой сложной системы лучше подходит другая парадигма.
Материалы и методы
Район исследования
Мы провели наше исследование в Песчаном хребте Кишкунсаг (Центральная Венгрия), недалеко от самых западных продолжений евразийского лесостепного пояса. Мы выбрали шесть исследовательских участков, где лесо-луговые мозаики сохранились в условиях, близких к естественным: Чевхараст (47°17′ с.ш., 19°24′ в.д.), Татарсентдьёрдь (47°02′ с.ш., 19°22′ в.д.), Фюлёфаза. (46°52′ северной широты, 19°25′ восточной долготы), Боча (46°41′ северной широты, 19°27′ восточной долготы), Тазлар (46°30′ северной широты, 19°30′ восточной долготы) и Недьестелеп (46° северной широты). 17′, Е 19°35′). Каждый участок характеризуется стабилизированными известняковыми песчаными дюнами эолового происхождения. Высота участков варьируется от 105 до 140 м над уровнем моря. Среднегодовая температура 10,0–10,7°С, среднегодовое количество осадков 520–580 мм (Dövényi, 2010). Почвы представляют собой бедные гумусом песчаные почвы с низкой водоудерживающей способностью (Várallyay, 1993).
Естественная растительность исследуемых участков представляет собой мозаику участков леса и лугов. Травяной компонент мозаики растительности в основном образован открытыми многолетними песчаными лугами, в которых преобладают Festuca vaginata , Stipa borysthenica и S. capillata . Другие распространенные виды включают Alkanna tinctoria , Dianthus serotinus , Euphorbia seguieriana , Fumana procumbens , Koeleria glauca и Potentilla areria . Лесная составляющая представлена разновеликими участками можжевелово-тополиных насаждений. Ярус полога имеет высоту 15–20 м, в нем преобладают Populus alba и P. × canescens . Навес открытый, и его покрытие обычно колеблется от 40 до 70%. Высота кустарникового яруса составляет 1–3 м, а его покрытие обычно колеблется от 20 до 80 %. Наиболее распространены кустарники Crataegus monogyna , Juniperus communis , Ligustrum vulgare и Rhamnus cathartica . Травянистый ярус редкий (10–40%) и состоит из таких видов, как Anthriscus cerefolium , Carex flacca , Carex liparocarpos 9.0727, Pimpinella saxifraga , Polygonatum odoratum и Stellaria media , а также многочисленные сеянцы деревьев и кустарников. Края довольно узкие, обычно с обширным покровом из кустарников (в основном C. monogyna и J. communis ) и трав (например, Calamagrostis epigeios , Poa angustifolia , Teucrium chamaedrys ). Названия видов растений даны по Király (2009).
Все исследовательские центры находятся под защитой закона. Их нынешние мозаичные узоры являются результатом полузасушливого климата, дополненного экстремальными почвенными условиями. Имеющиеся данные свидетельствуют о том, что пространственное расположение лесных и пастбищных участков стабильно, а существование пастбищного компонента не зависит от выпаса скота, пожаров или других форм нарушений (Фекете, 19).92; Erdős и др., 2015). Благодаря правовой охране антропогенные нарушения на исследуемых объектах минимальны (низкий уровень неразрушающих исследований и строго регламентированный туризм). Natural disturbances include the effects of grazers and browsers ( Capreolus capreolus , Cervus elaphus , Dama dama , and Lepus europaeus ) as well as the activity of burrowing animals ( Meles meles , Talpa europaea , и Vulpes vulpes ), но их влияние на лесо-луговой баланс, по-видимому, незначительно. В последние десятилетия лесные пожары случались только на одном из исследуемых участков, но в ходе нашего исследования не были опробованы районы, пострадавшие от пожара.
Отбор проб растительности
На каждом участке мы выделили шесть типов местообитаний: крупные участки леса (>0,5 га), средние участки леса (0,2–0,4 га), небольшие участки леса (<0,1 га), северные опушки леса, южные опушки леса и луга. В пределах каждого местообитания 25 м Заложено 2 участка. Этот размер достаточно мал, чтобы опробовать даже самые маленькие участки леса, но достаточно велик для стандартного ценологического исследования. Форма участка была 5 м × 5 м на участках леса и лугах, в то время как мы использовали участки 2 м × 12,5 м на краях, чтобы убедиться, что участки не заходят внутрь леса или пастбища. Предыдущие исследования показали, что форма графика не искажает результаты в этом масштабе (Keeley and Fotheringham, 2005; Bátori et al., 2018). Край был определен как зона за пределами самых удаленных стволов деревьев, но все еще под кронами деревьев. Для краевых участков учитывались только участки леса >0,2 га.
Всего было отобрано 176 площадок: 27 площадок на больших лесных участках, 29 площадок на средних лесных участках и по 30 площадок на небольших лесных участках, северных опушках, южных опушках и лугах. На каждом участке процентное покрытие всех видов сосудистых растений каждого яруса растительности оценивалось визуально в апреле и июле 2016 г. Для каждого вида для последующего анализа данных использовалось наибольшее значение покрытия. Значения процентного покрытия для всех видов, зарегистрированных на 176 исследуемых участках, приведены в дополнительной таблице S1.
Анализ данных
Мы выполнили неметрическое многомерное шкалирование (NMDS) для двух измерений, надежный метод неограниченной ординации, широко применяемый в экологии сообществ (Minchin, 1987), основанный на данных о присутствии-отсутствии видов, с использованием Sørensen – Индекс кости. Для анализа использовался «веганский» пакет R (Oksanen et al., 2018). Несколько (минимум 500, максимум 5000) NMDS были запущены со случайного запуска, чтобы облегчить сходимость к нелокальному оптимуму. Результат был визуализирован с использованием центрирования, масштабирования с половинным изменением и поворота к осям анализа основных компонентов (PCA), поэтому дисперсия точек максимальна по первому измерению.
Чтобы визуализировать перекрытия шести изученных местообитаний и оценить их отчетливость, мы применили методику Lex et al. (2014). Этот метод показывает количество видов, ограниченных определенными типами местообитаний (т. е. обособленность местообитаний), а также количество видов, присутствующих в двух или более местообитаниях (т. е. местообитания перекрываются). Расчеты выполнены в MS Excel, графики подготовлены в Adobe Photoshop 7.0.
Рассчитаны средние значения экологических показателей температуры, влажности почвы и светообеспеченности для каждого участка. Мы использовали значения индикатора Борхиди (1995), которые основаны на значениях Ellenberg et al. (1992), но распространен на Карпатский бассейн. Как показали многочисленные полевые измерения, значения экологических индикаторов могут обеспечить надежную оценку условий участка (например, Schaffers and Sýkora, 2000; Dzwonko, 2001; Tölgyesi et al., 2014). Хотя использование средних значений показателей часто подвергается критике, было показано, что они хорошо работают и имеют прочную теоретическую основу (ter Braak and Gremmen, 1987; Diekmann, 2003). Значения экологических индикаторов предоставляют важную информацию, поскольку они интегрируют флуктуирующие значения во времени (а не предоставляют данные за очень короткий период) и отражают условия участка в отношении потребностей видов (а не просто абсолютные значения параметров окружающей среды) (Zonneveld, 19).83; Барта, 2002 г.; Дикманн, 2003).
Линейные модели смешанных эффектов использовались для проверки влияния типа среды обитания на средние значения показателей температуры, влажности почвы и доступности света. Мы использовали сайт как случайный эффект в анализе. Статистические тесты были реализованы с использованием пакета nlme R (Pinheiro et al., 2013). Выполнение предположений о нормальности и гомоскедастичности моделей проверяли визуальными оценками диагностических графиков. Поскольку остатки модели показали неоднородность дисперсии, мы использовали структуру дисперсии, которая допускала различные остаточные разбросы для каждого уровня объясняющей переменной в соответствии с Zuur et al. (2009 г.). Тесты HSD Тьюки были реализованы для постфактум попарных сравнений категорий местообитаний с использованием пакета multcomp R (Hothorn et al., 2008).
Виды были классифицированы по категориям жизненных форм, типам геоэлементов и группам фитосоциологических предпочтений. Категории были основаны на Horváth et al. (1995) и Борхиди (1995). Типы геоэлементов отражают глобальное распространение видов, в то время как фитосоциологические группы предпочтений описывают региональные предпочтения видов к определенным растительным сообществам. Для фитосоциологических предпочтений учитывались только местные виды, поскольку неместные виды, как правило, имеют неопределенные предпочтения. Распределения частот были рассчитаны для всех трех категорий для каждой среды обитания и сравнивались с использованием критерия хи-квадрат Пирсона.
Все статистические анализы проводились в статистической среде R вер. 2.15.2 (Основная группа разработчиков R, 2010 г.).
Результаты
На 176 участках было обнаружено 232 вида сосудистых растений. NMDS сошлась после 2267 попыток и 90 итераций, достигнув значения напряжения 0,1784. Ориентация показала, что местообитания были выровнены в пространстве ординации в следующей последовательности: большие участки леса – средние участки леса – небольшие участки леса – северные опушки – южные опушки – пастбища (рис. 1). Смена видов происходила в основном постепенно. Хотя среда обитания пастбищ составляла относительно отдельную группу, ее видовой состав в некоторой степени пересекался с таковым на южных окраинах.
Рис. 1. Диаграмма рассеяния ординации неметрического многомерного масштабирования (NMDS) из 176 графиков. Отображаются только 20 лучших видов в соответствии с корреляцией с пространством ординации (т. Е. Квадратным корнем качества подгонки). LF — крупные участки леса; МП – участки среднего леса; SF – небольшие участки леса; СВ, северные края; ЮВ, южные края; Г, луга. Центроиды для каждого местообитания нарисованы более крупными знаками. Виды следующие: 1, Kochia laniflora ; 2, Erophila verna ; 3, Алканна красильная ; 4, Crepis rhoeadifolia ; 5, Polygonum arenarium ; 6, Holosteum umbellatum ; 7, Мятлик луковичный ; 8, Сирения Кана ; 9, Arenaria serpyllifolia ; 10, Euphorbia seguieriana ; 11, Stipa borysthenica + capillata ; 12, Festuca vaginata ; 13, Ligustrum vulgare ; 14, Барбарис обыкновенный ; 15, Rhamnus catharticus ; 16, Celtis occidentalis ; 17, Crataegus monogyna ; 18, Bromus sterilis ; 19, Stellaria media ; 20, Anthriscus cerefolium .
Значительное количество видов было общим для всех местообитаний (рис. 2). Лесные местообитания (т. е. участки леса плюс опушки) были тесно связаны, о чем свидетельствует большое количество обычных видов. Количество видов, ограниченных отдельными типами местообитаний, также было многочисленным, особенно на северных окраинах, хотя на лугах, небольших участках леса и на южных окраинах также было значительное количество видов, которые не встречались в других местах.
Рис. 2. Взаимоотношения изученных местообитаний по перекрытию и различию видового состава. На верхней панели показано количество тех видов, которые были обнаружены в местообитаниях, обозначенных точками на нижней панели. Например, в первом столбце показано, что 31 вид был обнаружен во всех шести местообитаниях, тогда как в третьем столбце показано, что 19 видов обитают только в пастбищах, обращенных на север. Небольшая панель в левом нижнем углу показывает общее (то есть объединенное) количество видов в каждой среде обитания. LF — крупные участки леса; МП – участки среднего леса; SF – небольшие участки леса; СВ, северные края; ЮВ, южные края; Г, луга.
Общее (т. е. объединенное) количество видов было самым высоким на северных окраинах (рис. 2), за которыми следовали южные окраины и небольшие участки леса. В средних лесных участках было меньше видов, в то время как в больших лесных участках и лугах было наименьшее общее количество видов.
Тип местообитания существенно повлиял на средние значения экологического показателя температуры F (5,165) = 92,55, P < 0,001, влажности почвы F (5,165) = 157,325, P < 0,001, а доступность света F (5,165) = 226,47, P < 0,001. Средние экологические показатели температуры показали, что самые низкие значения были у крупных и средних участков леса, а самые высокие значения у лугов (рис. 3А). Небольшие лесные участки, северные опушки и южные опушки были промежуточными, но их значения были аналогичны значениям больших и средних лесных участков. Средние экологические показатели влажности почвы демонстрировали непрерывную тенденцию к снижению от крупных и средних участков леса к пастбищам, которые имели самые низкие значения (рис. 3Б). Что касается средних экологических показателей наличия света, то наблюдалась четко выраженная, постепенно увеличивающаяся тенденция от больших участков леса к пастбищам (рис. 3С).
Рис. 3. Средние значения экологических показателей (А) температуры, (Б) влажности почвы и (В) светообеспеченности для изучаемых местообитаний. Среды обитания, не разделяющие букву, существенно различаются. LF — крупные участки леса; МП – участки среднего леса; SF – небольшие участки леса; СВ, северные края; ЮВ, южные края; Г, луга.
Критерий хи-квадрат Пирсона показал, что частотные распределения категорий жизненных форм, типов геоэлементов и групп фитосоциологических предпочтений в шести типах местообитаний различались (хи-квадрат = 635,2, df = 30, P < 0,001; хи-квадрат = 339,2, df = 25, P <0,001; хи-квадрат = 587,8, df = 25, P < 0,001 соответственно). Категории жизненных форм постоянно менялись от больших участков леса к пастбищам (рис. 4А). Крупные и средние участки леса существенно не отличались друг от друга, равно как и мелкие участки леса на северных опушках. Доля кустарников и деревьев уменьшилась в сторону лугов. Терофиты были наиболее распространены на пастбищах, тогда как гемикриптофиты достигали максимального значения на северных опушках.
Рис. 4. Распределение частот для (A) категорий жизненных форм, (B) типов геоэлементов и (C) групп фитосоциологических предпочтений шести изученных местообитаний. Th, терофиты; HTh, гемитерофиты; Г — геофиты; Н — гемикриптофиты; N — нанофанерофиты; Ch — хамефиты; М — микрофанерофиты; ММ – мезо- и мегафанерофиты; Adv, adventives; Cos, космополиты; Конец, эндемики; SubMed, субсредиземноморские виды; Cont, континентальные виды; Eur, европейские и евразийские виды; индифферентные, индифферентные виды; другие виды опушек, кустарников и нарушенных сообществ; злаковые сорняки, виды, связанные с пастбищами и сообществами сорняков; для травянистых видов, обычно встречающихся в лесах и лугах; травы, виды, связанные с пастбищами; для лесных видов; LF — крупные участки леса; МП – участки среднего леса; SF – небольшие участки леса; СВ, северные края; ЮВ, южные края; Г, луга. Среды обитания, не разделяющие букву над полосами, существенно различаются.
На больших участках леса преобладали европейские и евразийские виды, доля которых постепенно уменьшалась по мере приближения к пастбищам (рис. 4В). Напротив, доля континентальных видов показала обратную картину. Остальные геоэлементы в основном играли подчиненную роль. На пастбищах было наибольшее количество эндемиков и наименьшее количество адвентивов.
Доля связанных с лесом видов демонстрировала тенденцию к уменьшению от больших участков леса к пастбищам (рис. 4C). Виды, относящиеся к пастбищам, показали противоположную тенденцию. Индифферентные виды (т.е. виды, предпочитающие несколько растительных сообществ) играли важную роль во всех исследованных местообитаниях, но их доля была наибольшей в участках среднего леса.
Обсуждение
Компоненты лесо-луговых мозаичных экосистем удобно классифицировать на две основные категории: леса и пастбища. Классификация основана на доминирующих формах жизни, но она коррелирует с многочисленными другими характеристиками, как биотическими, так и абиотическими (например, Breshears, 2006; Innes et al. , 2013; Luza et al., 2014). Хотя эта классификация может быть приемлемой для некоторых целей, недавние данные свидетельствуют о том, что лесостепи не следует рассматривать как простые двухфазные системы (Erdős et al., 2018b).
Наше исследование выявило четкий градиент по изучаемым характеристикам. Произошла в основном постепенная смена видов с крупных участков леса на пастбища и постепенные изменения значений экологических показателей и функциональных групп видов. Края оказались переходными между лесами и лугами, причем южные края больше похожи на луга. Малые участки леса представляли собой переход между средними участками леса и опушками. Общее количество видов достигло пика вблизи середины градиента, т. е. на северных краях.
В соответствии со средними значениями экологических показателей, рассчитанными в нашем исследовании, более ранние исследования также показали, что в недрах леса более низкая температура и более высокая влажность почвы, чем в соседних безлесных районах (Davies-Colley et al. , 2000; Mosquera et al. , 2014; Schmidt et al., 2017), тогда как ребра, как правило, занимают промежуточное положение по вышеуказанным параметрам (Kapos, 1989; Gehlhausen et al., 2000; Mosquera et al., 2014; Schmidt et al., 2017). Кроме того, мы обнаружили, что значения индикаторов северных краев напоминают значения индикаторов участков леса, что согласуется с выводами Мэтлака (19).93). Напротив, значения показателей краев, обращенных на юг, были наиболее схожими с показателями пастбищ. Рис и др. (2004) предсказали, что края, обращенные на юг, должны отклоняться от внутренней части леса больше, чем края, обращенные на север, из-за большего воздействия солнечного света. Таким образом, представляется вероятным, что изучаемые местообитания представляют собой непрерывный градиент факторов окружающей среды, начиная от самых больших участков леса и заканчивая открытыми пастбищами. Это может иметь чрезвычайно важные последствия. Лесостепи отличаются очень высоким видовым богатством (Zlotin, 2002; Erdős et al. , 2018a), и неоднородность среды обитания была предложена в качестве важной движущей силы этого высокого видового разнообразия (Erdős et al., 2018b; Tölgyesi et al., 2018). . Наши результаты подкрепляют и дополняют эти выводы, подчеркивая, что большое количество видов может найти подходящие места обитания где-то вдоль градиента при условии, что в мозаике сохраняется весь спектр неоднородности.
По большинству изучаемых характеристик небольшие участки леса были аналогичны северным опушкам. Этот результат указывает на то, что небольшие участки леса представляют собой краевые места обитания; то есть они слишком малы, чтобы иметь основную площадь. Показано, что на микроклимат внешних участков лесных массивов большое влияние оказывает ширина в десятки метров (Ries et al., 2004; Hennenberg et al., 2008; Dodonov et al., 2013; Schmidt et al. , 2017).
Как показывают спектры геоэлементов, в песчаных лесах нашего региона преобладают европейские и евразийские виды, а в песчаных лугах больше континентальных видов. Это явление можно объяснить биогеографическими причинами. Лиственные леса простираются широкой полосой от Западной Европы до Внутренней Азии (Walter and Breckle, 19).89; Шульц, 2005). Напротив, естественные пастбища встречаются от Восточной Европы до Дальнего Востока, в то время как большинство пастбищ Центральной и Западной Европы являются вторичными и зависят от деятельности человека, такой как выпас скота и кошение. Луга в Карпатском бассейне можно понимать как самые западные продолжения континентальных пастбищ.
Последовательность местообитаний, выявленная в этом исследовании, может быть названа ценоклином (Whittaker, 1967, 1975). Существование довольно непрерывного градиента исследуемых местообитаний позволяет предположить, что дихотомическое деление лесостепных местообитаний на участки леса и пастбища является серьезным и вводящим в заблуждение упрощением, так как плохо отражает реальную неоднородность лесостепной экосистемы. Мы считаем, что лесостепные паттерны лучше соответствуют градиентной парадигме ландшафтной структуры (Cushman et al. , 2005; McGarigal and Cushman, 2005), которая вместо использования дихотомических категоризаций способна отражать более непрерывные вариации. В частности, при использовании категорий леса и пастбищ все участки леса (независимо от их размеров) и опушки классифицируются в категорию леса без учета различий между ними, что приводит к потере информации (Cushman et al. , 2005). Таким образом, выход за рамки дихотомии лес-луг представляется необходимым для лучшего и более правильного понимания лесостепных экосистем. Наше исследование показывает, что парадигма на основе градиента может оказаться полезной в других мозаичных экосистемах лес-луг, таких как саванны, лесные пастбища или экотоны прерий-леса. Точно так же подход на основе градиента может иметь важное значение в лесных экосистемах, страдающих от сильной антропогенной фрагментации.
Что касается градиента растительности, выявленного в этом исследовании, возникает вопрос, каковы движущие силы этого градиента. В районах песчаных дюн в районе Кискунсаг фоновые факторы, такие как глубина грунтовых вод, влажность почвы и параметры микроклимата, сильно зависят от особенностей рельефа (например, Bátori et al. , 2014; Tölgyesi et al., 2014). Однако как наш полевой опыт, так и более ранние исследования (например, Halupa, 1967; Bölöni et al., 2011) показывают, что участки тополевых лесов можно найти в различных топографических положениях: на вершинах и гребнях песчаных дюн, на наветренных и подветренных склонах дюн. , и в дюнных брюках, хотя эти позиции сильно различаются по абиотическим параметрам. Другими словами, нет очевидной связи между наличием лесных участков разного размера и текущими параметрами окружающей среды. Вероятное объяснение этому двоякое. Во-первых, горизонтальные корни P. alba может достигать 40 м и более (Magyar, 1961; Halupa, 1967; Szodfridt, 1969). Таким образом, деревья, находящиеся в агрессивной среде (например, в сухом гребне дюн), способны достигать почв с более высоким содержанием гумуса и лучшей влагообеспеченностью (например, в провисании дюн). Во-вторых, участки леса иногда зависят от одного или нескольких слоев гумуса, погребенных под песком (Halupa, 1967; Szodfridt, 1969; Bodrogközy, 1982; Molnár, 2003). Эти погребенные слои образовались в более ранние периоды вегетации, поскольку в течение голоцена ветер несколько раз повторно откладывал песчаные дюны (Molnár, 2009).; Молнар и др., 2012). Таким образом, текущая структура лесных участков разного размера может отражать более раннюю топографию песчаных дюн и связанные с ними параметры окружающей среды.
Наше исследование сосредоточилось на лесных местообитаниях, анализируя только один тип пастбищ, открытые многолетние песчаные пастбища, которые преобладают на исследуемых участках. Другие травянистые местообитания играют подчиненную роль на изучаемых участках. Можно предположить, что нелесные местообитания также образуют градиент. Например, Salix rosmarinifolia кустарничковых сообществ распространены в основном в дюнных понижениях с благоприятной водообеспеченностью (Бодрогкозы, 1982; Борхиди и др., 2012). Сомкнутые песчаные степи процветают в полусухих условиях (Borhidi et al., 2012) и, как было показано, по своему составу связаны с опушками леса (Erdős et al. , 2013), в то время как открытые многолетние песчаные луга и однолетние песчаные луга живут в самых суровых условиях. обстоятельства. Кроме того, могут быть значительные различия между пастбищами разного размера, причем наименьшие участки пастбищ с естественными просветами напоминают опушки леса (Molnár, 19).98). Возможное градиентное расположение нелесных сообществ в изучаемой мозаичной экосистеме требует дальнейших исследований для понимания лесостепной неоднородности в рамках градиентной парадигмы ландшафтного строения.
Заявление о доступности данных
Все наборы данных, созданные для этого исследования, включены в статью/дополнительный материал.
Вклад авторов
Идея исследования принадлежит Л.Э., ГК-Д и П.Т. Данные собирали LE, GK-D, ZB, CT, PT и PK. KS, AB-F, CT и PK провели статистический анализ. LE и PT написали рукопись при участии GK-D. Все авторы обсудили результаты и прокомментировали рукопись.
Финансирование
Это исследование финансировалось Венгерским фондом научных исследований (OTKA PD 116114 до LE, OTKA K 112576 до GK-D и OTKA PD 132131 до CT), Национальным бюро исследований, разработок и инноваций (NKFIH K 124796 до ZB, NKFIH K 119225 и KH 129483 до PT), Оперативная программа экономического развития и инноваций (GINOP-2. 3.2-15-2016-00019 до AB-F) и Исследовательская стипендия Янош Бойяи Венгерской академии наук для GK- Д.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы благодарят Долли Толнай и Михая Секе-Тота за их помощь в полевых работах, а также Аттилу Ленгьел (Центр экологических исследований MTA) за его замечания относительно рукоположения.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fpls.2020.00236/full#supplementary-material
Ссылки
Барта, С. (2002). «A változó vegetáció leírása indikátorszámokkal», в Magyar Botanikai Kutatások az Ezredfordulón , изд. Э Саламон-Альберт (Pécs: PTE Növénytani Tanszék), 527–556.
Google Scholar
Барта С. , Кампетелла Г., Рупрехт Э., Кун А., Хази Дж., Хорват А. и др. (2008). Увеличится ли межгодовая изменчивость в сообществах песчаных пастбищ с изменением климата? Комм. Экол. 9, 13–21. doi: 10.1556/ComEc.9.2008.S.4
Полный текст CrossRef | Google Scholar
Батори З., Эрдёш Л., Келемен А., Деак Б., Валко О., Галле Р. и др. (2018). Модели разнообразия в песчаных лесостепях — сравнительное исследование западной и центральной Палеарктики. Биодайверы. Консерв. 27, 1011–1030. doi: 10.1007/s10531-017-1477-7
Полный текст CrossRef | Google Scholar
Батори З., Ленгьел А., Мароти М., Кёрмёци Л., Толдьеси К., Биро А. и др. (2014). Взаимоотношения микроклимата и растительности на островах естественной среды обитания: сохранение видов и перспективы сохранения. Идохарас 118, 257–281.
Google Scholar
Бергмайер Э., Петерманн Дж. и Шредер Э. (2010). Геоботаническое исследование лесопастбищных местообитаний в Европе: разнообразие, угрозы и сохранение. Биодайверы. Консерв. 11, 2995–3014. doi: 10.1007/s10531-010-9872-3
Полный текст CrossRef | Google Scholar
Бодрогкози, Г. (1982). Гидроэкология растительности песчаного лесостепного характера в Эмлекердо в Асотталоме. Акта Биол. Сегед. 28, 13–39.
Google Scholar
Бёлёни Дж., Молнар З. и Кун А. (ред.). (2011). Magyarország Élőhelyei. Вакратот: MTA ÖBKI.
Google Scholar
Борхиди, А. (1995). Типы социального поведения, естественность и относительные значения экологических показателей высших растений венгерской флоры. Акта Бот. Повесили. 39, 97–181.
Google Scholar
Борхиди А., Кеви Б. и Лендваи Г. (2012). Растительные сообщества Венгрии. 907:27 Будапешт: Academic Press.
Google Scholar
Breshears, DD (2006). Континуум лугопастбищных угодий и лесов: тенденции в свойствах экосистем для мозаики древесных растений? Фронт. Экол. Окружающая среда. 4:96–104. doi: 10.1890/1540-92952006004[0096:TGCTIE]2. 0.CO;2
CrossRef Full Text | Google Scholar
Кушман С.А., Гуцвейлер К., Эванс Дж.С. и МакГаригал К. (2005). «Парадигма градиента: концептуальная и аналитическая основа ландшафтной экологии», в Проблемы и перспективы ландшафтной экологии , редакторы Дж. А. Винс и М. Р. Мосс (Кембридж: издательство Кембриджского университета), 112–119.
Google Scholar
Дэвис-Колли, Р. Дж., Пейн, Г. В., и Ван Эльсвейк, М. (2000). Градиенты микроклимата на опушке леса. NZ J. Ecol. 24, 111–121.
Google Scholar
Дикманн, М. (2003). Значения видовых индикаторов как важный инструмент прикладной экологии растений – обзор. Базовое приложение Экол. 4, 493–506. doi: 10.1078/1439-1791-00185
Полный текст CrossRef | Google Scholar
Додонов П., Харпер К. А. и Сильва-Матос Д. М. (2013). Роль опушечной контрастности и структуры леса в опушечном влиянии: растительность и микроклимат опушек в бразильском серрадо. Завод Экол. 214, 1345–1359. doi: 10.1007/s11258-013-0256-0
Полный текст CrossRef | Google Scholar
Дёвеньи, З. (ред.). (2010). Magyarország Kistájainak Katastere. 907:27 Будапешт: MTA FKI.
Google Scholar
Дзвонко З. (2001). Оценка светового и почвенного режима древних и современных редколесий по значениям индикатора Элленберга. Дж. Заявл. Экол. 38, 942–951. doi: 10.1046/j.1365-2664.2001.00649.x
Полный текст CrossRef | Google Scholar
Элленберг Х., Вебер Х. Э., Дюлль Р., Вирт В., Вернер В. и Паулиссен Д. (1992). Zeigerwerte von Pflanzen в Средней Европе. Скрипт. Геоботаника 18, 1–248.
Google Scholar
Эрдёш Л., Амбарлы Д., Аненхонов О. А., Батори З., Черхалми Д., Кисс М. и др. (2018а). Грань двух миров: новый обзор и синтез по евразийским лесостепям. Заяв. Вег. науч. 21, 345–362. doi: 10.1111/avsc.12382
Полный текст CrossRef | Google Scholar
Erdős, L., Gallé, R., Körmöczi, L., and Bátori, Z. (2013). Видовой состав и разнообразие естественных лесных опушек: опушечные реакции и местные опушечные виды. Комм. Экол. 14, 48–58. doi: 10.1556/ComEc.14.2013.1.6
Полный текст CrossRef | Google Scholar
Erdős, L., Kröel-Dulay, G., Bátori, Z., Kovács, B., Németh, C., Kiss, P.J., et al. (2018б). Неоднородность среды обитания как ключ к высокой природоохранной ценности мозаики лес-луг. биол. Консерв. 226, 72–80. doi: 10.1016/j.biocon.2018.07.029
CrossRef Полный текст | Google Scholar
Erdős, L., Tölgyesi, C., Cseh, V., Tolnay, D., Cserhalmi, D., Körmöczi, L., et al. (2015). История растительности, недавняя динамика и перспективы на будущее венгерского песчаного лесостепного заповедника: отношения лес-луг, видовой состав деревьев и распределение размерных классов. Комм. Экол. 16, 95–105. doi: 10.1556/168.2015.16.1.11
Полный текст CrossRef | Google Scholar
Erdős, L., Tölgyesi, C., Horzse, M., Tolnay, D., Hurton, Á, Schulcz, N., et al. (2014). Комплексность местообитаний лесостепной зоны Паннонии и ее природоохранные последствия. Экол. Комплекс 17, 107–118. doi: 10.1016/j.ecocom.2013.11.004
CrossRef Полный текст | Google Scholar
Фекете, Г. (1992). Целостный взгляд на преемственность пересмотрен. Ценозы 7, 21–29.
Google Scholar
Гельхаузен С.М., Шварц М.В. и Аугспургер С.К. (2000). Растительность и микроклиматические краевые эффекты в двух фрагментах смешанно-мезофитного леса. Завод Экол. 147, 21–35. doi: 10.1023/A:1009846507652
Полный текст CrossRef | Google Scholar
Хайс М., Читри М. и Хорсак М. (2016). Лесостепь, связанная с экспозицией: разнообразный ландшафтный тип, определяемый рельефом и климатом. J. Засушливая среда. 135, 75–84. doi: 10.1016/j.jaridenv.2016.08.011
CrossRef Полный текст | Google Scholar
Халупа, Л. (1967). A fehér- és szürkenyár termesztésének néhány kérdése a Duna-Tisza közi homokháton. Эрдесети Лапок 102, 319–323.
Google Scholar
Хенненберг К. Дж., Гетце Д., Саржински Дж. , Ортманн Б., Райнекинг Б., Штайнке И. и др. (2008). Выявление сезонной изменчивости микроклиматических границ и экотонов между лесом и саванной. Базовое приложение Экол. 9, 275–285. doi: 10.1016/j.baae.2007.02.004
Полный текст CrossRef | Google Scholar
Хорват Ф., Добольи З. К., Моршхаузер Т., Лёкеш Л., Карас Л. и Шердахели Т. (1995). FLÓRA adatbázis 1.2. Вакратот: MTA ÖBKI.
Google Scholar
Hothorn, T., Bretz, F., and Westfall, P. (2008). Синхронный вывод в общих параметрических моделях. Биом. Дж. 50, 346–363. doi: 10.1002/bimj.200810425
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
Хаус, Дж. И., Арчер, С., Брешерс, Д. Д., Скоулз, Р. Дж., Взаимодействие дерева и травы Nceas и участники. (2003). Загадки в смешанных древесно-травяных растительных системах. Ж. Биогеогр. 30, 1763–1777 гг. doi: 10.1046/j.1365-2699.2003.00873.x
Полный текст CrossRef | Google Scholar
Иннес, К. , Ананд, М., и Баух, К.Т. (2013). Влияние взаимодействия человека и окружающей среды на устойчивость лесо-луговых мозаичных экосистем. Науч. Респ. 3:2689. doi: 10.1038/srep02689
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Капос, В. (1989). Влияние изоляции на состояние воды в лесных массивах бразильской Амазонки. Дж. Троп. Экол. 5, 173–185. doi: 10.1017/S0266467400003448
Полный текст CrossRef | Google Scholar
Кили, Дж. Э., и Фотерингем, К. Дж. (2005). Влияние формы участка на измерения видового разнообразия растений. Дж. Вег. науч. 16, 249–256. doi: 10.1111/j.1654-1103.2005.tb02362.x
Полный текст CrossRef | Google Scholar
Кирай, Г. (ред.). (2009). Új Magyar Füvészkönyv. Йосвафё: Национальный парк Аггтелек.
Google Scholar
Лащинский Н., Королюк А., Макунина Н., Аненхонов О. и Лю Х. (2017). Продольные изменения видового состава лесов и лугов лесостепной зоны Северной Азии. Фолиа Геобот. 52, 175–197. дои: 10.1007/s12224-016-9268-6
Полнотекстовая перекрестная ссылка | Google Scholar
Лекс А., Геленборг Н., Стробельт Х., Вюйлемот Р. и Пфистер Х. (2014). UpSet: визуализация пересекающихся наборов. IEEE T. Vis. Комп. гр. 20, 1983–1992 гг. doi: 10.1109/TVCG.2014.2346248
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Луза А.Л., Карлуччи М.Б., Харц С.М. и Дуарте Л.Д.С. (2014). Переход от взглядов на леса и пастбища к комплексному подходу к сохранению мозаики лесов и пастбищ. Нац. Conservacao 12, 166–169. doi: 10.1016/j.ncon.2014.09.005
CrossRef Полный текст | Google Scholar
Мадьяр, П. (1961). Альфельдфаситас II. Академия Киадо: Будапешт.
Google Scholar
Мэннинг А.Д., Гиббонс П. и Линденмайер Д.Б. (2009). Разбросанные деревья: дополнительная стратегия для облегчения адаптивных реакций на изменение климата в измененных ландшафтах? Дж. Заявл. Экол. 46, 915–919. doi: 10.1111/j.1365-2664.2009.01657.x
Полнотекстовая перекрестная ссылка | Google Scholar
Мэтлак, Г. Р. (1993). Изменчивость микросреды внутри и между лесными опушками на востоке США. биол. Консерв. 66, 185–194. doi: 10.1016/0006-3207(93)
-K
Полный текст CrossRef | Google Scholar
МакГаригал, К., и Кушман, С.А. (2005). «Градиентная концепция ландшафтной структуры», в Issues and Perspectives in Landscape Ecology , eds JA Wiens and MR Moss (Cambridge: Cambridge University Press), 112–119.. doi: 10.1017/cbo9780511614415.013
CrossRef Full Text | Google Scholar
Minchin, PR (1987). Оценка относительной надежности методов экологического ординирования. Вегетацио 69, 89–107. doi: 10.1007/978-94-009-4061-1_9
Полный текст CrossRef | Google Scholar
Молнар, З. (1998). «Интерпретация особенностей современной растительности на основе исторических данных о ландшафте: пример мозаичного ландшафта лесов и лугов (Лес Надькёрёш, Кискуншаг, Венгрия)», в Экологическая история европейских лесов , редакторы К. Дж. Кирби и К. Уоткинс (Уоллингфорд: CAB International), 241–263.
Google Scholar
Молнар, З. (2003). A Kiskunság Száraz Homoki Növényzete. Будапешт: TermészetBúvár Alapítvány Kiadó.
Google Scholar
Молнар, З. (2009). Дуна-Тиса köze és Tiszántúl fontosabb vegetációtípusainak holocén kori története: irodalmi értékelés egy vegetációkutató szemszögéből. Каниция 16, 93–118.
Google Scholar
Молнар З., Биро М., Барта С. и Фекете Г. (2012). «Прошлые тенденции, настоящее состояние и будущие перспективы венгерских лесостепей», в евразийских степях: экологические проблемы и средства к существованию в меняющемся мире , редакторы М. Дж. А. Вегер и М. А. ван Шталдуйнен (Дордрехт: Springer), 209–252. doi: 10.1007/978-94-007-3886-7_7
Полный текст CrossRef | Google Scholar
Москера, А. М., Мартинес, Дж. П., и Фигероа, А. (2014). Микроклиматические градиенты в переходных зонах андского леса: на примере национального парка Пурас. Науч. Рез. Очерки 9, 703–715. doi: 10.5897/SRE2014.5993
Полный текст CrossRef | Google Scholar
Оксанен Дж., Бланше Ф. Г., Френдли М., Киндт Р., Лежандр П. и МакГлинн Д. (2018). веган: экологический пакет сообщества. Версия пакета R 2.4-6. Доступно в Интернете по адресу: CRAN.R-project.org/package=vega (по состоянию на 25 января 2018 г.).
Google Scholar
Пинейро Дж., Бейтс Д., Деброй С. и Саркар Д., а также основная группа разработчиков R (2013). Nlme: линейные и нелинейные модели смешанных эффектов. Версия пакета R 3.1-108 . Доступно в Интернете по адресу: http://CRAN.Rproject.org/package=nlme. (по состоянию на 28 января 2013 г.).
Google Scholar
Преведелло, Дж. А., Алмейда-Гомес, М., и Линденмайер, Д. Б. (2018). Важность разбросанных деревьев для сохранения биоразнообразия: глобальный метаанализ. Дж. Приложение. Экол. 55, 205–214. doi: 10.1111/1365-2664.12943
Полный текст CrossRef | Академия Google
9Основная группа разработчиков 0002 R (2010 г. ). R: Язык и среда для статистических вычислений. R Фонд статистических вычислений . Доступно в Интернете по адресу: http://www.R-project.org (по состоянию на 31 декабря 2010 г.).Google Scholar
Райс Л., Флетчер Р. Дж., Баттин Дж. и Сиск Т. Д. (2004). Экологические реакции на края среды обитания: объяснение механизмов, моделей и изменчивости. год. Преподобный Экол. Эвол. Сист. 35, 491–522. doi: 10.1146/annurev.ecolsys.35.112202.130148
Полнотекстовая перекрестная ссылка | Google Scholar
Сантана Дж., Порто М., Рейно Л., Морейра Ф., Рибейро П. Ф., Сантос Дж. Л. и др. (2017). Использование бета-разнообразия для обоснования сельскохозяйственной политики и природоохранных мероприятий на средиземноморских сельскохозяйственных угодьях. Дж. Заявл. Экол. 54, 1825–1835 гг. doi: 10.1111/1365-2664.12898
Полный текст CrossRef | Google Scholar
Шафферс А. П. и Сикора К. В. (2000). Достоверность значений индикатора Элленберга для влажности, азота и реакции почвы: сравнение с полевыми измерениями. Дж. Вег. науч. 11, 225–244. doi: 10.2307/3236802
Полный текст CrossRef | Google Scholar
Шмидт М., Йоххайм Х., Керсебаум К.-К., Лишайд Г. и Нендель К. (2017). Градиенты микроклимата, углерода и азота в переходных зонах фрагментированных ландшафтов – обзор. Агр. Лесной метеорол. 232, 659–671. doi: 10.1016/j.agrformet.2016.10.022
CrossRef Полный текст | Google Scholar
Шульц, Дж. (2005). Экозоны мира. 907:27 Берлин: Springer.
Google Scholar
Зодфридт, И. (1969). Borókás-nyárasok Bugac környékén. Бот. Кезлем. 56, 159–165.
Google Scholar
ter Braak, C.F.J., and Gremmen, NJM (1987). Экологические амплитуды видов растений и внутренняя согласованность значений индикатора Элленберга для влажности. Вегетацио 69, 79–87. doi: 10.1007/978-94-009-4061-1_8
Полный текст CrossRef | Google Scholar
Tölgyesi, C., Bátori, Z., and Erdős, L. (2014). Использование статистических тестов относительных экологических показателей для сравнения единиц растительности – различные подходы и методы взвешивания. Экол. индик. 36, 441–446. doi: 10.1016/j.ecolind.2013.09.002
Полный текст CrossRef | Google Scholar
Толдьеси, К., Батори, З., Галле, Р., Урак, И., и Хартель, Т. (2018). Кустарниковые насаждения под деревьями разнообразят травяной ярус в румынской лесопастбищной системе. Пастбище экол. Управлять. 71, 571–577. doi: 10.1016/j.rama.2017.09.004
Полный текст CrossRef | Google Scholar
Варальяй, Г. (1993). «Почвы в районе между реками Дунай и Тиса (Венгрия)», в Флора национального парка Кискунсаг , редакторы Й. Шуйко-Лача и Д. Ковац (Будапешт: Венгерский музей естественной истории), 21–42.
Google Scholar
Уолтер Х. и Брекл С.-В. (1989). Экологические системы геобиосферы 3: Умеренные и полярные зонобиомы Северной Евразии. Берлин: Springer.
Google Scholar
Уиттакер Р. Х. (1967). Градиентный анализ растительности. биол. Ред. 42, 207–264. дои: 10.1111/j.1469-185X.1967.tb01419.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
Уиттакер Р. Х. (1975). Сообщества и экосистемы , второе изд. Нью-Йорк, штат Нью-Йорк: издательство Macmillan Publishing.
Google Scholar
Злотин Р. (2002). «Биоразнообразие и продуктивность экосистем», в сб. Физическая география Северной Евразии , изд. М. Шахгеданова (Оксфорд: издательство Оксфордского университета), 169–190.
Google Scholar
Зонневельд И. С. (1983). Принципы биоиндикации. Окружающая среда. Монит. Оценивать. 3, 207–217. doi: 10.1007/978-94-009-6322-1_2
Полный текст CrossRef | Google Scholar
Зуур А.Ф., Иено Е.Н., Анатолий Н.В., Савельев А. и Смит Г.М. (2009). Модели смешанных эффектов и расширения в экологии с R. Нью-Йорк, штат Нью-Йорк: Springer.
Google Scholar
Степной климат: расположение и характеристики | Климатология
РЕКЛАМА:
В этой статье мы обсудим:- 1. Расположение степного климата 2. Температура степного климата 3. Осадки 4. Характеристики.
Расположение степного климата : Степной климат средних широт (BSk), распространенный на лугах умеренного пояса, расположен во внутренних частях континентов, которые входят в пояс западных ветров, но из-за своего более внутреннего расположения они не получить достаточное количество осадков и, следовательно, луга практически безлесные.
Луговые степи умеренного пояса южного полушария расположены вдоль юго-восточных окраин континентов (рис. 39)..7) и, следовательно, имеют более умеренный климат, чем их аналоги в северном полушарии, из-за большего морского влияния, поскольку они ближе к морской среде.
РЕКЛАМА:
Луга умеренного пояса Евразии, известные как степи, наиболее обширны, поскольку простираются на расстояние более 3200 км от берегов Черного моря через Великую Русскую равнину до предгорий Алтайских гор. Их непрерывность нарушена в нескольких местах горной местностью.
Есть также несколько изолированных участков степей, например, в Голодной (известной как Пустаз) и на равнинах Маньчжурии (Маньчжурские пастбища). Луга умеренного пояса в Северной Америке (распространяющиеся как в Канаде, так и в США) в местном масштабе известны как прерии, которые простираются от предгорий Скалистых гор на западе до биома лиственных лесов умеренного пояса на востоке.
Луга южного полушария включают пампасы Аргентины и Уругвая в Южной Америке, буш-вельд и высокогорный вельд Южной Африки, а также холмы бассейнов Мюррей-Дарлинг на юго-востоке Австралии и луга Кентербери в Новой Зеландии (рис. 39)..7).
Умеренные степи характеризуются континентальным климатом, в котором хорошо выражены экстремальные летние и зимние температуры, но умеренные луга южного полушария отличаются более умеренным климатом. Лето теплое, с температурой июля выше 20°C (Виннипег, Канада) и выше 22°C в январе (Петория, ЮАР, январь — летний месяц в южном полушарии).
РЕКЛАМА:
Зимний сезон становится очень холодным в северном полушарии из-за огромных расстояний умеренных лугов от ближайшего моря. Виннипег фиксирует -20°C в январе. Средние температуры зимнего сезона для Ташкента (Республика Узбекистан) и Семипалатинска (Республика Казахстан) составляют 0°C и -12°C соответственно.
Степной климат южного полушария никогда не бывает суровым, скорее умеренным из-за близости к морю. Средняя зимняя температура колеблется от 1°C до 12°C в южном полушарии.
Степной климат характеризуется высокой годовой амплитудой температур. Например, в Виннипеге (Канада) зарегистрирован средний годовой диапазон 40°C. Ларамин (штат Вайоминг в США) регистрирует годовой диапазон 23,4 ° C (-6,7 ° C в январе и 16,7 ° C в июле).
РЕКЛАМА:
Из-за влияния морской среды среднегодовой диапазон температур в южном полушарии намного ниже, чем в северном полушарии, так как он составляет от 10°C до 12°C. Суточная амплитуда температур также очень высока в умеренном степном климате.
Осадки в степном климате :Среднегодовое количество осадков колеблется от 25 см до 75 см в различных местах степных пастбищ умеренного пояса. Зимние осадки в северном полушарии обычно выпадают в виде снега, и большая часть евразийских степей покрыта снегом в течение нескольких месяцев в течение северных зим. Большая часть годового количества осадков выпадает в летний сезон.
Характеристика степного климата : Евразийский степной климат охватывает самую большую территорию бывшего Советского Союза (ныне Содружество Независимых Государств-СНГ и другие республики бывшего СССР), где он простирается от Восточной Европы до Западной Сибири и между умеренными хвойными лесами в север и засушливые районы на юго-западе.
Евразийские степи подразделяются на:
(i) лесостепи и
(ii) травяные степи, при этом в первой из них среднегодовое количество осадков составляет от 50 до 60 см, а во второй — от 40 до 50 см в среднем за год. годовое количество осадков.
С севера на юг встречаются следующие последовательности растительных сообществ:
(1) Лесостепь (состоящая из дуба, вяза, липы, клена, немногочисленной древесной растительности сибирской степи, такой как береза с примесью осины и ива).
(2) Луговая степь (состоящая из видов дерновых трав, таких как ковыль и овсяница, и многочисленных травянистых цветковых растений, таких как многочисленные травянистые цветочные растения, такие как Trifolium и несколько видов маргариток.
РЕКЛАМА:
(3) Травяная степь (состоящая из злаков, в основном кочкообразующих видов Stipa, нескольких цветущих ксерофитных кустарниковых видов Artemisia.
(4) Полузасушливые ксероморфные степи с преобладанием ксерофитных трав, таких как овсяница и виды ковылей (среднегодовое количество осадков от 30 см до 35 см)
Североамериканские прерии образовались в США и Канаде между предгорьями Скалистых гор на западе и биомом лиственных лесов умеренного пояса на востоке
На основе тенденции уменьшения среднегодового количества осадков с востока (105 см) на запад (40 см) прерии Северной Америки делятся на 3 субрегиона, например:
(1) Высокотравные прерии (наиболее виды самой высокой травы — голубика и просо, которые достигают высоты от 1,5 до 2 м, также встречается несколько участков дуба и гикори),
(2) Смешанные прерии (наиболее обширный покров на Великих равнинах США, смесь средних трав высотой от 0,6 до 1,5 м и низкорослых злаков, таких как голубая стебель, игольчатая трава-ковыль спартеа, июньская трава, а также низкорослые и пучковые травы, такие как буйволиная трава и голубая граммовка, и
(3) Низкотравные прерии (развиты в западной части Великих равнин, где преобладают низкотравные травы высотой 60 см).
Южноамериканские пампасы занимают более 12 процентов территории Аргентины и являются более влажными, чем евразийские степи. Среднее годовое количество осадков уменьшается с востока (прибрежная полоса, 90 см) на запад (45 см).
Таким образом, пампасы делятся на два подтипа, например:
(1) влажная пампа и
(2) субгумидная пампа.
Влажная пампа, развитая в восточной части Аргентины, характеризуется высокими травами, в то время как усиление засушливости на западе приводит к росту низкотравья в западной субгумидной пампе.
Африканский вельд образовался на высокогорных плато различной высоты (от 1500 до 2000 м) в юго-восточной части Южной Африки. Африканские Вельды включают луга с умеренным климатом на юге Трансуолла и Оранжевого Свободного государства в Южной Африке, а также некоторые части Лисото. Рост растений невозможен из-за неопределенности количества осадков, увеличения засушливости, сильных ночных заморозков и высокого дневного диапазона температур в зимний сезон, и, таким образом, сформировались настоящие климаксные пастбища африканских вельдов.